
Abstract
The suitability of any location for a given species is determined by the available resources. However, there are many species that occur in more than one habitat type and their successful conservation may be particularly difficult. The dryad Minois dryas, a locally endangered butterfly, occurs in two contrasting habitats—xerothemic and wet grasslands. We investigated the influence of various habitat characteristics, such as vegetation height, grass cover, proximity of shrubs, plant species composition, Ellenberg indices of trophic and microclimatic conditions, on the microhabitat selection by the species. The nectaring of randomly selected butterflies was observed and habitat characteristics were compared at random points within the meadow and at the butterfly’s nectaring and resting places. The butterflies generally preferred to stay close to shrubs and avoided invasive goldenrods. Thermal conditions and the availability of nectar plants were the factors limiting the dryad’s use of wet grassland. In xerothermic habitats grass cover affected the distribution of butterflies. Concerning the availability of larval host plants, wet meadows proved potentially more favourable, whereas nectar resources for adults were more abundant in xerothermic grasslands. Based on our findings, conservation strategies for this butterfly must differ in the two habitats. Rotational mowing in xerothermic grasslands and the removal of invasive goldenrods in wet grasslands are the recommended actions. At a larger spatial scale, a habitat mosaic composed of xerothermic and wet grasslands in close proximity would seem to be the most suitable areas for the conservation of the dryad.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Habitat availability has become a key problem in conservation biology (WallisDeVries et al. 2002; Collinge et al. 2003; Dennis et al. 2003; Timonen et al. 2011), because the worldwide problem of habitat fragmentation has led to a loss of continuous habitat, reduction in habitat patch sizes and an increase of patch isolation (Andrén 1994; Fahrig 2003). However, habitat delimitation is a complex task (Hall et al. 1997; Dennis et al. 2003; Vanreusel and van Dyck 2007; Dover and Settele 2009). Recently the concept of resource-based habitats has been introduced (Dennis et al. 2003; Dennis 2010; Turlure et al. 2009), which offers an alternative to the traditional classification of landscape into patches and matrix (Vanreusel and van Dyck 2007). The concept assumes that the landscape is a continuum of overlapping resource distributions. Resource availability may affect patch quality and structure, as well as inter-resource distances (Dennis et al. 2006). The resource-based approach may help to recognize and explain the spatial distribution patterns of animals, and to capture the functional and spatial interaction of animals with their environment (Dennis et al. 2003).
The concept of resource-based habitats is particularly important for the conservation of many insects, such as butterflies or bees. For these insects, habitats are determined not only by the presence of foodplants and nectar plants, but also by the microclimate and vegetation structure, which provides crucial components such as resting, roosting, mate-location and oviposition places, as well as shelter from wind and refuge or escape from predators (Sparks and Parish 1995; Shreeve et al. 2001; Konvicka et al. 2002; Hardy and Dennis 2007; Slamova et al. 2011). Thus, for the successful conservation of an endangered species, knowledge of the crucial habitat components as well as the species biology is essential (Dennis et al. 2006; Turlure et al. 2009). However, information about habitat requirements of many endangered invertebrates is still missing and, therefore, the management strategies for their habitats remain unknown (WallisDeVries et al. 2002).
Furthermore, there are many animal species that occur in more than one habitat type (e.g. Bell et al. 2001; Bonte et al. 2002; Mildenstein et al. 2005; Skórka et al. 2006; Hahn et al. 2011). These habitat types are sometimes so distinctive in terms of their resources, vegetation and structure that the behavior and biology of a species may differ substantially depending upon the type (Boudjemadi et al. 1999; Potts et al. 2001; Allen and Thompson 2010). Species conservation, especially for endangered species using differing habitat types, is particularly challenging and requires a differentiated application of methods (Mayberry and Elle 2010). One such species is the dryad Minois dryas (Scopoli, 1763), a grassland butterfly of the family Nymphalidae.
The dryad occurs in two distinct habitat types, namely (1) wet grasslands (ordo Molinietalia) and (2) xerothermic grasslands and steppe-like habitats, dominated by the communities of the class Festuco-Brometea (Krzywicki 1982; Ebert and Rennwald 1991; Dąbrowski 1999; Beneš et al. 2002). Both habitat types are themselves endangered. Cessation of extensive management has led to the expansion of shrubs on both xerothermic and wet grassland, and specifically in our study area also to the invasion of alien goldenrods Solidago spp., causing substantial habitat deterioration (Skórka et al. 2007; Rosin et al. 2012).
The aim of this study was to answer the following questions: (1) are the resources used by the dryad in the two habitat types the same? (2) what are the key factors deciding the choice of microhabitats by the butterflies in the two habitats? and (3) what are the management implications for the conservation of the dryad butterfly?
Methods
Study species and area
The dryad has a Euro-Siberian distribution range, extending from the northern part of the Iberian Peninsula, through Europe and Asia to Japan (Dąbrowski 1999; Kudrna 2002). In Europe, it occurs in a number of countries, though usually only locally (Warecki and Sielezniew 2008). Despite the fact that in the past the species was recorded in various places in Poland, at present its occurrence is restricted to the southern part of the country, namely the Kraków region and the Eastern Carpathians (Buszko and Masłowski 2008). It is protected under Polish law and is listed as a critically endangered species in the Polish Red Data Book of Animals (Głowaciński and Nowacki 2004). On a global scale, in terms of the IUCN Red List its conservation status is of less concern (Van Swaay et al. 2010).
The larval food plants include Poaceae grasses occurring in the aforementioned habitats, mainly Molinia caerulea as well as Festuca rubra, Bromus erectus, Calamagrostis epigeios, Lolium perenne, Arrhenatherum elatius and, exceptionally, Carex segdes (Settele et al. 1999; Głowaciński and Nowacki 2004). The dryad flight period is from the end of July to the beginning of September.
The study was carried out in an extensive grassland area in Kraków, southern Poland, located approximately 8 km south-west of the city centre (Fig. 1, Appendix S1). The dryad occurs there in a mosaic of wet meadows in the Vistula valley and xerothermic calcareous grasslands located on hills. Two meadows of each habitat type were selected, namely, the Skołczanka Reserve (A) and Uroczysko Kowadza (B) in the case of xerothermic grasslands, as well as two wet meadows located on either side of a small wooded hill (Fig. 1, Appendix S1). All grasslands under study are located in the Bielańsko-Tyniecki Landscape Park and are part of the Natura 2000 site “Dębnicko-Tyniecki Obszar Łąkowy” (PLH 120065).

Map of the study area, presenting the investigated xerothermic grasslands (A = the Skołczanka Reserve, B = Uroczysko Kowadza) and wet meadows (C, D)
Field sampling
The field surveys were carried out in August 2009 and 2010, with the exception of the Uroczysko Kowadza meadow, where the fieldwork only took place in 2010. Observations of butterflies were carried out on sunny and windless days, while the measurements of the microhabitats at random points were taken regardless of the weather.
To obtain data on the selection of resting and nectaring places by the dryad, randomly-spotted individuals were followed and the places where they sat on plants were marked with a bamboo pole. In order to describe the general availability of resources and the vegetation composition, 31 and 54 random points were selected respectively at wet and xerothermic meadows through random generation of geographic coordinates. The vegetation within a 1.5-m radius was described for both the random points and the places utilized by the butterflies. Five measurements of the vegetation height (naturally arranged plants), one at the central point and the remaining four 1.5 m apart to the north, south, west and east, were taken with a measuring tape of 1-cm precision, and their mean was calculated. In addition, the distance to the nearest shrub was measured with a precision of 10 cm. Finally, all plant species within a 1.5-m radius were counted and their ground cover was recorded. The cover was measured at the following scale: 1 ≤ 10 %; 2 = 10–20 %, …, 9 = 80–90 % and 10 = 90–100 %. We have also recorded the percentage of bare ground (existent only in xerotermic meadows), but it was not included in the analyses as it was strongly negatively correlated with vegetation height and generally very low and little variable, hardly ever exceeding 10 %.
In each habitat type approximately 40 butterflies were followed. For each individual we noted its sex, and classified its behavior on plants as either resting or nectaring (other types of behavior, e.g. oviposition or copulation, were rarely observed and consequently excluded from the analysis). In order to increase sample size of nectaring observations and to receive the knowledge of the range of plant utilized by the dryads, the behavior of randomly-spotted individuals was also recorded. In such cases the butterflies were followed until they settled on a plant. When the individual started nectaring, the duration of the behavior was measured and plant species was noted.
Statistical analysis
In a preliminary analysis we compared the characteristics of the surveyed meadows, based on the parameters measured in random points. The multivariate ANOVA was applied with the two habitat types as the main groups and meadow ID nested within them (Appendix S2). The analyses revealed sharp differences between wet and xerotermic meadows, which is obvious taking into consideration different plant species composition in both habitats (cf. Appendix S3). In contrast, there were no differences between particular meadows of each type, apart from those in vegetation height and some of the Ellenberg indicator values (Ellenberg 1974) between the two xerotermic meadows. Nevertheless, since the above differences were relatively small as compared with those between habitats, in the subsequent analyses we tested the patterns of habitat selection by the dryad separately for each habitat type, but not for each meadow.
A detrended correspondence analysis DCA (Ter Braak and Prentice 1988) was performed in order to ascertain whether the butterflies occur more often in places with particular plant species composition. Plant species of both habitats were classified according to following groups: recorded nectar sources, potential nectar sources, grass species comprising potential larval host plant, Apiaceae, and other species, among which special attention was given to the invasive goldenrods (see Appendix S3). The analysis was conducted on the basis of data on the percentage covers of the plant species in the above groups. Plant species communities were compared for the random points, as well as for butterfly nectaring and resting places.
We also investigated microhabitat selection by the dryad, comparing the characteristics of butterfly observation points and random points within their habitats. The analyses were performed using multivariate ANOVA with random points and butterfly observations as the main groups, as well as with nectaring and resting behavior nested within the latter group. The tested microhabitat parameters included distance from shrubs, vegetation height, grass cover, and Ellenberg indicators of light, temperature, humidity and trophic conditions (Ellenberg 1974), determined on the basis of plant species composition using Zarzycki’s scale, which is the most appropriate for Poland (Zarzycki et al. 2002). Distances from shrubs were ln-transformed to achieve normality.
In addition, we analysed the dryad preferences for different nectar sources. Nectar plants were classified into the following groups: composite plants (Asteraceae and Dipsacaceae), plants with radial symmetry flowers, plants with dorsiventral symmetry flowers and Apiaceae to analyze nectaring preferences of butterflies (see Appendix S3). The availability of the above groups was then calculated separately for each habitat type, using the entire cover of all the plants in a given group, pooled across all the random points within the habitat type. The proportions of nectaring observations in particular plant groups were compared with their availability, using Bailey’s use-availability tests (Bailey 1980). The same procedure was applied to test the dryad’s preferences towards invasive alien goldenrod and native nectar plants within the composite plants.
A general linear mixed model (GLMM) was applied to investigate the effects of habitat type, flower type, butterfly sex, as well as their interactions and random effect in the form of the meadow number on the ln-transformed nectaring duration. The Apiaceae plants were omitted from the analysis due to the very small number of nectaring cases in this group. Since the inclusion of random factors did not improve the fit of the model, the final one was reduced to a generalized linear model (GLM) comprising only significant factors.
The detrended correspondence analysis DCA was performed in Canoco for Windows 4.52 (Lepš and Šmilauer 2003), while the GLM was done in JMP 9 (SAS Institute 2010). The remaining analyses as well as assumption testing for all the analyses were performed in Statistica 9.0 and MS Excel 2007.
Results
Selection of microhabitats
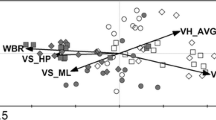
The detrended correspondence analysis biplot in xerothermic habitat (Fig. 2) shows that plant species composition in the dryad resting sites did not differ from that of the random points. However, plant species communities in both the nectaring and resting sites were subsets of the plant communities at the random points, as indicated by the considerably smaller convex polygons. For wet meadows, plant species compositions in the dryad resting and nectaring sites were similar, but, in turn, they only partly overlapped with the plant species composition at the random points (Fig. 2). In wet meadows, several grass species with higher cover were observed, while in the xerothermic habitats there were fewer species of grass and their cover was relatively poor (see Appendices S1 and S3).

Ordination diagram of the detrended correspondence analysis DCA, showing the effects of plant species composition on habitat use by the dryad in the investigated meadows: a xerothermic grasslands, b wet meadows. The black circles represent plant species communities at random points. The grey squares and blank triangles indicate the dryad resting and nectaring places. The mutual distances of the points show their similarity in terms of plant species communities
The analyses of microhabitat conditions based on Ellenberg’s indicators showed that, in the wet meadows, there were no differences in either the quantities of light, humidity or the trophic conditions between the resting and nectaring places as well as in these sites versus random points (Table 1). However, in the case of thermal conditions, the butterflies selected places that were significantly warmer than random points (Table 1). On the other hand, in the xerothermic meadow, the thermal, light, humidity and trophic conditions had no effect on the occurrence of butterflies (Table 1).
The influence of shrubs, vegetation height and grass cover on habitat use
In both habitats butterflies selected sites that were closer to shrubs than random points (Table 2; Fig. 3). In the xerothermic grasslands the dryad also appeared to prefer higher vegetation; the test statistic was close to the significance level (Table 2). In addition, although we have not detected a difference in vegetation height between random points and the dryad observation points, among the latter resting places were characterized by significantly higher grass cover than nectaring places (Table 2, Fig. 3). The ANOVA results for wet meadows revealed no significant differences in vegetation height and grass cover for nectaring, resting, and random points (Table 2).

Mean (with 95 % confidence intervals) distance to shrubs, vegetation height, and grass cover at random points (white bars) as well as in the dryad nectaring and resting places (grey bars) in both investigated habitat types: a, c, e xerotermic grasslands; b, d, f wet meadows
Nectaring preferences
The Bailey’s test results showed that, in the xerothermic habitat the dryad more frequently foraged on plants with dorsiventral symmetry flowers than on composite plants and those with radial symmetry flowers. In wet meadows, in turn, plants with radial symmetry flowers were avoided, and the dryad preferred composite plants. Thus, the butterflies foraged relatively more frequently on composite plants than those with dorsiventral symmetry flowers (Fig. 4). In both habitats, the dryad avoided Apiaceae (Fig. 4). Native species were selected while goldenrods were avoided (Fig. 5).

Nectaring preferences of the dryad in both investigated habitat types (a xerothermic grasslands, n = 332; b wet meadows, n = 134). The grey bars represent the proportions of nectaring observations (with 95 % confidence intervals of the Bailey test) for particular groups of nectaring plants; the white bars show their availability. Bailey’s test P values are also shown if significant: * P < 0.05 ** P < 0.01; *** P < 0.001

Nectaring preferences of the dryad towards native composite plants and invasive alien goldenrods occurring at wet meadows, tested with the Bailey test. The grey bars represent the proportion of nectaring observations (n = 73; shown with 95 % confidence intervals) and the white bars depict their availability. Bailey’s test P values are also shown if significant: *** P < 0.001
The dryad nectaring time was longer in wet meadow than in the xerothermic habitats (Fig. 6). The butterflies fed significantly longer on composite plants (Tukey post hoc, P = 0.001) and plants with dorsiventral symmetry flowers (Tukey post hoc, P = 0.002) than on those with radial symmetry flowers (Fig. 6). Females fed longer than males (Fig. 6). Nectaring time decreased with the progression of the season (GLM: F2,420 = 16.905; P < 0.001; slope = −0.048), (Fig. 6). The GLM showed no significant interactions between habitat type, flower type and sex.

Duration of the dryad nectaring (with 95 % confidence intervals) on different flowering plant groups in both investigated habitat types: a xerothermic grasslands, b wet meadows. White and grey bars represent females and males respectively. GLM model results: n = 426; habitat type effect F1,420 = 4.806, P = 0.029; plant group effect: F2,420 = 8.423, P < 0.001; gender effect F1,420 = 7.364; P = 0.007
Discussion
The results of this study demonstrate a differential use of resources and microhabitats by the dryad in wet meadows and xerothermic habitats; however, some similarities in the utilization of these two contrasting habitats have also been found. In wet meadows, the factor limiting the dryad’s use of microhabitat seems to be the thermal conditions. In the xerotherimic habitat, on the other hand, none of the factors examined affected the distribution of butterflies during resting behavior; however, nectaring butterflies chose slightly drier places, as indicated by the Ellenberg indicators.
It can be concluded that the dryad is a stenothermic species and its distribution may be restricted by unsuitable thermal conditions within its potential habitats. Xerothermic grasslands are usually sunny and warm, as evidenced by the presence of such plant species as Seseli annuum and Vincetoxicum hirundinaria (Zarzycki et al. 2002); thus, in terms of thermal conditions, they are beneficial to the dryad. Nectaring places in the xerothermic habitat were generally open, with low vegetation. At these sites, limestone rocks often protruded above the ground. However, such places may be too dry or too hot for the dryad to carry out all its daily activities there and hence it chooses more humid places with higher vegetation and with higher grass cover for resting.
The results of the study have shown that shrubs are an important component of the dryad habitat in both types of meadow. The presence of shrubs in these meadows is a result of meadow abandonment and the resulting natural succession (Skórka et al. 2007; Rosin et al. 2012). Shrubs provide refuges for butterflies and safe places for mating as copulations were observed there (authors’ unpublished data). Shrubs may also provide shelter for butterflies during windy and rainy conditions (Sparks and Parish 1995; Dover et al. 1997; Dover and Sparks 2000; Dennis and Sparks 2006).
The composition of plant species communities did not affect the distribution of the dryad in xerothermic habitats. Nectaring plant resources were widespread there and thus they apparently did not limit the occurrence of butterflies. The same refers to grasses, i.e. potential larval food plants, despite the fact that they were less numerous in xerothermic grasslands. In contrast, in the wet meadows, the butterflies were concentrated predominantly in places where nectar plants were abundant. In the wet meadows there were already less flowering plants in August than there were in the xerothermic grasslands and this may have affected the dryad’s flight period. In both 2009 and 2010, the number of flying butterflies fell sharply in the wet meadows after 14th August, while they could be observed flying in substantial numbers till the end of that month in the xerothermic grasslands (authors’ unpublished data).
The dryad preferred composite flowers that offer spatially concentrated nectar resources. The longer nectaring time in the wet meadows may imply a lower availability of nectar, especially towards the end of the flight period. This longer nectaring time is in line with the predictions of the marginal value theorem (Charnov 1976; Parker and Stuart 1976; Parker 1992). If the resources, which, in our case are nectaring plants, are scarce, then individuals should opt for the maximal utilization of the resources available at one plant, rather than searching for a new one, because the energy losses during the search may exceed the potential profit to be obtained from new plants. The avoidance of flowers with radial symmetry in the wet meadows is also in agreement with the above explanation. These flowers may have less nectar, which is particularly true for the pollen flowers of Hypericum spp. (Kugler 1955).
Conservation recommendations
Our findings allow a conclusion that a xerothermic habitat is better for the dryad in terms of nectar resources. In turn, a wet meadow habitat is likely to be better in terms of larval food-plant availability, owing to greater abundance of grasses. The conservation strategies for this endangered butterfly must therefore differ for these two habitats. In wet habitats, it would be profitable to increase the cover of favorable nectar sources by proper mowing regimes.
In xerothermic meadows, vegetation height should be diverse to provide both favorable resting and nectaring sites. Thus, it may be good practice to apply rotational grazing or mowing (Dolek and Geyer 1997; WallisDeVries et al. 2002; Cremene et al. 2005), since patches under a diverse mowing regime are preferred by butterflies and other insects (Cizek et al. 2012). A single fragment should be mown every 2–3 years. Mowing should be conducted either in the spring, at the end of May, or in the fall, in mid-September at the earliest (Johst et al. 2006; Grill et al. 2008; Wynhoff et al. 2011). In wet meadows, however, mowing should only take place in the late autumn, because they are also an important habitat for endangered Maculinea (= Phenagris) butterflies (Nowicki et al. 2005, 2007). Mowing is necessary not only to sustain the nectar and host plants favorable to the butterflies, but also to prevent the invasion of some notorious alien plants, such as goldenrods Solidago sp. These alien invasive species are a serious threat for biodiversity, predominantly in wet meadows (Skórka et al. 2007; Powell et al. 2011; Vilà et al. 2011) and the dryad did not use this invasive plant as a nectar source. Alien plants can outcompete native plants such as, for example, grasses (Callaway and Aschehoug 2000) leading to deterioration of the habitat quality. Invasive plants may be actively removed, preferably in the early summer, before they flower, and in the autumn (Güsewell 2003).
In many grassland conservation programmes, shrubs are indicators of habitat deterioration and thus their removal is postulated (Erhardt 1985; Krauss et al. 2004; Lenda and Skórka 2010). In the dryad, however, this is not the case. Shrubs appeared to be a key component of the dryad habitat in both the wet and xerothermic grasslands. Therefore, shrubs should be maintained in these meadows, even when grazing and mowing is applied. However, they should be scarce and scattered within a patch, in order not to overgrow flowering plants.
On a larger spatial scale, the sustaining or creation of a mosaic of both wet and xerothermic meadows located close to one another may be a favorable conservation strategy for the dryad. Obviously, it can only be applied in areas where the topographic conditions allow such a mosaic. Butterflies are known to be able to move between different habitat types in order to seek the appropriate resources (Schwarzwälder et al. 1997). In the case of the dryad, the occurrence of the both wet and xerothermic meadows in close proximity may be also advantageous because local populations of the dryad could utilize both habitat types differentially, depending on annual weather patterns and management. An arrangement of both habitats such as this should increase the viability of the entire metapopulation by reducing the impacts of natural catastrophes such as fires on xerothermic grasslands and flooding in wet meadows. Finally, a mosaic of wet and xerothermic meadows may be the most suitable areas for the reintroduction of the dryad to its former localities.
References
Allen KA, Thompson DJ (2010) Movement characteristics of the Scarce Blue-tailed Damselfly, Ischnura pumilio. Insect Conserv Divers 3:5–14
Andrén H (1994) Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos 71:355–366
Bailey BJR (1980) Large sample simultaneous confidence intervals for the multinomial probabilities based on transformations of the cell frequencies. Technometrics 22:583–589
Bell JR, Wheater CP, Cullen WR (2001) The implications of grassland and heathland management for the conservation of spider communities: a review. J Zool 255:377–387
Beneš J, Konvicka M, Dvorák J, Fric Z, Havelda Z, Pavlícko A, Vrabec V, Weidenhoffer Z (2002) Butterflies of the Czech Republic: distribution and conservation. I, II. SOM, Praha
Bonte D, Baert L, Maelfait JP (2002) Spider assemblage structure and stability in a heterogeneous coastal dune system (Belgium). J Arachnol 30:331–343
Boudjemadi K, Lecomte J, Clobert J (1999) Influence of connectivity on demography and dispersal in two contrasting habitats: an experimental approach. J Anim Ecol 68:1207–1224
Buszko J, Masłowski J (2008) Motyle Dzienne Polski. Koliber, Nowy Sącz
Callaway RM, Aschehoug ET (2000) Invasive plants versus their new and old neighbors: a mechanism for exotic invasion. Science 290:521–523
Charnov EL (1976) Optimal foraging, the marginal value theorem. Theor Popul Biol 9:129–136
Cizek O, Zamecnik J, Tropek R, Kocarek P, Konvicka M (2012) Diversification of mowing regime increases arthropods diversity in species-poor cultural hay meadows. J Insect Conserv 16:215–226
Collinge SH, Prudic KL, Oliver JC (2003) Effects of local habitat characteristics and landscape context on grassland butterfly diversity. Conserv Biol 17:178–187
Cremene C, Groza G, Rakosy L, Schileyko AA, Baur A, Erhardt A, Baur B (2005) Alterations of steppelike grasslands in Eastern Europe: a threat to regional biodiversity hotspots. Conserv Biol 19:1606–1618
Dąbrowski JS (1999) Skalnik driada Minois dryas (Scop.), (Lepidoptera: Satyridae)—gatunek zagrożony wyginięciem na ostatnich znanych stanowiskach w Polsce. Chr Przyr Ojcz 55:91–94
Dennis RLH (2010) A Resource-based habitat view for conservation: butterflies in the British landscape. Wiley-Blackwell, UK
Dennis RLH, Sparks TH (2006) When is a habitat not a habitat? Dramatic resource use changes under differing weather conditions for the butterfly Plebejus argus. Biol Conserv 129:291–301
Dennis RLH, Shreeve TG, Van Dyck H (2003) Towards a functional resource-based concept for habitat: a butterfly biology viewpoint. Oikos 102:417–426
Dennis RLH, Shreeve TG, Van Dyck H (2006) Habitats and resources: the need for a resource-based definition to conserve butterflies. Biodivers Conserv 15:1943–1966
Dolek M, Geyer A (1997) Influence of management on butterflies of rare grassland ecosystems in Germany. J Insect Conserv 1:125–130
Dover J, Settele J (2009) The influences of landscape structure on butterfly distribution and movement: a review. J Insect Conserv 13:3–27
Dover J, Sparks T (2000) A review of the ecology of butterflies in British hedgerows. J Environ Manage 60:51–63
Dover JW, Sparks TH, Greatorex-Davies JN (1997) The importance of shelter for butterflies in open landscapes. J Insect Conserv 1:89–97
Ebert G, Rennwald E (eds) (1991) Die Schmetterlinge Baden Württembergs, Band 2, Tagfalter II.E. Ulmer, Stuttgart
Ellenberg H (1974) Zegerwerte der Gefäßpflanzen Mitteleuropas. Scripta Geobotan 9:1–97
Erhardt A (1985) Diurnal lepidoptera: sensitive indicators of cultivated and abandoned grassland. J Appl Ecol 22:849–861
Fahrig L (2003) The effects of habitat fragmentation on biodiversity. Annu Rev Ecol Syst 4:487–515
Głowaciński Z, Nowacki J (eds) (2004) Polska Czerwona Księga Zwierząt. Instytut Ochrony Przyrody PAN, Kraków, Akademia Rolnicza, Poznań
Grill A, Cleary DFR, Stettmer C, Bräu M, Settele J (2008) A mowing experiment to evaluate the influence of management on the activity of host ants of Maculinea butterflies. J Insect Conserv 12:617–627
Güsewell S (2003) Management of Phragmites australis in Swiss fen meadows by mowing in early summer. Wetl Ecol Manag 11:433–445
Hahn IJ, Vergara PM, Römer U (2011) Habitat selection and population trends in terrestrial bird species of Robinson Crusoe Island: habitat generalists versus forest specialists. Biodivers Conserv 20:2797–2813
Hall LS, Krausman PR, Morrison ML (1997) The habitat concept and a plea for standard terminology. Wildlife Soc B 25:173–182
Hardy PB, Dennis RLH (2007) Seasonal and daily shifts in substrate use by settling butterflies: conserving resources for invertebrates has a behavioral dimension. J Insect Behav 20:181–199
Johst K, Drechsler M, Thomas J, Settele J (2006) Influence of mowing on the persistence of two endangered large blue butterfly species. J Appl Ecol 43:333–342
Konvicka M, Benes J, Kuras T (2002) Microdistribution and diurnal behaviour of two sympatric mountainous butterflies (Erebia epiphron and E. euryale): relations to vegetation and weather. Biologia 57:221–233
Krauss J, Steffan-Dewenter I, Tscharntke T (2004) Landscape occupancy and local population size depends on host plant distribution in the butterfly Cupido minimus. Biol Conserv 120:355–361
Krzywicki M (1982) Monografia Motyli Dziennych Polski. Papilionidea i Hesperioidea, Lublin
Kudrna O (2002) The distribution atlas of european butterflies. Oedippus 20:1–343
Kugler H (1955) Einführung in die Blütenökologie. G. Fischer, Jena
Lenda M, Skórka P (2010) Patch occupancy, number of individuals and population density of the Marbled White in a changing agricultural landscape. Acta Oecol 36:497–506
Lepš J, Šmilauer P (2003) Multivariate analysis of ecological data using CANOCO. Cambridge University Press, Cambridge, UK
Mayberry RJ, Elle E (2010) Conservation of a rare plant requires different methods in different habitats: demographic lessons from Actaea elata. Oecologia 164:112–130
Mildenstein T, Stier S, Nuevodiego C, Mills L (2005) Habitat selection of endangered and endemic large flying-foxes in Subic Bay, Philippines. Biol Conserv 126:93–102
Nowicki P, Witek M, Skórka P, Settele J, Woyciechowski M (2005) Population ecology of the endangered butterflies Maculinea teleius and M. nausithous and the implications for conservation. Popul Ecol 47:193–202
Nowicki P, Pepkowska A, Kudlek J, Skórka P, Witek M, Settele J, Woyciechowski M (2007) From metapopulation theory to conservation recommendations: lessons from spatial occurrence and abundance patterns of Maculinea butterflies. Biol Conserv 140:119–129
Parker GA (1992) Marginal value theorem with exploitation time costs: diet, sperm reserves, and optimal copula duration in dung flies. Am Nat 139:1237–1256
Parker GA, Stuart RA (1976) Animal behavior as a strategy optimizer: evolution of resource assessment strategies and optimal emigration thresholds. Am Nat 110:1055–1076
Potts SG, Dafni A, Ne’eman G (2001) Pollination of a core flowering shrub species in Mediterranean phrygana: variation in pollinator diversity, abundance and effectiveness in response to fire. Oikos 92:71–80
Powell KI, Chase JM, Knight TM (2011) A synthesis of plant invasion effects on biodiversity across spatial scales. Am J Bot 98:539–548
Rosin ZM, Myczko Ł, Skórka P, Lenda M, Moroń D, Sparks TH, Tryjanowski P (2012) Butterfly responses to environmental factors in fragmented calcareous grasslands. J Insect Conserv 16:321–329
Schwarzwälder B, Lörtscher M, Erhardt A, Zettel J (1997) Habitat utilization by the heath fritillary butterfly, Mellicta athalia ssp. celadussa (Rott.) (Lepidoptera: Nymphalidae) in montane grasslands of different management. Biol Conserv 82:157–165
Settele J, Feldmann R, Reinhardt R (eds) (1999) Die tagfalter deutschlands. Ulmer, Stuttgart
Shreeve TG, Dennis RLH, Roy DB, Moss D (2001) An ecological classification of British butterflies: ecological attributes and biotope occupancy. J Insect Conserv 5:145–161
Skórka P, Martyka R, Wójcik JD (2006) Species richness of breeding birds at a landscape scale—which habitat is the most important? Acta Ornithol 41:49–54
Skórka P, Settele J, Woyciechowski M (2007) Effects of management cessation on grassland butterflies in southern Poland. Agr Ecosyst Environ 121:319–324
Slamova I, Klecka J, Konvicka M (2011) Diurnal behavior and habitat preferences of Erebia aethiops, an aberrant lowland species of a mountain butterfly clade. J Insect Behav 24:230–246
Sparks TH, Parish T (1995) Factors affecting the abundance of butterflies in field boundaries in Swavesey fens, Cambridgeshire, UK. Biol Conserv 73:221–227
Ter Braak CJF, Prentice CI (1988) A theory of gradient analysis. Adv Ecol Res 18:217–317
Timonen J, Gustafsson L, Kotiaho JS, Mönkkönen M (2011) Hotspots in cold climate: conservation value of woodland key habitats in boreal forests. Biol Conserv 144:2061–2067
Turlure C, Van Dyck H, Schtickzelle N, Baguette M (2009) Resource-based habitat definition, niche overlap and conservation of two sympatric glacial relict butterflies. Oikos 118:950–960
Van Swaay C, Cuttelod A, Collins S, Maes D, Lopez Munguira M, Šašić M, Settele J, Verovnik R, Verstrael T, Warren M, Wiemers M, Wynhof I (2010) European red list of butterfies. Publications Office of the European Union, Luxembourg
Vanreusel W, Van Dyck H (2007) When functional habitat does not match vegetation types: a resource-based approach to map butterfly habitat. Biol Conserv 135:202–211
Vilà M, Espinar JL, Hejda M, Hulme PE, Jarošík V, Maron JL, Pergl J, Schaffner U, Sun Y, Pyšek P (2011) Ecological impacts of invasive alien plants: a meta-analysis of their effects on species, communities and ecosystems. Ecol Lett 14:702–708
WallisDeVries MF, Poschlod P, Willems JH (2002) Challenges for the conservation of calcareous grasslands in northwestern Europe: integrating the requirements of flora and fauna. Biol Conserv 104:265–273
Warecki A, Sielezniew M (2008) Dryad Minois dryas (Lepidoptera, Nymphalidae) in south-eastern Poland: a recent range expansion or oversight of an endangered species? Pol J Entomol 77:191–198
Wynhoff I, Gestel R, Swaay C, Langevelde F (2011) Not only the butterflies: managing ants on road verges to benefit Phengaris (Maculinea) butterflies. J Insect Conserv 15:189–206
Zarzycki K, Trzcińska-Tacik H, Różański W, Szeląg Z, Wołek J, Korzeniak U (2002) Ecological indicators values of vascular plants of Poland. Szafer Institute of Botany, Polish Academy of Sciences, Kraków
Acknowledgments
The authors are grateful to the students of the Jagiellonian University and the volunteers who participated in the fieldwork. Thanks are also due to Magda Lenda for her help with identification of some plant species. Two anonymous referees provided useful critical comments on earlier version of the manuscript. The study was funded by the Polish National Science Centre grant No. N304 064139 and by the German Federal Ministry of Education and Research under its FP6 BiodivERsA Eranet project CLIMIT. Fundacja Studentów i Absolwentów UJ “Bratniak” (the Jagellonian University Students and Graduates’ Bratniak Foundation) and Rada Kół Naukowych UJ (the Council of the Jagellonian University Sciences Circle) also offered financial support for conducting the fieldwork.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Kalarus, K., Skórka, P. & Nowicki, P. Resource use in two contrasting habitat types raises different challenges for the conservation of the dryad butterfly Minois dryas . J Insect Conserv 17, 777–786 (2013). https://doi.org/10.1007/s10841-013-9560-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-013-9560-1