
Abstract
Accurate assessment of postfire vegetation recovery is important for forest management and the conservation of species diversity. Topography is an important factor affecting vegetation recovery but whether species composition varies with different recovery stages and between valleys and slopes is unclear. Using field data and a space-for-time substitution method, we quantified species richness and diversity to obtain the successional trajectories of valleys and slopes. We surveyed the species of 10 burned areas from 1986 to 2010 in the Greater Khingan Mountains in northeastern China, and found that with increasing postfire recovery time, species richness in both valleys and slopes gradually decreased. However, species richness in valleys was relatively higher. Shrubs recovered rapidly in the valleys, and species diversity maximized approximately 11 years after fire. However, it maximized 17–18 years after fire on the slopes. Numerous shade-tolerant species were present in the valleys 11 years after fire but not until after 18 years on slopes. Larch appeared earlier than 11 years after fire and its recovery was slow in the valleys but appeared quickly on slopes and established dominance early. Our study provides some new insights into vegetation succession after fire at local scales. After fire, the vegetation recovery processes differ with topography and it affects the initial rate of recovery and species composition at different successional stages.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wildfires are one of the most destructive natural disasters of forest ecosystems and hundreds of thousands occur annually around the world (Ubeda and Sarricolea 2016; Yue et al. 2020). They result in not only severe loss of resources but increase spatial heterogeneity of the forest environment, alter succession and composition, and reshape landscape diversity, changes that have an impact on vegetation regeneration and recovery (Wang et al. 2004; Xing et al. 2006; Boucher et al. 2020; Moradizadeh et al. 2020). Vegetation growth and recovery in burned forests are affected by solar radiation, hydrological conditions, soil nutrients, and other factors (Liu et al. 2017; Whitman et al. 2018a; Zhang et al. 2021a). Accurate evaluation of postfire vegetation recovery using species diversity indicators (Gonzalez-Tagle et al. 2008), especially under different topographic conditions, provides a scientific basis for forest decision-making and management (Francos et al. 2020; Liu et al. 2020; Guo et al. 2021; Oettel and Lapin 2021).
In postfire vegetation recovery studies, field surveys, remote sensing, or a combination of both have been used to investigate species change and the direction of vegetation recovery at multiple scales (Kilgore and Dan 1979; Wang et al. 2004; Bright et al. 2019; Moya et al. 2020; Stinca et al. 2020; Smith-Ramirez et al. 2022). Smith-Ramirez et al. (2021) found that frequent fires made it difficult for vegetation to recover to its pre-fire state, even after more than 20 years. Fire severity and recovery time affect the distribution and composition of understory vegetation (Mathews and Kinoshita 2021; Weber et al. 2022). After low-severity fires, the vegetation becomes more homogeneous over time (Bataineh et al. 2006; Sun and Hu 2010; Shive et al. 2013; Harris and Taylor 2020), but high-intensity fires have a fundamental impact on vegetation composition and may change the direction of postfire succession (Han et al. 2015a, b; Meng et al. 2015; Moradizadeh et al. 2020). Moreover, climate is closely related to postfire vegetation recovery (Liu 2016; Hamberg et al. 2020; Guo et al. 2022). Bright et al. (2019) found a close relationship between postfire NBR (normalized burn ratio) and postfire precipitation and temperature. Prolonged drought can lead to the change of forest vegetation to open shrublands after fire (Baudena et al. 2020). Whitman et al. (2018a) found that climate and topography were the main factors in community change after fire in boreal coniferous forests. However, in postfire Siberian larch forests, Thuan et al. (2017) concluded that the main controlling factors for larch recovery were the extent of the fire and soil moisture content, and larch recovery rate was higher under a moderate fire. Smith-Ramirez et al. (2022) found that woody species richness could reach pre-fire levels after 20 years with no other intervention.
Remote sensing techniques have been used to produce vegetation indices, including NDVI (normalized differential vegetation index) and EVI (enhanced vegetation index), to represent the “greenness” of vegetation recovery at a large scale (Vasilakos et al. 2018; Shvetsov et al. 2019; Barmpoutis et al. 2020; Lacouture et al. 2020). Yang et al. (2017) found that NDVI recovery time would be longer than estimated by existing studies. However, there are limitations in using NDVI to monitor vegetation recovery, as it does not indicate species change and interspecific competition. Moreover, the index may reach “saturation” when vegetation cover density is high. For example, Guo et al. (2021) used sun-induced chlorophyll fluorescence (“SIF”) and EVI to compare dynamic vegetation recovery changes in postfire vegetation in northern China and found that 20 years after the fire was insufficient for vegetation recovery because SIF was still increasing. Compared to field investigations, Guo et al. (2021) found that EVI reached a pre-fire level but the vegetation was still recovering.
Postfire vegetation recovery is affected by topography, soil, and fire intensity (Näsholm et al. 2013; Han et al. 2015a, b; Kong et al. 2018; Whitman et al. 2018a, b), factors that influence the rate and direction of vegetation succession (Hart and Chen 2006; Liu et al. 2017; Gavito et al. 2021; Li et al. 2021). Changes in surface temperature and soil properties can affect seedling development (Bonan and Shugart 1989; Stefanski 2022; Vassilis et al. 2022). Furthermore, secondary succession can occur (Shvetsov et al. 2019; Stinca et al. 2020), and vegetation responsiveness influences recovery (Brooker et al. 2008; Gao et al. 2022). However, soil nutrients and moisture influence vegetation response (Veihmeyer and Hendrickson 1950; Adagbasa et al. 2020; Jiao et al. 2021); for example, they drive gully vegetation recovery processes (Zhang et al. 2021b; Zhu et al. 2021). However, topography and fire severity are the main drivers of postfire vegetation recovery (Ireland and Petropoulos 2015; Thuan et al. 2017; Whitman et al. 2018a). Topographic factors are important predictors of postfire forest patterns because they alter the hydrology conditions for vegetation growth and influence vegetation composition (Kong et al. 2003; Hart and Chen 2006; Cai et al. 2012; Liu et al. 2017; Moradi and Vacik 2018; Boag et al. 2020). Marler and del Moral (2018) found that slopes differed in light and water qualities, which changed species composition. Bassett et al. (2017) reported that topography caused regional vegetation structure differences. Shrub and fern cover in valleys was higher, and more complex than on slopes.
The Greater Khingan Mountains have ecological importance for China’s economic and social development (Zheng et al. 1986; Li 2004; Song et al. 2010). Wildfire is the most predominant natural disturbance and causes a serious loss of resources (Fang et al. 2021). To accurately assess boreal forest successional processes, studies have been carried out on vegetation composition and species diversity changes at community, ecosystem, and landscape scales (Huang et al. 2020b), monitoring dynamic changes in large-scale vegetation recovery, and exploring factors affecting them (Shu et al. 1999; Kong 2003; Yang et al. 2013; Han et al. 2015a, b; Wang et al. 2021). Additionally, researchers have found that vegetation recovery varies with fire intensity. Vegetation recovery is most pronounced following moderate fire disturbance (Sun and Hu 2010; Wei et al. 2016). However, there has been no quantitative research on species changes during vegetation recovery under the influence of topography in the Greater Khingan Mountains. This is also true for postfire succession comparisons between valleys and slopes. Essentially, it is unclear what, if any, are differences in species composition and population changes in postfire vegetation succession between slopes and valleys (Cai et al. 2018). Moreover, the diversity of understory vegetation plays an important role in maintaining ecosystem productivity. Differences in soil moisture and nutrients between valleys and slopes affect vegetation diversity and understanding species composition and diversity differences between valleys and slopes can enrich forest management system science, important for species diversity conservation and improving ecosystem productivity (Yang et al. 2013).
Because of hydrothermal differences due to topography, it is assumed that vegetation recovery to a pre-fire state will be faster in valleys than on slopes. For this study, 10 burned and 2 unburned areas (reference samples) from 1986 to 2010 were selected in the Greater Khingan Mountains. Based on a space-for-time substitution method, it was hypothesized that all burned and unburned areas had similar site conditions (including altitude, slope, vegetation cover, and in the case of burned areas, fire intensity), and formed a chronosequence to study species composition and diversity trends in valleys and on slopes. The study aims to: (1) quantify and compare changes in species diversity during vegetation recovery after fire in valleys and on slopes of the Greater Khingan Mountains; and (2) analyze changes in species composition in succession and determine vegetation recovery time to a pre-fire state.
Materials and methods
Study region
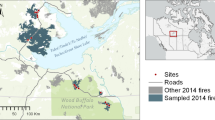
The study region is in the Greater Khingan Mountains of northeastern China (48.56°–51.45° N, 121.34°–123.41° E) across Inner Mongolia and Heilongjiang Province (Fig. 1). The area is a coniferous forest region and is the southern extension of the Eurasian coniferous zone. It has a cold temperate continental climate, with long, dry winters and short, humid summers. The average annual temperature is − 2.8 °C, and average annual precipitation 460 mm (Wen et al. 2021b). Zonal vegetation is mixed coniferous and broad-leaved forest dominated by larch and birch. Understory shrubs are mainly Rhododendron dauricum L., Vaccinium uliginosum L. Betula fruticosa Pallas, Vaccinium vitis-idaea L., and Rosa davurica Pallas (Wu 1982; Han et al. 2015a, b; Wang 2016a).

Study area and selected burn locations surrounding the Landsat images (the same year or 1 year after the fire) of each burn scar; the Landsat image of burned area h is not shown because it was very close to g
Precipitation is relatively low and vegetation burns readily; thus, wildfires are the main natural disturbance in the region (Guo et al. 2021). The annual average number of fires exceeds 35, and the average area burnt is 76,600 ha, usually by predominantly moderately intense fires, with few fires of low and high intensity. Topography, climate, and vegetation types tend to cause spatial heterogeneity of fire intensity (Liu 2011; Huang et al. 2020a; Yuan 2020). Due to the relatively simple vegetation composition, the area is ideal for studying postfire vegetation recovery processes. We selected 10 different-aged burned areas (1986–2010) in the Greater Khingan Mountains. The locations of burned sites, fire dates, and postfire Landsat images are shown in Fig. 1.
Field survey data
In the summers of 2020 and 2021, the 10 burned and 2 unburned areas were surveyed (detailed information in Supplementary Table S1, and fire intensities of the 10 burned areas shown in Fig. 2). Since the vegetation types in the area are dominated by larch and birch and habitats basically similar, two controls were selected and tree ages between 80 to 120 years determined. It was assumed that the control sites were a fire-origin site 100 years after a fire in the valley or slope (Sun and Hu 2010).

Spatial distribution of fire intensity in 10 burned areas. Sample point locations were obtained with photos taken by GPS cameras during the sampling process
Vegetation recovers rapidly and grows well after moderate fires and burn patches are ideal areas to study postfire vegetation recovery (Roberts 2004; Sun and Hu 2010; Yang et al. 2013). In each burned area, three 20 m × 20 m tree quadrats and six 5 m × 5 m shrub quadrats were established in moderate fire patches in valleys and on slopes according to the composite burn index (Wang et al. 2013). Species number, plant number, plant DBH (diameter at breast height) and plant crown width were recorded. Typical field survey sites of forests/shrublands in valleys and on slopes are shown in Fig. 3.

Photos of areas burned in 1986 (a valley, b slope; photo taken in 2021), 2003 (c valley, d slope; photo taken in 2020) and 2006 (e valley, f slope, photo taken in 2021)
In addition, the differential normalized burn ratio (“dNBR”) was calculated, and three fire intensities set (Fig. 3) using the TOA (top-of-atmosphere reflectance) threshold method (Fang 2014). In subsequent data processing, we excluded sample point data at other fire intensities.
Ecological indices
Five indices were selected for analysis (Zhang 2004, 2007; Wang et al. 2015). The equations for the indices (1–5) and ecological significance (Table 1) are as follows:
where Y is the importance value, a is the relative abundance, b is the relative frequency, and c is the relative significance.
Shannon–Wiener diversity index (H):
where \({P}_{i}={n}_{i}/N\), represents the relative abundance of the ith species; \({n}_{i}\) is the number of individuals of species \(i\); N is the total number of individuals of all species in the community.
Simpson diversity index (D):
where \({N}_{i}\) is the number of species \(i\) and \(N\) the number of individuals of all species in the community.
Pielou evenness index (J):
where \(S\) is the number of species in the sample and H is the Shannon–Wiener diversity index.
Margalef richness index:
where \(N\) is the number of individuals of all species in the community and \(S\) the number of species.
Space-for-time substitution method
This method is primarily used to study slow/long-term ecological processes, and widely used in ecology and geomorphology using spatial data to replace different stages of the process (Ma et al. 2017; Damgaard 2019; Huang et al. 2020a).
Normally areas at different successional stages in regions with similar environments are selected. The terrain in the Greater Khingan Mountains is less undulating (Han et al. 2015a, b) with mainly brown coniferous forest soil, which is distributed on slopes dominated by larch, and swampy soil distributed in the valley lowlands where there is more birch and less larch (Zhao 2009; Chen et al. 2012). However, in general the regional vegetation types are similar (Guo et al. 2020). In this study, we selected several moderately burned areas from different fire years for sampling. As noted previously, the study areas had similar elevations, slopes, and aspects and it was assumed that moderate fire scars on valleys and slopes were homogeneous. Field sampling data in different years constituted the valley and slope vegetation recovery sequences.
Data analysis
All analyses were conducted using Microsoft Excel 2016 and RStudio (the “vegan” package). One hundred and eight 108 shrubs and 54 tree quadrats were surveyed within 18 sample plots in the 10 burned areas and two unburned areas. Mean species number, vegetation canopy width and tree DBH for each quadrat were recorded to represent vegetation recovery information (Wang et al. 2004). The species diversity index of each sample plot was calculated using RStudio (Weinhold et al. 2021). Quadrat species importance values were determined using methods described above. Species richness and species diversity of the 10 valleys and eight slope plots were calculated, and species compositional changes for valleys and slopes at six recovery stages were analyzed.
Results
Species richness changes
Shrub and tree species numbers changed over time in valleys and on slopes (Figs. 4 and 5). With increasing recovery time, there was a decrease in the total number of species in the community and the total number of shrub species. Tree species richness did not change drastically; the most common species were Larix gmelinii and Betula platyphylla. The maximum number of total species and shrub species in both valleys and slopes occurred 14-year postfire, and there were no significant differences between valleys and slopes. The minimum number of total species and shrub species in valleys occurred 100-year postfire, and 15-year postfire on slopes. For the slopes, this was mainly due to the large, burned areas affecting species migration and to the vegetation having not fully recovered after the previous fire disturbance. It appears that species moved into the burned slope areas before the peak years, followed by competition until species populations stabilized.

Variation in species richness over time in postfire valleys

Variation in species richness over time on postfire slopes
Species diversity index changes
To further understand the characteristics of community change in burned areas by recovery years, the species diversity index of the valleys and slopes in different recovery years were calculated (Fig. 6).

Changes in species diversity over time in postfire valleys and slopes: a, a' Shannon–Wiener diversity index; b, b' Simpson diversity index; c, c' Pielou evenness index; and d, d' Margalef richness index
With increasing recovery time in valleys, the Shannon–Wiener diversity, Simpson diversity, and Pielou evenness indices all showed a decrease (Fig. 6a–c), indicating that the degree of species diversity declined, and species distribution was uneven. However, the Margalef richness index increased initially, then decreased and followed the same pattern with increasing recovery time, indicating marked fluctuations in the number of species over time.
In contrast, with increasing recovery time on slopes, the four indices showed an overall trend of initially increasing and then decreasing (although some individual years did not match the overall trend) (Fig. 6a'–d'), indicating marked initial increases in community species richness and diversity and then a decline. However, as discussed in the previous section, species change at 15-year postfire was unique, which led to a difference in the Margalef richness index from the other indices.
Overall, community diversity reached its maximum at approximately 11–14-year postfire in valleys and gradually decreased with recovery time, while it was 17 or 18 years after the fire on slopes.
Species importance value changes
To quantify the dynamic change in species vegetation at different recovery times in the valleys, the importance value of recovered species was calculated (Table 2). From 11- to 35-year postfire, shade-tolerant shrubs such as Vaccinium vitis-idaea, V. uliginosum, Salix rosmarinifolia, and Betula fruticosa were continuously present and increased. Light-demanding shrubs such as Rhododendron dauricum and Rosa davurica gradually decreased or disappeared entirely. The transition from light-demanding to shade-tolerant shrubs indicates an increase in canopy density and a gradual recovery of the tree layer (Wang et al. 2004).
In the initial stage of postfire vegetation recovery, shrubs rapidly appeared and became abundant (Table 2). At the same time, tree recovery was slow. Larch began to dominate 30-year postfire while shrubs were relatively few.
The importance values of species vegetation at different recovery times on slopes were calculated (Table 3). From 11- to 35-year postfire, shade-tolerant shrub species such as Ledum palustre, Spiraea salicifolia Vaccinium vitis-idaea and V. uliginosum appeared and became dominant. However, light-demanding species with low importance values such as Rosa davurica were less dominant.
From 11- to 35-year postfire, larch and B. platyphylla were early abundant species and larch was dominant in the middle and late recovery stages. Generally, larch and B. platyphylla were abundant 11-year postfire, accompanied by a small amount of Populus davidiana. With increasing recovery time, larch increasingly dominated the canopy layer accompanied by some B. platyphylla and Pinus sylvestris. Unlike in the valleys, trees on the slopes such as larch and B. platyphylla dominated in the early stages of postfire recovery, while shrub species were relatively scarce.
To more vividly and intuitively understand species succession in valleys and on slopes, conceptual figures based on our results, analyses and field surveys were drawn (Figs. 7 and 8). Probable years for different successional stages were evaluated. In the early stages of valley vegetation succession, tall shrubs dominated. With increasing recovery time, they were gradually replaced by larch and birch, accompanied by a small number of low shrubs. However, in the early stage of slope vegetation succession, larch and birch existed in large numbers. With increasing recovery time, other tall shrubs began to appear and gradually decreased. Finally, a mixed forest of larch and birch formed accompanied by some low shrubs.

Species succession in valleys

Species succession on slopes
Effect of slope on postfire vegetation recovery
Slope changes light, soil moisture, and nutrient conditions (Hart and Chen 2006; Han et al. 2015a, b; Liu et al. 2017), which affects vegetation growth. For the same burned area, there were significant vegetation recovery differences between slopes and valleys. Therefore, we selected six burned scars with both slopes and valleys and compared species richness and diversity (Fig. 9).

Changes in species richness and diversity postfire in valleys and slopes in the same burn
The 17-year postfire was the cutoff point in species richness changes in valleys and slopes from Fig. 9a. Initially, species richness in the valleys was higher than on slopes, but 17-year postfire it was the opposite. Species diversity (as measured by the four indices) in the valleys was higher than on slopes to 17 years, but after 17 years, this pattern was reversed. Since species richness determines community richness, slope richness was higher than in the valleys 17-year postfire.
Figure 9 shows that slope affects the rate of vegetation succession. In the initial stage, shrubs rapidly established on the scar of the burn in valleys. Both species richness and diversity were significantly higher than in burns on slopes. However, with increasing time, species richness gradually increased on slopes. In the middle and later stages of vegetation recovery, species richness and diversity of valleys and slopes were similar, or marginally higher on slopes.
Discussion
Characteristics of postfire vegetation succession
After fire, the forest community re-established from shrubs and grass to trees, although the recovery pattern varied with the terrain. Quantifying the different patterns of vegetation recovery at different successional stages is important for understanding the recovery process.
Several researchers have found that postfire, species richness initially increases, then decreases and stabilizes over time (Wang et al. 2004; Han et al. 2015a, b). Moreover, species composition transitions from light-demanding to shade-tolerant shrubs (Song et al. 2010; Yang et al. 2013; Liu et al. 2017). Shrub and herb recovery is rapid but tree recovery is slow (Wang et al. 2004; Han et al. 2015a, b). Our results confirm these patterns. However, we further considered the impact of topographical factors on vegetation recovery and arrived at some new findings. Species such as larch recovered faster than shrubs on slopes. Conversely, shrub species flourished in valleys where moisture conditions were more favorable (Zhao 2009; Wang 2016b). Previous studies of postfire vegetation succession have generally focused on regional or landscape scales (Vasilakos et al. 2018; Lacouture et al. 2020; Guo et al. 2021), while neglecting the local scale which can more accurately reflect succession patterns.
The speed and direction of vegetation succession after fire disturbance depend on the degree and duration of environmental factors affecting vegetation growth (Cai et al. 2012; Zong and Tian 2022). Light, water, and soil fertility are necessary for vegetation growth and important determinants of species composition, soil nutritional status, mobilization of elements and organic matter turnover and can change soil fertility and thus affect vegetation growth and development (Mao and Zhu 2006; Zhu et al. 2021). Fire disturbances can consume soil organic matter, improve soil structure and composition (Kong et al. 2018), and increase nutrient return rate (Yang et al. 2013). Conversely, fires burn large areas of vegetation (Wang et al. 2004; Yue et al. 2020), which reduce canopy density and improve light transmittance (Hart and Chen 2006; Cai et al. 2012; Yang et al. 2013).
At the initial stage of postfire recovery, many understory species rapidly colonized and thrived due to sufficient light and rich soil nutrients. Biomass turnover was high, thus favoring stable and durable light-demanding shrubs (Liu et al. 2017). However, the fundamental condition for vegetation growth is a supply of sufficient seed and subsequent seed germination (Liu et al. 1995). Studies have shown that a moderate fire has little effect on seeds (Sun and Hu 2010). Also, larch seed supply is sufficient (Zhou 1988; Shan 1996). However, a larger burnt area may cause a shortage of seed sources. Suitable temperature and humidity are required for seed germination (Ibáñez et al. 2022). In valleys with deep organic layers (Bassett et al. 2017), there is sufficient moisture but a thick moss layer on the surface (shown through field investigation), making it difficult for larch and birch seeds to germinate (Bonan and Shugart 1989; Ibáñez et al. 2022; Kilpeläinen et al. 2022). Therefore, in the initial stage of vegetation recovery after fire, shrubs dominate (Wang 2016a). As recovery time increased, tree species gradually established (Wang et al. 2004), canopy density increased as trees occupied a dominant position, and shrub species began to decline, leading to a gradual decline in overall species richness (Zhao 2009; Han et al. 2015a, b).
Slope affects soil moisture and nutrients and changes the conditions for growth, which leads to differences in vegetation recovery between valleys and slopes (Luo 2002; Liu et al. 2017). Shrubs thrive when moisture is abundant and the soil rich in nutrients (Bassett et al. 2017), while larch is adaptable and can grow well where soil moisture and nutrient conditions are poor, even in permafrost (Qu et al. 2022). Shrubs and B. platyphylla are pioneer species in the early stages of vegetation recovery (Wang 2016a). B. platyphylla has adequate seed production, long-distance seed dispersal, and good sexual reproduction (Shi et al. 2014). Larch however, produces fewer seeds and grows slowly when young (Wang et al. 2004). Therefore, it was expected that shrub establishment and growth in the valleys to be rapid, B. platyphylla to be present and larch to be absent. However, in the survey, birch as a pioneer species in moist valleys was rare, and most were shrubs such as bush-birch. The possible reason for this is that the thickness of the active permafrost layer in the Greater Khingan Mountains is shallow and impedes nutrient absorption by B. platyphylla roots, thereby hindering growth (Wen et al. 2021a). In the field survey, we also found that in some low elevation valleys, there was no B. platyphylla stage, succession went from shrub to larch directly because of abundant permafrost. On slopes, water and soil nutrients are lower, shrub growth is limited, and B. platyphylla and larch quickly establish (Yang et al. 2013; Liu et al. 2020). On slopes, forests dominated by larch developed more quickly, although the change in shrub species was relatively slow (Fig. 10).

a abundant larch seedlings on a burnt slope in 2006; b abundant Betula middendorfii and B. fruticosa in a burnt valley in 2006. Photos taken in 2021
Topography generally led to an unsynchronized process of vegetation succession, but species composition of valleys and slopes was similar in the later stages as both were dominated by larch and birch.
Diversity characteristics of vegetation succession
The recovery of species richness and diversity is an important factor and indicator of ecosystem recovery and development (Wang et al. 2021). With increasing species diversity, community stability and productivity increased (Wang et al. 2015) and forest ecosystems gradually recovered. Researchers have found that species diversity is dynamic in postfire vegetation recovery (Wang et al. 2004, 2015; Han et al. 2015a, b). Generally, as recovery time increases, species diversity initially increases, decreases, and then stabilizes (Song et al. 2010). Wang (2016a) found that the community diversity index maximized 20-year postfire. In this study, species diversity indices of burned areas on slopes initially increased and then declined, a similar trend to that of previous studies (Li and Shao 2004; Yang 2019); however, the time to attain maximum species diversity was different. Our results indicate that valleys reach their maximum approximately 11- and 17-year postfire for slopes.
Dynamic changes in species diversity during postfire vegetation recovery were closely related to the growth environment. Initially, after fire, light, water, and soil nutrients were sufficient (Wang et al. 2004, 2015; Yang et al. 2013); thus, shrubs rapidly colonized (Song et al. 2010; Liu et al. 2017). With increasing time, light, soil nutrients, and moisture became limited (Zong and Tian 2022). At this point, vegetation growth and development were hindered and interspecific competition became the limiting factor affecting community diversity (Chu et al. 2021). Interspecies competition can generate new niches to increase species diversity. Subject to the limits of the environment, in mid-late vegetation recovery, interspecific competition and intraspecific competition are intense, and vegetation enters a self-thinning stage (Bolnick et al. 2010; Jensen and Lof 2017), and community species diversity decreases.
Important value characteristics of vegetation succession
Species importance values reveal dominant changes at different successional stages and dynamic changes during vegetation recovery (Wang 2016a). Species changes depend on differences in environmental conditions for growth (Kong et al. 2003; Mao and Zhu 2006). Because of the different adaptabilities of vegetation to the environment, different dominant species appear in different successional stages after fire (Song et al. 2010). Research has shown that light-demanding species dominate the initial stages of vegetation recovery and shade-tolerant species (Vaccinium vitis-idaea, Salix floderusii, Betula fruticosa, appear and gradually dominate 8–9-year postfire (Wang et al. 2004; Mao and Zhu 2006; Sun 2011). Additionally, Wang et al. (2015) found that shade-tolerant species may appear or increase after fire for 20 years. However, these studies did not distinguish different topographies. In this study, light-demanding species gradually disappeared in both valleys and on slopes, while shade-tolerant species initially increased and then declined. The difference between valleys and slopes is that shade-tolerant species predominated 11- and 18-year postfire, respectively.
Vegetation recovery of burned areas is a long and gradual process (Kong et al. 2003) whereby shrub and grass communities gradually give way to communities dominated by larch (Zhao et al. 2013). Some researchers consider that larch only appears 16-year postfire, while others consider that it can recover as early as five years after fire, depending on the growth environment (Wang et al. 2004; Yang 2019). We documented the presence of larch and birch in valleys and slopes 11-year postfire. Larch recovered quickly and became dominant on the slopes in far greater numbers than in valleys. Water and soil nutrient conditions on slopes are unsuitable for the growth of many shrubs (Luo 2002; Liu et al. 2020) and thus provide an ecological niche for the growth of larch (Qu et al. 2022). The thin organic layer creates ideal conditions for the germination of larch seeds. B. platyphylla is a pioneer species that has small seeds, strong germination and is conducive to spreading. The sparse structure of B. platyphylla stands also creates conditions for larch regeneration (Han et al. 2015a, b; Sun 2011). In the early stages of vegetation recovery, larch and birch mixed forests readily appear, which are more obvious on slopes, while shrubs dominate in valleys.
Limitations
Vegetation succession after fire is a long-term process that requires many of years. We selected the longest recovery time as 35 years but did not find recovery steps. This may lead to errors in the recovery stage division and may not allow the appearance of some species.
Secondly, we only distinguished vegetation recovery between slopes and valleys and did not distinguish slope angles. Therefore, differences in vegetation recovery on different slope angles could not be determined.
Finally, we did not survey areas only lightly or severely burned, and only sampled moderately burned areas. Therefore, only general insights into vegetation recovery of burned areas can be provided.
In a subsequent study, vegetation recovery over a longer period will be examined and under different slope angles and burn intensities.
Conclusion
A chronosequence of 10 burned areas and two unburned areas from 1986 to 2010 in the Greater Khingan Mountains was established to study post-fire vegetation recovery in valleys and on slopes. Topography played an important role in successional processes, and although it did not change their direction, it did affect the initial rate of recovery and species composition at different successional stages. Species diversity in valleys and slopes initially increased and then decreased, with species diversity peaking earlier in valleys than on slopes. In the middle and late successional periods (35-year postfire), a stable community of larch and birch with a small number of low shrubs formed in both valleys and slopes. This study could enrich boreal forest management plans. However, due to the short and inconsistent intervals of the time series constructed, there may be differences in the time of vegetation recovery to different stages.
References
Adagbasa EG, Adelabu SA, Okello TW (2020) Development of post-fire vegetation response-ability model in grassland mountainous ecosystem using GIS and remote sensing. ISPRS J Photogramm Remote Sens 164:173–183. https://doi.org/10.1016/j.isprsjprs.2020.04.006
Barmpoutis P, Papaioannou P, Dimitropoulos K, Grammalidis N (2020) A review on early forest fire detection systems using optical remote sensing. Sensors (switzerland) 20(22):1–26. https://doi.org/10.3390/s20226442
Bassett M, Leonard SWJ, Chia EK, Clarke MF, Bennett AF (2017) Interacting effects of fire severity, time since fire and topography on vegetation structure after wildfire. For Ecol Manag 396:26–34. https://doi.org/10.1016/j.foreco.2017.04.006
Bataineh AL, Oswald BP, Bataineh MM, Williams HM, Coble DW (2006) Changes in understory vegetation of a ponderosa pine forest in northern Arizona 30 years after a wildfire. For Ecol Manag. https://doi.org/10.1016/j.foreco.2006.09.003
Baudena M, Santana VM, Jaime Baeza M, Bautista S, Eppinga MB, Hemerik L, Garcia Mayor A, Rodriguez F, Valdecantos A, Ramon VV, Vasques A, Rietkerk M (2020) Increased aridity drives post-fire recovery of Mediterranean forests towards open shrublands. New Phytol 225(4):1500–1515. https://doi.org/10.1111/nph.16252
Boag AE, Ducey MJ, Palace MW, Hartter J (2020) Topography and fire legacies drive variable post-fire juvenile conifer regeneration in eastern Oregon, USA. For Ecol Manag. https://doi.org/10.1016/j.foreco.2020.118312
Bolnick DI, Ingram T, Stutz WE, Snowberg LK, Lau OL, Paull JS (2010) Ecological release from interspecific competition leads to decoupled changes in population and individual niche width. Proc R Soc B Biol Sci 277(1689):1789–1797. https://doi.org/10.1098/rspb.2010.0018
Bonan GB, Shugart HH (1989) Environmental factors and ecological processes in boreal forests. Ann Rev Ecol Syst 1−28
Boucher D, Gauthier S, Thiffault N, Marchand W, Girardin M, Urli M (2020) How climate change might affect tree regeneration following fire at northern latitudes: a review. New for 51(4):543–571. https://doi.org/10.1007/s11056-019-09745-6
Bright BC, Hudak AT, Kennedy RE, Braaten JD, Khalyani AH (2019) Examining post-fire vegetation recovery with Landsat time series analysis in three western North American forest types. Fire Ecol. https://doi.org/10.1186/s42408-018-0021-9
Brooker RW, Maestre FT, Callaway RM, Lortie CL, Cavieres LA, Kunstler G, Michalet R (2008) Facilitation in plant communities: the past, the present, and the future. J Ecol 96(1):18–34. https://doi.org/10.1111/j.1365-2745.2007.01295.x
Cai WH, Yang J, Liu ZH, Hu YM, Liu SJ, Jing GZ, Zhao ZF (2012) Forest regeneration and its influencing factors after the fire in the Greater Khingan Mountains, Heilongjiang Province. Acta Ecol Sin. https://doi.org/10.5846/stxb201105030574
Cai WH, Liu ZH, Yang YZ, Yang J (2018) Does environment filtering or seed limitation determine post-fire forest recovery patterns in boreal larch forests? Front Plant Sci. https://doi.org/10.5846/stxb201105030574
Chen Q, Zhao YS, Xin Y, Han XC, Xia XY, Han CH (2012) Plant species diversity and correlation with soil factors after vegetation restoration in burned area of Great Xing’an Mountains. Dissertation, Northeast Forestry University, Harbin, China
Chu XL, Zhang QG, Buckling A, Castledine M (2021) Interspecific niche competition increases morphological diversity in multi-species microbial communities. Front Microbiol. https://doi.org/10.3389/fmicb.2021.699190
Damgaard C (2019) A critique of the space-for-time substitution practice in community ecology. Trends Ecol Evol 34(5):416–421. https://doi.org/10.1016/j.tree.2019.01.013
Fang L (2014) Spatial-temporal pattern and driving force of forest fire disturbance intensity in Greater Khingan Mountains. Dissertation, Shenyang Institute of Applied Ecology, Chinese Academy of Sciences, Shenyang, China
Fang KY, Yao QC, Guo ZT, Zheng B, Du JH, Qi FZ, Yan P, Li J, Ou TH, Liu JN, He MS, Trouet V (2021) ENSO modulates wildfire activity in China. Nat Commun. https://doi.org/10.1038/s41467-021-21988-6
Francos M, Pereira P, Ubeda X (2020) Effect of pre- and post-wildfire management practices on plant recovery after a wildfire in Northeast Iberian Peninsula. J for Res 31(5):1647–1661. https://doi.org/10.1007/s11676-019-00936-7
Gao W, Zheng C, Liu X, Lu Y, Chen Y, Wei Y, Ma Y (2022) NDVI-based vegetation dynamics and their responses to climate change and human activities from 1982 to 2020: a case study in the Mu Us Sandy Land, China. Ecol Indic. https://doi.org/10.1016/j.ecolind.2022.108745
Gavito ME, Paz H, Barragan F, Siddique I, Arreola-Villa F, Pineda-Garcia F, Balvanera P (2021) Indicators of integrative recovery of vegetation, soil and microclimate in successional fields of a tropical dry forest. For Ecol Manag. https://doi.org/10.1016/j.foreco.2020.118526
Gonzalez-Tagle MA, Schwendenmann L, Perez JJ, Schulz R (2008) Forest structure and woody plant species composition along a fire chronosequence in mixed pine-oak forest in the Sierra Madre Oriental, Northeast Mexico. For Ecol Manag 256(1/2):161–167. https://doi.org/10.1016/j.foreco.2008.04.021
Guo M, Li J, Huang SB, Wen LX (2020) Feasibility of using MODIS products to simulate sun-induced chlorophyll fluorescence (SIF) in boreal forests. Remote Sens 12(4):680. https://doi.org/10.3390/rs12040680
Guo M, Li J, Yu FB, Yin S, Huang SB, Wen LX (2021) Estimation of post-fire vegetation recovery in boreal forests using solar-induced chlorophyll fluorescence (SIF) data. Int J Wildland Fire 30(5):365–377. https://doi.org/10.1071/WF20162
Guo BY, Wang JS, Mantravadi VS, Zhang L, Liu GZ (2022) Effect of climate and ecological restoration on vegetation changes in the “Three-River Headwaters” region based on remote sensing technology. Environ Sci Pollut Res 29(11):16436–16448. https://doi.org/10.1007/s11356-021-16927-3
Hamberg LJ, Fraser RA, Robinson DT, Trant AJ, Murphy SD (2020) Surface temperature as an indicator of plant species diversity and restoration in oak woodland. Ecol Indic. https://doi.org/10.1016/j.ecolind.2020.106249
Han FL, Bu RC, Chang Y, Hu YM, Ma J, Nie ZW (2015a) Dynamics of recovery process of understory vegetation of Betula platyphylla-Larix gmelinii forest in Daxing’an Mountains after fire disturbance. Chin J Ecol 34(2):312–318
Han J, Shen Z, Ying L, Li G, Chen A (2015b) Early post-fire regeneration of a fire-prone subtropical mixed Yunnan pine forest in Southwest China: effects of pre-fire vegetation, fire severity and topographic factors. For Ecol Manag 356:31–40. https://doi.org/10.13292/j.1000-4890.2015.0043
Harris LB, Taylor AH (2020) Rain-shadow forest margins resilient to low-severity fire and climate change but not high-severity fire. Ecosphere. https://doi.org/10.1002/ecs2.3258
Hart SA, Chen HYH (2006) Understory vegetation dynamics of North American boreal forests. Crit Rev Plant Sci 25(4):381–397. https://doi.org/10.1080/07352680600819286
Huang SB, Wen LX, Yin S, Guo M, Yu FB (2020a) Decay rate of Larix gmelinii coarse woody debris on burned patches in the Greater Khingan Mountains. J for Res 32(6):2349–2363. https://doi.org/10.1007/s11676-020-01270-z
Huang ZB, Cao CX, Chen W, Xu M, Dang YF, Singh RP, Bashir B, Xie B, Lin XJ (2020b) Remote sensing monitoring of vegetation dynamic changes after fire in the Greater Hinggan Mountain area: the algorithm and application for eliminating phenological impacts. Remote Sens. https://doi.org/10.3390/rs12010156
Ibáñez TS, Wardle DA, Gundale MJ, Nilsson MC (2022) Effects of soil abiotic and biotic factors on tree seedling regeneration following a Boreal forest wildfire. Ecosystems 25(2):471–487. https://doi.org/10.1007/s10021-021-00666-0
Ireland G, Petropoulos GP (2015) Exploring the relationships between post-fire vegetation regeneration dynamics, topography and burn severity: a case study from the Montane Cordillera Ecozones of Western Canada. Appl Geogr 56:232–248. https://doi.org/10.1016/j.apgeog.2014.11.016
Jensen AM, Lof M (2017) Effects of interspecific competition from surrounding vegetation on mortality, growth and stem development in young oaks (Quercus robur). For Ecol Manag 392:176–183. https://doi.org/10.1016/j.foreco.2017.03.009
Jiao W, Wang L, Smith WK, Chang Q, Wang H, D’Odorico P (2021) Observed increasing water constraint on vegetation growth over the last three decades. Nat Commun 12(1):1–9. https://doi.org/10.1038/s41467-021-24016-9
Kilgore BM, Dan T (1979) Fire history of a sequoia-mixed conifer forest. Ecology 60(1):129–142. https://doi.org/10.2307/1936475
Kilpeläinen J, Domisch T, Lehto T, Kivimäenpää M, Martz F, Piirainen S, Repo T (2022) Separating the effects of air and soil temperature on silver birch. Part II. The relation of physiology and leaf anatomy to growth dynamics. Tree Physiol. https://doi.org/10.1093/treephys/tpac093
Kong FH (2003) Understory vegetation dynamics of North American boreal forests. Dissertation, Shandong Normal University, Jinan, China
Kong FH, Li XZ, Wang XG, Zhao SL, Shi BL, Gao ZL (2003) Advances in forest restoration in forest fire areas. Chin J Ecol 02:60–64
Kong JJ, Yang J, Cai WH (2018) Topography controls post-fire changes in soil properties in a Chinese boreal forest. Sci Total Environ 651:2662–2670. https://doi.org/10.1016/j.scitotenv.2018.10.164
Lacouture DL, Broadbent EN, Crandall RM (2020) Detecting vegetation recovery after fire in a fire-frequented habitat using normalized difference vegetation index (NDVI). Forests. https://doi.org/10.3390/f11070749
Li XZ (2004) Effects of forest fire factors on forest vegetation succession in the Greater Khingan Mountains. J Fujian Coll For
Li YY, Shao MA (2004) Changes of plant diversity during natural vegetation restoration in Ziwuling Mountains. China Acta Ecologica Sinica 02:252–260
Li XY, Jin HJ, Wang HW, Marchenko SS, Shan W, Luo DL, He RX, Spektor V, Huang YD, Li XY (2021) Influences of forest fires on the permafrost environment: a review. Adv Clim Chang Res 12(1):48–65. https://doi.org/10.1016/j.accre.2021.01.001
Liu B (2011) Dynamic changes of forest fires in the Greater Khingan Mountains. Dissertation, Chinese Academy of Forestry
Liu ZH (2016) Effects of climate and fire on short-term vegetation recovery in the boreal larch forests of Northeastern China. Sci Rep. https://doi.org/10.1038/srep37572
Liu EH, Zan DP, Liu XG, Wu GC, Ma HY, Zhang ZF, Luan CS, Liu ZY (1995) Studies on flowering and Fruiting regularity of Pinus sylvestris (i)—relation between cone variation and seed prediction method. For Sci Technol (02):19–21+16
Liu B, Yang J, Johnstone JF (2017) Understory vascular plant community assembly in relation to time-since-fire and environmental variables in a Chinese boreal forest. J Mt Sci 14(7):1317–1328. https://doi.org/10.1007/s11629-016-4158-1
Liu B, Biswas SR, Yang J, Liu ZH, He HS, Liang Y, Lau MK, Fang YT, Han SJ (2020) Strong influences of stand age and topography on post-fire understory recovery in a Chinese boreal forest. For Ecol Manag. https://doi.org/10.1016/j.foreco.2020.118307
Luo JC (2002) Impact of forest fire on forest ecosystem in the Greater Khingan Mountains. J Beijing for Univ Z1:105–111
Ma J, Xiao X, Bu R, Doughty R, Hu Y, Chen B, Li X, Zhao B (2017) Application of the space-for-time substitution method in validating long-term biomass predictions of a forest landscape model. Environ Model Softw 94:127–139. https://doi.org/10.1016/j.envsoft.2017.04.004
Mao ZH, Zhu JJ (2006) Effects of disturbance on species composition and diversity of plant communities. Acta Ecol Sin 08:2695–2701
Marler TE, del Moral R (2018) Increasing topographic influence on vegetation structure during primary succession. Plant Ecol 219(8):1009–1020. https://doi.org/10.1007/s11258-018-0853-z
Mathews LEH, Kinoshita AM (2021) Urban fire severity and vegetation dynamics in Southern California. Remote Sens. https://doi.org/10.3390/rs13010019
Meng R, Dennison PE, Huang C, Moritz MA, D’Antonio C (2015) Effects of fire severity and post-fire climate on short-term vegetation recovery of mixed-conifer and red fir forests in the Sierra Nevada Mountains of California. Remote Sens Environ. https://doi.org/10.1016/j.rse.2015.10.024
Moradi G, Vacik H (2018) Relationship between vegetation types, soil and topography in southern forests of Iran. J for Res 29(6):1635–1644. https://doi.org/10.1007/s11676-018-0601-1
Moradizadeh H, Heydari M, Omidipour R, Mezbani A, Prevosto B (2020) Ecological effects of fire severity and time since fire on the diversity partitioning, composition and niche apportionment models of post-fire understory vegetation in semi-arid oak forests of Western Iran. Ecol Eng. https://doi.org/10.1016/j.ecoleng.2019.105694
Moya D, Sagra J, Esteban Lucas-Borja M, Antonio Plaza-Alvarez P, Gonzalez-Romero J, De Las Heras J, Ferrandis P (2020) Post-fire recovery of vegetation and diversity patterns in semiarid Pinus halepensis. Mill habitats after salvage logging. Forests. https://doi.org/10.3390/f11121345
Näsholm T, Högberg P, Franklin O, Metcalfe D, Keel SG, Campbell C, Hurry V, Linder S, Högberg MN (2013) Are ectomycorrhizal fungi alleviating or aggravating nitrogen limitation of tree growth in boreal forests? New Phytol. https://doi.org/10.1111/nph.12139
Oettel J, Lapin K (2021) Linking forest management and biodiversity indicators to strengthen sustainable forest management in Europe. Ecol Indic. https://doi.org/10.1016/j.ecolind.2020.107275
Qu L, Wang Y, Masyagina O, Kitaoka S, Fujita S, Kita K, Koike T (2022) Larch: a promising deciduous conifer as an eco-environmental resource. IntechOpen. https://doi.org/10.5772/intechopen.101887
Roberts MR (2004) Response of the herbaceous layer to natural disturbance in North American forests. Can J Bot Revue Canadienne De Botanique 82(9):1273–1283. https://doi.org/10.1139/b04-091
Shan JP (1996) Changes of long and short branches of Larix principis-Larix after fire and their effects on survival. Chin J Appl Ecol S1:6–10
Shi ZG, Li CF, Zhao SY (2014) The growth dynamics of natural birch forest in southern part of the Daxing'anling mountains. For Resourc Manag (01): 62–65+97
Shive KL, Sieg CH, Fulé PZ (2013) Pre-wildfire management treatments interact with fire severity to have lasting effects on post-wildfire vegetation response. For Ecol Manag 297:75–83. https://doi.org/10.1016/j.foreco.2013.02.021
Shu LF, Tian XR, Ma LT (1999) Study and application of forest fire ecology. Res for Sci 04:422–427
Shvetsov EG, Kukavskaya EA, Buryak LV, Barrett K (2019) Assessment of post-fire vegetation recovery in Southern Siberia using remote sensing observations. Environ Res Lett. https://doi.org/10.1088/1748-9326/ab083d
Smith-Ramirez C, Castillo-Mandujano J, Becerra P, Sandoval N, Allende R, Fuentes R (2021) Recovery of Chilean Mediterranean vegetation after different frequencies of fires. For Ecol Manag. https://doi.org/10.1016/j.foreco.2021.118922
Smith-Ramirez C, Castillo-Mandujano J, Becerra P, Sandoval N, Fuentes R, Allende R, Acuna MP (2022) Combining remote sensing and field data to assess recovery of the Chilean Mediterranean vegetation after fire: effect of time elapsed and burn severity. For Ecol Manag. https://doi.org/10.1016/j.foreco.2021.119800
Song QL, Dong XB, Li Y, Qin SL (2010) Evaluation on Burned Slash vegetation natural restoration effects in Daxinganling. For Eng 26(04):14–18. https://doi.org/10.16270/j.cnki.slgc.2010.04.012
Stefanski A (2022) Even modest climate change may lead to major transitions in boreal forests. Nature 608(7923):540–545. https://doi.org/10.1038/s41586-022-05076-3
Stinca A, Ravo M, Marzaioli R, Marchese G, Cordella A, Rutigliano FA, Esposito A (2020) Changes in multi-level biodiversity and soil features in a burned beech forest in the Southern Italian Coastal Mountain. Forests. https://doi.org/10.3390/f11090983
Sun MX (2011) Effects of forest fire on soil properties and vegetation restoration in Tahe Forest region. Dissertation, Beijing Forestry University, Beijing, China
Sun JB, Hu HQ (2010) Community succession in a burnt larch forest of the Greater Khingan Mountains. J Northeast for Univ 38(05):30–33
Thuan C, Guo X, Takeda K (2017) Effects of Burn severity and environmental conditions on post-fire regeneration in Siberian Larch Forest. Forests. https://doi.org/10.3390/f8030076
Ubeda X, Sarricolea P (2016) Wildfires in Chile: a review. Glob Planet Chang 146:152–161. https://doi.org/10.1016/j.gloplacha.2016.10.004
Vasilakos C, Tsekouras GE, Palaiologou P, Kalabokidis K (2018) Neural-network time-series analysis of MODIS EVI for post-fire vegetation regrowth. ISPRS Int J Geo-Inf. https://doi.org/10.3390/ijgi7110420
Vassilis A, Vassilia K, Victoria T, Polychronis K, Milto M, Chistos D (2022) Time series analysis of landsat data for investigating the relationship between land surface temperature and forest changes in Paphos Forest, Cyprus. Remote Sens. https://doi.org/10.3390/rs14041010
Veihmeyer FJ, Hendrickson AH (1950) Soil moisture in relation to plant growth. Annu Rev Plant Physiol 1(1):285–304
Wang D (2016a) Study on characteristics of plant communities at Burned Area of Daxing'an Mountains. Dissertation, Inner Mongolia Agricultural University, Hohhot, China
Wang LH (2016b) Ecological effects of Larix Gmelinii Plantation in Burned Areas of Great Xing'an Mountains. Dissertation, Northeast Forestry University, Harbin, China
Wang XG, Li XZ, He HS, Leng WF, Wen QC (2004) Postfire succession of larch forest in the northern slope of Daxinganling. Chin J Ecol 5:35–41
Wang XL, Wang WJ, Chang Y, Feng YT, Chen HW, Hu YM, Chi JG (2013) Analysis of forest fire intensity based on NBR index in Huzhong forest fire area of the Greater Khingan Mountains. Chin J Appl Ecol 24(04):967–974. https://doi.org/10.13287/j.1001-9332.2013.0250
Wang LH, Xin Y, Zou ML, Zhao YS, Liu SJ, Ren QS (2015) Plant diversity and biomass allocation patterns during vegetation restoration in burned land in the Greater Khingan Mountains. J Beijing for Univ 37:41–47
Wang B, Zhang JY, Meng M, Zhang QL (2021) Study on vegetation restoration characteristics of Daxing’anling Burned Area based on EVI. For Res 34(02):32–41. https://doi.org/10.13275/j.cnki.lykxyj.2021.02.004
Weber RN, Powers MD, Kennedy RE (2022) Vegetation recovery rates provide insight into reburn severity in southwestern Oregon, USA. For Ecol Manag. https://doi.org/10.1016/j.foreco.2022.120292
Wei YM, Hu HQ, Sun JB, Yuan Q, Sun L, Liu HF (2016) Effect of fire intensity on active organic and total soil carbon in a Larix gmelinii forest in the Daxing’anling Mountains, Northeastern China. J for Res 27(6):1351–1359. https://doi.org/10.1007/s11676-016-0251-0
Weinhold A, Döll S, Liu M, Schedl A, Pöschl Y, Xu X, Van Dam NM (2021) Tree species richness differentially affects the chemical composition of leaves, roots and root exudates in four subtropical tree species. J Ecol 110(1):97–116. https://doi.org/10.1111/1365-2745.13777
Wen LX, Guo M, Huang SB, Yu FB, Zhong C, Zhou FF (2021a) The response of vegetation to the change of active layer thickness in permafrost region of the north Greater Khingan Mountains. J Glaciol Geocryol 43(05):1531–1541. https://doi.org/10.7522/j.issn.1000-0240.2021.0103
Wen LX, Guo M, Yin S, Huang SB, Li XL, Yu FB (2021b) Vegetation phenology in permafrost regions of Northeastern China based on MODIS and solar-induced chlorophyll fluorescence. Chin Geogr Sci 31(3):459–473. https://doi.org/10.1007/s11769-021-1204-x
Whitman E, Parisien MA, Thompson DK, Hall RJ, Skakun RS, Flannigan MD (2018a) Variability and drivers of burn severity in the northwestern Canadian boreal forest. Ecosphere. https://doi.org/10.1002/ecs2.2128
Whitman E, Parisien MA, Thompson DK, Flannigan MD (2018b) Topoedaphic and forest controls on post-fire vegetation assemblies are modified by fire history and burn severity in the Northwestern Canadian Boreal Forest. Forests. https://doi.org/10.3390/f9030151
Wu ZY (1982) Flora of China. Science Press, Beijing
Xing W, Ge ZW, Li JQ (2006) The effect of fire severity on the Larix gmelini community in North Great Xing’an Mountains. Sci Technol Eng 14:2042–2046
Yang Y (2019) Effects of fire on forest carbon storage, plant composition diversity and community structure in the Greater Khingan Mountains. Dissertation, Northeast Forestry University, Harbin, China
Yang J, Kong JJ, Liu B (2013) A review of effects of fire disturbance on understory vegetation in boreal coniferous forest. Chin J Plant Ecol 37(05):474–480. https://doi.org/10.3724/SP.J.1258.2013.00049
Yang J, Pan SF, Dangal S, Zhang B, Wang SY, Tian HQ (2017) Continental-scale quantification of post-fire vegetation greenness recovery in temperate and boreal North America. Remote Sens Environ 199:277–290. https://doi.org/10.1016/j.rse.2017.07.022
Yuan H (2020) Temporal and spatial patterns and driving forces of fires in the Greater Khingan Mountains. Dissertation, Shenyang University, Shenyang, China
Yue C, Luo CF, Shu LF, Shen ZH (2020) A review on wildfire studies in the context of global change. Acta Ecol Sin 40(2):1–17. https://doi.org/10.5846/stxb201812202762
Zhang JT (2004) Quantitative ecology. China Science and Technology Press
Zhang L, Yuan XY, Zhang DL (2007) A comparative study of species diversity in the Greater and Lesser Hinggan Mountains flora and crisscross zones. Plant Res 356−360
Zhang SL, Xiao ZL, Huo JP, Zhang HJ (2021a) Key factors influencing on vegetation restoration in the gullies of the Mollisols. J Environ Manag. https://doi.org/10.1016/j.jenvman.2021.113704
Zhang W, Li Y, Wu XC, Chen YH, Chen AP, Schwalm CR, Kimball JS (2021b) Divergent Response of vegetation growth to soil water availability in dry and wet periods over central Asia. J Geophys Res Biogeosci. https://doi.org/10.1029/2020JG005912
Zhao J (2009) Study on process of burned areas forest natural restoration in Daxinganling. Dissertation, Northeast Forestry University, Harbin, China
Zhao FJ, Wang LZ, Chen PY, Shu LF (2013) Review on the recovery after the catastrophic forest fire in Daxing’anling Mountains. For Resour Manag (02):125–129+140
Zheng HN, Jia SQ, Hu HQ (1986) Forest fire and forest restoration in the Greater Khingan Mountains. J Northeast for Univ 04:1–7
Zhou YL (1988) Characteristics, regeneration law and rational management of the Larch forest in northeast China Plant Research (01):127–146
Zhu P, Zhang G, Wang H, Zhang B, Liu Y (2021) Soil moisture variations in response to precipitation properties and plant communities on steep gully slope on the Loess Plateau. Agric Water Manag (256):107086. https://doi.org/10.1016/j.agwat.2021.107086
Zong XZ, Tian XR (2022) The process of vegetation recovery and burn probability changes in post-burn boreal forests in northeast China. Int J Wildland Fire. https://doi.org/10.1071/WF22033
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding: This study was funded by the National Natural Science Foundation of China (No. 41871103).
The online version is available at http://springerlink.fh-diploma.de.
Corresponding editor: Yu Lei.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhong, C., Guo, M., Zhou, F. et al. Forest succession trajectories after fires in valleys and on slopes in the Greater Khingan Mountains, China. J. For. Res. 34, 623–640 (2023). https://doi.org/10.1007/s11676-023-01602-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-023-01602-9