
Abstract
The introduction of alien species can have a significant impact on the food preferences of native phytophagous insects. The moth Earias clorana L. has previously been considered to be monophagous, ingesting only plants in the genus Salix. In recent years, we have observed larval E. clorana feeding on Spiraea tomentosa L., an invasive shrub species in Central Europe that is native to North America. We hypothesised that this insect can feed on Spiraea tomentosa leaves with no negative effects on its growth and development, and that the leaves of Spiraea tomentosa as a source of food for E. clorana are equally as good as leaves of Salix viminalis L. Our results showed that despite significant differences in the chemical composition of the studied species’ leaves, including a much higher concentration of defence compounds (total soluble phenols and condensed tannins) in Spiraea tomentosa leaves than in those of Salix viminalis, feeding on a new host plant did not significantly affect the survival of larvae. The change in host plant had an unfavourable effect, however, on several parameters of growth and development for the larvae (masses of larvae and pupae, relative growth rates, and efficiency of conversion of ingested food). We conclude that, in comparison to Salix viminalis, Spiraea tomentosa is not a particularly favourable food for larval development. Perhaps, even without direct improvements in adult foraging efficiency, however, the costs of switching hosts may be minimised in larvae that develop on very abundant, invasive species, such as Spiraea tomentosa in Central Europe.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
An insect’s choice of host plant depends on the insect’s food preferences and its ability to change the host, which depends on the structure and chemistry of the new host plant, as well as the availability and abundance of the host species in the insect’s habitat (Cunningham et al. 2001; Harvey et al. 2010). As insects are strongly dependent on their preferred food (Ehrlich and Raven 1964; Coley et al. 2006), feeding on the optimal host plant allows them to obtain a larger body size (Haukioja and Neuvonen 1985; Ohmart et al. 1985; Awmack and Leather 2002). Choosing a suboptimal host plant may result in a decrease in adult insect mass (Mąderek et al. 2015) and may extend larval development time, thereby increasing larval mortality due to prolonged exposure to predators and parasitoids (Häggström and Larsson 1995; Williams 1999; Chen and Chen 2016). Decrease in the number of natural enemies is usually observed in the case of plant species relocated beyond their natural range (Keane and Crawley 2002), but native insect species are able to utilise neophyte plant species (e.g. Karolewski et al. 2014). Based on the literature, the impact of alien plants on insects and their host selection preferences remains equivocal (Sunny et al. 2015). Many studies have shown that insects feeding on alien species, especially those that feed on congeneric alien plant species (e.g. Łukowski et al. 2017), develop as well as or better than they would if feeding on native host plants (e.g. Karowe 1990; Hull-Sanders et al. 2007; Chupp and Battaglia 2014). Other researchers have shown a negative impact of neophytes on insects (Tallamy 2004; Casagrande and Dacey 2007; Keeler and Chew 2008; Tallamy et al. 2010; Fortuna et al. 2013), which is confirmed by meta-analyses showing that non-invaded habitats, in comparison to habitats with a high proportion of alien species, had higher total abundance and taxonomic richness of arthropods (Bezemer et al. 2014; van Hengstum et al. 2014; Litt et al. 2014). In situations where the optimal host is a rare species or there is high competition for food, the cost of feeding on the optimal host may be large enough for an insect that it favours the strategy of increasing the number of potential host plants by utilising a new species (Mayhew 1997). The preference–performance hypothesis (aka “mother knows best” hypothesis; Wennström et al. 2010; Kohandani et al. 2016) proposes that oviposition choice (host choice) should correspond with optimal conditions to hatch, survive, grow, and eventually reproduce offspring. Thus, host plant choice should be intended to maximise the performance of offspring. Conversely, another hypothesis, the “bad motherhood” hypothesis, suggests alternate scenarios in which a female could sometimes make “bad” (not the best) oviposition choices (Mayhew 2001), resulting in detrimental conditions for the growth and development of progeny (Digweed 2006; Gripenberg et al. 2007). As the search for an alternative food source is a gradual process and is reversible upon encountering other host species, insects are most likely to utilise the most abundant host species (Cunningham et al. 2001). This appears to favour the colonisation of invasive species, characterised by mass occurrence (Karolewski et al. 2017).
In Poland and Germany, the steeplebush Spiraea tomentosa L. (Rosaceae) is an invasive alien species (Kott 2009; Tokarska-Guzik et al. 2012). Spiraea tomentosa naturally occurs in North America, where its wide strip of range extends along the east coast from Québec in Canada, to Louisiana and Georgia in the southern United States (Gille 1950; USDA—The Plants Database 2017). In its area of natural occurrence, this shrub grows in wetland habitats, where it enters transitional and raised bogs (Reschke 1990; Schafale and Weakley 1990; Faber-Langendoen 2001), and also covers forest clearances and pastures (Gille 1950), as well as hollows in the land (Fike 1999), roadsides, and ditches (Darbyshire 2003). In Europe, this species has been cultivated as an ornamental plant since the eighteenth century (Symes 1983). Currently, it is recognised as a fully acclimated neophyte in six Central European countries (Dajdok et al. 2011).
In Poland, the expansion of Spiraea tomentosa has been the cause of significant changes in the natural environment and impediments to forest management (Wiatrowska and Danielewicz 2016). The shrub often develops patches several dozen hectares in size in peat bogs, moor grass woods (Molinio-Pinetum), and alder-elm-ash riparian forests (Alno-Ulmion), as well as covering ditches, ponds, roadsides, unused meadows, and fallow land (Dajdok et al. 2011; Wiatrowska and Danielewicz 2016; Fig. 1a). The invasive potential of this species results from its biological properties, such as its ability to produce large numbers of viable seeds, its rapid growth, and its early generative maturity (Wiatrowska 2016). This species is not grazed by herbivores in Europe (Burkart 2003) and no literature has yet suggested that insectivorous insects feed or lay eggs on plants of this species (Kott 2009). In recent years, however, we have observed mass feeding by the cream-bordered green pea Earias clorana L. (Fig. 1b) (Lepidoptera: Nolidae) on the shoots and leaves of Spiraea tomentosa, in the Lower Silesian Wilderness (Poland; 51°39′48″N, 15°15′70″E).

a Spiraea tomentosa in Bory Dolnośląskie Forest (western Poland); b Earias clorana leaf tiers on S. tomentosa; c E. clorana leaf tiers on Salix viminalis; and d E. clorana larva
Earias clorana is a small moth with a broad, mainly Eurasian, range extending from the Iberian Peninsula to southwestern Siberia (Buszko and Masłowski 2012). This moth occurs in riparian forests, where its larvae graze mainly on narrowleaf willows Salix sp., with particular preference for Salix viminalis L. (Salicaceae) (Czerniakowski 2005; Nijak 2011). There are also known cases of larval feeding on other species of the genus Salix, including S. triandra L. (West 1985), S. dasyclados Wimm. (Noreĭka and Smalukas 1996), S. fragilis L., and S. purpurea L. (Buszko and Masłowski 2012).
On shoots of Salix viminalis, the larvae wrap the top leaves in silk and feed inside the leaf tiers (Bochniarz 2014) (Fig. 1c, d). The larvae add new leaves to the leaf tiers during the feeding period (Buszko and Masłowski 2012), eating young leaves, bark, and bud tips (Bochniarz 2014). Larval feeding on Spiraea tomentosa shoots is similar to their feeding on shoots of Salix species. Larvae graze mainly on Spiraea tomentosa shoots with inflorescences. In addition to the leaves and bark, flower buds and panicle axes are also grazed, which can damage the inflorescence. On Spiraea tomentosa (pers. observ.), similarly to on Salix species (West 1985), one leaf tier normally feeds one larva, although sometimes two, three, or more larvae can exist together on a tier. It is worth emphasising that approximately 40% of larvae that complete development require more than one leaf tier (West 1985), and thus one larva damages several shoots. Larval feeding significantly reduces the growth and biomass production of Salix viminalis, decreasing it by an average of 30% (West 1985), or even 50% (Czerniakowski 2005); thus, E. clorana is considered one of the most dangerous pests for Salix viminalis (Sulewska et al. 2013).
The aim of this study was to compare survival and performance parameters of larval E. clorana feeding on Salix viminalis (one of their main host plants) and Spiraea tomentosa (a new host on which we have observed larvae). We wanted to determine if egg laying in a localised and invasive alien species is favourable for this moth, or if Spiraea tomentosa will act as a population sink (Casagrande and Dacey 2007) or an evolutionary trap (Schlaepfer et al. 2005) for this native insect. We hypothesised that (1) this insect can survive on Spiraea tomentosa leaves, just as on Salix viminalis, and (2) the leaves of the invasive Spiraea tomentosa are an equally good source of food for E. clorana as are leaves of Salix viminalis, allowing larvae to feed on Spiraea tomentosa with no negative effects on their growth and development. We also hypothesised that (3) there are few, if any, significant differences in leaf structure and chemical composition of food attractants and repellents (e.g. water, carbon nitrogen, sugars, tannins, and phenols) between the two host species. Implementation of this research was essential for understanding the interaction between the native moth E. clorana and its new host Spiraea tomentosa, an invasive alien species.
Materials and methods
Insects and plant materials
We carried out a laboratory experiment on larvae and pupae of the cream-bordered green pea moth E. clorana. Second-generation larvae (L2 instar) were collected from two host plants: Salix viminalis and Spiraea tomentosa. Larvae living on Salix viminalis leaf tiers were sampled from the experimental plot of Poznan University of Life Sciences in Złotniki (Poland; 52°48′57″N, 16°81′31″E), and larvae naturally living on Spiraea tomentosa leaf tiers were collected from a meadow near the Ruszów village in the Lower Silesian Wilderness (Poland; 51°39′48″N, 15°15′70″E). On both plots, plants grew in full light. Larvae were collected in the first half of July 2016, for a total of ca. 250 leaf tiers from each of the host plants.
Seedlings of Spiraea tomentosa (n = 30), obtained from Ruszów, were planted in pots (15 dm3 each) in mid-May 2016 (at the Institute of Dendrology PAS, Kórnik). The pots were filled with native peat (collected with plants in Ruszów) mixed with neutralised peat (1:1). In these studies, we used individuals of Salix viminalis (n = 30) that remained after the completion of other studies carried out at the Institute. Conditions of growth for both species (e.g. soil substrate, water availability) were similar. Seedlings were grown under full light, as in nature, and all plants were watered as necessary.
Experimental design
We carried out a “no-choice” laboratory experiment using a total of 120 larvae of similar masses, in the earliest possible stage of their development. On each of the two host species, 60 larvae were used. Considering the influence of the first food consumed by larvae on their performance in later stages of development (Fortuna et al. 2013), the larvae collected from the main and new host were transferred to the same host plants (from Salix viminalis to Salix viminalis, and from Spiraea tomentosa to Spiraea tomentosa). The larvae were reared individually in Petri dishes (ø 15 cm) with a leaf of the given species, at room temperature with artificial lighting at light levels similar to natural conditions (23 °C; RH 60%; D:N = 16:8). For each larva, their leaf was replaced by a new one and mass was measured daily. The double daily requirement of food for one larva was provided for each larva. The petiole of each leaf was wrapped in a tissue paper soaked in water. Uneaten leaf residues and faeces were weighed after they were dried in a forced air oven at 65 °C for a minimum of 72 h (ULE 600 Memmert GmbH + Co. KG, Germany). To estimate the fresh/dry weight ratio (for each host species and date during feeding period), the fresh weight of a leaf was compared with its dry weight, based on ten samples collected from the leaves of each species.
Daily observations allowed us to compare the mortality of larvae feeding on Salix viminalis and Spiraea tomentosa. To determine insect performance, we used various parameters of growth and development. For the larvae that pupated during the experiment, the duration of larval development (DD) was determined, which we assumed was the time (days) from the beginning of the experiment to pupation. We also determined maximum larval masses and pupal masses (in mg). In addition, we used the following parameters: total food eaten (TFE; mg dry mass), consumption index (CI; g g−1 day−1), relative growth rate (RGR; g g−1 day−1), efficiency of conversion of ingested food (ECI; g g−1 × 100%), approximate digestibility (AD; %) and efficiency with which digested food was converted to body substance (ECD; %).
Total food eaten was calculated by summing the estimated dry mass eaten by the larvae from the start of the experiment to pupation. We determined CI based on the formula CI = TFE/DD × M, where M is the fresh mass of a larva during the feeding period. We assessed RGR using the formula RGR = (Mt − M0)/(DD × M0), where M0 and Mt are the initial and final larval masses, respectively. Based on the larval mass and TFE, we also defined the ECI, using the formula ECI = (final larval mass/TFE) × 100%. We calculated AD using the formula AD = (TFE – F)/(TFE) × 100%, where F is the total mass of faeces. ECD was calculated by dividing the final mass of the larva by TFE – F, and then multiplying the result by 100%. All masses were measured with an analytical balance (Sartorius CP225D; ± 0.01 mg). More details concerning the methods and parameters that we used in this study can be found in the literature (e.g. Waldbauer 1968; Mąderek et al. 2015).
Chemical analysis, SLA, and toughness measurements
We chemically analysed attractants and repellents in the leaves of Salix viminalis and Spiraea tomentosa (n = 10 seedlings/species). We analysed the basic chemical composition of leaves, including water content, carbon and nitrogen concentrations, total non-structural carbohydrates, and defensive compounds such as soluble phenolics and condensed tannins. Plant material was collected in the second half of July from individuals from which leaves had been collected for the laboratory experiments.
Total soluble phenols and condensed tannins were analysed in dried leaf tissue (40 °C for tannins and 65 °C for other phenols), ground in a Mikro-Feinmühle-Culatti mill (IKA Labortechnik Staufen, Germany). The phenolic compound content (TPh) was measured colourimetrically (λ = 660 nm) using Folin and Ciocalteu’s Phenol Reagent (SIGMA F-9252) and following the method of Johnson and Schaal (1957), as modified by Singleton and Rossi (1965). Chlorogenic acid was the standard used in these assays. The results were for total phenols expressed in terms of μM of chlorogenic acid g−1 dry mass (d.m.). Condensed (catechol) tannins, after extraction with absolute methanol, were determined colourimetrically (λ = 500 nm) using a colour reaction with vanillin in an acid medium (Price et al. 1978). These results were converted to μM catechin g−1 d.m. Nitrogen and carbon content (% d.m.) were determined using an Elemental Combustion System CHNS-O 4010 analyser (Costech Instruments, Pioltello, Italy). Total non-structural carbohydrates (TNC; i.e. soluble carbohydrates and starch) were determined as described by Haissig and Dickson (1979) and Hansen and Møller (1975). Soluble carbohydrates were assayed in methanol–chloroform–water extracts (λ = 625 nm) and TNC results were expressed as % d.m. Absorbance (tannins, phenols, and carbohydrates) was determined with a spectrophotometer (UV-1700 Visible Spectrophotometer; PharmaSpec, Shimadzu, Japan). Detailed descriptions of the methods used for these chemical analyses are described in a paper by Karolewski et al. (2013).
Simultaneously, undamaged leaves of both species were also collected for the assessment of specific leaf area (SLA; cm2/g). Each leaf was scanned (software: WinFOLIA 2004, Regent Instruments Inc., Quebec, Canada), dried (65 °C), and weighed. Each sample for SLA analysis was composed of 20 leaves per shrub species.
Toughness of leaves (gf mm−2) was measured with a penetrometer and expressed as punch strength (gf) needed to pass through a leaf lamina with a rod of a given tip area (mm−2). Leaf toughness was measured in mid-July 2016, using a penetrometer with a tip size of 3.5 mm (FHT 801; Equipment Depot, Fotronic, Melrose, MA, USA), in 30 undamaged leaves (for each species) by performing two measurements between the lateral veins, on the left and the right halves of the leaf.
Statistical analyses
Survival analysis (Log-Rank test) was used to determine the probability of survival of larvae over time. Analysis of covariance (ANCOVA) was used to assess the influence of host plant species on maximal larval and pupal masses, DD, TFE, CI, RGR, ECI, AD, and ECD. The initial larval mass was used as a covariate. To assess the significance of the differences in water content, C and N concentrations, TNC, total phenols, condensed tannins, SLA, and toughness of the leaves of the two species, we used one-way analysis of variance (ANOVA).
All statistical analyses were conducted using JMP 12PRO (SAS Institute, Cary, NC, USA) software. For data expressed as percentages, the Bliss arcsin formula was used. Tukey’s HSD test was employed to assess the significance of differences between treatments. Error bars denote the standard error of the mean (± SE).
Results
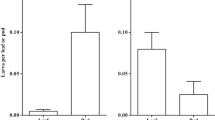
We found that the host plant species that larvae fed on did not affect larval survival (Fig. 2). The insects successfully completed the entire developmental cycle on both Salix viminalis and Spiraea tomentosa, and after the feeding period, the amount of pupated larvae on each of the two host plants was similar.

Survival of larval Earias clorana feeding on Salix viminalis and Spiraea tomentosa leaves in a laboratory experiment
Although the species of host plant did not appear to affect larval survival, some parameters of growth and development indicated that the more favourable food source for E. clorana was the native Salix viminalis (Table 1). Host plant species affected maximal larval and pupal masses. Larvae fed with leaves of the new host plant (Spiraea tomentosa) reached maximal larval and pupal masses that were ca. 7 and ca. 25% lower, respectively, than those of larvae raised on Salix viminalis. We found a statistically significant effect of the species of host plant on both RGR and ECI (Table 1). Larvae that fed on their native host plant reached ca. twofold higher values of RGR and ECI than did those raised on the new host plant. Thus, larvae feeding on Salix viminalis gained body mass at a faster rate and were characterised by better conversion of ingested food.
The species of host plant did not affect TFE or CI, which means that the larvae ingested similar amounts of food, regardless of host plant (Table 1). Host plant also did not affect AD or ECD; however, this is mainly due to the large spread of data. There was also no difference in DD of E. clorana in relation to host plant, estimated based on the larval period from the beginning of the experiment to the pupation (Table 1).
Chemical analysis and leaf toughness
Leaves of Salix viminalis and Spiraea tomentosa significantly differed in relation to water content, SLA, leaf toughness, and chemical compounds (Fig. 3). During the larval feeding period, Spiraea tomentosa leaves contained 36% more water than did Salix viminalis leaves (Fig. 3a). Leaves of Spiraea tomentosa had threefold greater SLA (Fig. 3b), but their toughness was 20% lower than that of Salix viminalis leaves (Fig. 3c). This demonstrates that Salix viminalis leaves are more xeromorphic than are the leaves of Spiraea tomentosa. Spiraea tomentosa leaves had 4% greater carbon content than did Salix viminalis leaves (Fig. 3d), but possessed 46% less nitrogen (Fig. 3e). Spiraea tomentosa leaves also had a significantly higher concentration of TNC than did leaves of Salix viminalis (Fig. 3f). In relation to defence-related compounds, Spiraea tomentosa leaves were characterised by their high concentration, as they contained on average 9.6 times more condensed tannins (Fig. 3g) and 2.8 times more soluble phenols (Fig. 3h) than did Salix viminalis leaves.

Various parameters of Salix viminalis and Spiraea tomentosa leaves characterising their food quality during the larval feeding period: a water; b nitrogen concentration, N; c carbon concentration; d leaf toughness; e specific leaf area, SLA; f total non-structural carbohydrates, TNC; g condensed tannins; and h total soluble phenols, TPh
Discussion
Larval E. clorana can grow on Spiraea tomentosa. Our study showed that the numbers of individuals that pupated were similar, regardless of whether larvae had fed on Salix viminalis or Spiraea tomentosa. Earias clorana larvae, which have until recently been regarded as monophagous, eating only Salix species including mainly Salix viminalis, can survive on Spiraea tomentosa, an alien and invasive shrub species in Central Europe. This result positively validates our first hypothesis, that E. clorana can survive and successfully end its development on a new host plant. Spiraea tomentosa can thus be considered to be a new host plant for E. clorana, although at this time it is difficult to determine whether it is a population sink or an evolutionary trap for this moth species. Grazing on this new host plant negatively affected growth parameters of larvae, however. Larvae fed with Spiraea tomentosa leaves attained 25% lower pupal mass than did larvae raised on Salix viminalis. Feeding on the new host also resulted in the reduction of other growth parameters, such as the relative rate at which larvae gain body mass (Table 1). Although the leaves of Spiraea tomentosa are not the optimal food for E. clorana, increasing host species range by utilising an additional species, however, may be considered beneficial to the population of this moth. Spiraea tomentosa is a widespread species in some areas of Central Europe. Even without direct improvements in adult foraging efficiency, evolution in the host selection behaviour of ovipositing insects can be advantageous because costs caused by switching hosts may be minimised in larvae that develop on a more abundant species (Cunningham et al. 2001). Increased foraging efficiency on a particular, more abundant host species can outweigh the disadvantages of ignoring higher quality, but less abundant, host species.
We found no statistically significant difference in the length of the feeding period of larval E. clorana in relation to host. Since feeding on Spiraea tomentosa does not affect the larval duration of development, there is no significant prolongation of the period that could result in greater risk from natural enemies (Häggström and Larsson 1995; Fortuna et al. 2013). Furthermore, the change of host plant undertaken by E. clorana may restrain intraspecific competition (Feder et al. 1995) and promote escape from natural enemies, such as parasites (Awmack and Leather 2002; Harvey and Fortuna 2012). According to the enemy release hypothesis (Keane and Crawley 2002; Colautti et al. 2004; Stutz et al. 2016), Spiraea tomentosa in its European range of occurrence has not been attacked by insects (Kott 2009). This creates a favourable opportunity for the population of E. clorana to avoid competition with other folivorous species, as well as its natural enemies. For E. clorana, the use of Spiraea tomentosa could therefore result in an enemy-free space (Denno et al. 1990), which is potentially an important factor affecting host plant choice by phytophagous insects (Mulatu et al. 2004). Utilising Spiraea tomentosa may also be beneficial for E. clorana as an opportunity to improve geographical and ecological expansion. In natural habitats, this moth occurs mainly near Salix viminalis trees (Czerniakowski 2010), particularly in lowland valleys on sandy and pebble alluvia within the reach of annual high water levels. Moreover, E. clorana is the most common willow pest on Salix viminalis plantations, used in the biomass energy industry (Tahvanainen and Rytkönen 1999; Sulewska et al. 2013). As it is known, a broader diet allows an herbivore to extend its range (Keeler and Chew 2008), and some herbivores benefit from the establishment and spread of exotic plants because they increase the amount of available resources for them to exploit (Harvey and Fortuna 2012). In the Lower Silesian Wilderness, where we first observed larval feeding on Spiraea tomentosa, Salix viminalis is not a widespread species (Zając and Zając 2001); therefore, the increase in host species range in this area may be conducive to the widening of the range of occurrence for E. clorana, as well as increasing the ecological importance of moth.
The rate at which neophytes are colonised by local herbivorous fauna depends on the degree of phylogenetic similarity of new host plants and their native counterparts, as well as the quality of food, which affects the development of these herbivores (Agrawal and Kotanen 2003; Pearse and Hipp 2009). For example, invasive Prunus serotina Ehrh. demonstrates a strong and close phylogenetic relationship with native P. padus L. (Bortiri et al. 2001), and differences in food quality are small, so native specialists (Karolewski et al. 2014) and generalists (Nowakowska and Halarewicz 2006; Mąderek et al. 2015) have adapted to carrying out the entire life cycle with this new plant. Between Salix viminalis and Spiraea tomentosa, there is no close phylogenetic affinity, and our measurements show that the leaves of these two species differ significantly in relation to both chemical and morphological features. Differences in nitrogen levels and defences in leaves had a significant effect on E. clorana, leading to better performance parameters when larvae were fed Salix viminalis leaves than when fed Spiraea tomentosa leaves. Thus, our second and third hypotheses, that the leaves of the alien Spiraea tomentosa are as good a source of food for E. clorana as are leaves of native Salix viminalis, could not be positively verified.
Salix viminalis leaves are characterised by significantly higher N content and toughness, and lower levels of sugars, phenolic compounds, and tannins, in comparison to leaves of the new host (Fig. 3). It is well known that nitrogen plays a positive role in the development of insects (Giertych et al. 2005; Muth et al. 2008). Increased body mass of insects is considered to be a good indicator of a habitat favourable for survival (Lee and Roh 2010; Kaplan et al. 2014) and of higher quality food (Giertych et al. 2005; Tremmel and Müller 2013). Increasingly, attention is also drawn to the fact that the characteristics determining food quality should include the toughness of leaves, which seriously hampers how much they can be grazed (Scriber and Slansky 1981; Awmack and Leather 2002; Yoshida and Murakami 2012). Taking into account the greater toughness of Salix viminalis leaves, the level of mechanical defence of Spiraea tomentosa leaves is obviously not sufficiently effective to defend against E. clorana because this species can also tolerate it. The leaves of Spiraea tomentosa offer larval E. clorana phagostimulants, like sugars (Harborne 1993; Levesque et al. 2002), the concentrations of which are greater in Spiraea tomentosa leaves than in those of Salix viminalis. Their higher levels usually result in a shorter duration of larval development and pupal period, as well as greater body mass (Hwang and Lindroth 1997; Kaitaniemi et al. 1998; Łukowski et al. 2015). In the case of larvae fed with Spiraea tomentosa leaves, however, we did not find this relationship (Table 1).
Most studies indicate that leaves containing lower concentrations of phenolic compounds and tannins are easier to consume for folivores (Koricheva et al. 1998; Niesenbaum and Kluger 2006; Karolewski et al. 2013). In the case of Spiraea tomentosa, plants were not protected against damage by E. clorana, despite their high concentration of defence compounds (soluble phenols and condensed tannins). According to the general hypothesis that insect specialists, which include E. clorana, are better able to withstand the presence of repellents than are generalists (Mathur et al. 2011; Ali and Agrawal 2012), it is possible that even the high concentration of defence compounds in Spiraea tomentosa leaves did not act as a sufficient barrier to E. clorana. Paradoxically, these compounds may be beneficial for an insect, as larvae sequester plant-derived compounds and the defensive traits of caterpillars are correlated with the defensive traits of their hosts (Bowers 1990; Wheeler et al. 2002). It can be speculated that the high concentrations of phenols and tannins in the leaves of the new host plant increase the defence ability of the moth feeding on them. This issue, however, requires further study.
To summarise our results, it is apparent that there are inherent ecological trade-offs for a herbivore confronted with choosing between its primary host (higher content of nitrogen, lower concentrations of soluble phenols and condensed tannins, but lower content of TNC and water, and higher toughness) and a secondary host (higher TNC and water content, but much higher concentration of defence compounds). We believe, however, that including leaves of Spiraea tomentosa in the diet may be somewhat beneficial for E. clorana. Despite the slightly negative effect of this host on the growth and development of larvae, in the case of the absence or reduction of availability of the main host plant, the widely spread Spiraea tomentosa may allow the moth to survive. Regarding our hypotheses, we conclude that E. clorana can feed on Spiraea tomentosa leaves with no negative effects on its growth, but the new host plant influences larval growth. The leaves of the invasive Spiraea tomentosa are not as good a source of food for E. clorana as are the leaves of Salix viminalis. The reasons for this could include the differences in leaf structure (like toughness), but are likely to be mainly related to the relatively lower N content and higher concentration of repellents (tannins and phenols) in leaves of Spiraea tomentosa.
From the perspective of the host plant, being utilised as a food source may cause a decrease in the number of seeds and reduce total generative potential. Larvae feed on the flower buds and, in extreme cases, this can lead to complete destruction of the inflorescences. Another consequence of increasing the population of E. clorana may also be an increase in damage to Salix viminalis plantations, which already suffer significant damage (Noreĭka and Smaliukas 2005; Nijak 2009; Remlein-Starosta and Nijak 2010). Utilising a new host plant increases the range and abundance of insect populations (Keeler and Chew 2008), which, in the case of determining the location of a new Salix viminalis plantation, would make it necessary to analyse their position relative to localities invaded by Spiraea tomentosa.
The results of the current study, as well as our previous related studies, should also be taken into account when attempting to use E. clorana beyond its natural range, such as for the purpose of reducing populations of invasive species. Recently, Australia proposed a biological control of alien species of willows (including Salix viminalis; Adair et al. 2006), which are major invaders of riparian zones in Australia (Greenwood et al. 2004). The list of insects (mainly specialists restricted to Salix species) that could potentially be used as biological control agents includes E. clorana. Our research shows that E. clorana is not a typical monophagous species, increasing the risk of a possible transfer to another plant species. Furthermore, due to the fact that E. clorana is the first insect species whose larvae have been observed to feed on the invasive Spiraea tomentosa on a massive scale in Europe, in addition to this being the first observation of E. clorana utilising a species belonging to a completely separate phylogenetic family, this issue requires further research. Our current research covers only part of this insect’s life, however. Nobody has analysed the similarity between Salix viminalis and Spiraea tomentosa so far. Therefore, we are planning to determine whether there is a common group of volatile compounds which could serve as insect attractants for both species. We also plan to determine how the new host plant will influence the potential fertility of females and the survival, growth, and development of progeny.
References
Adair R, Sagliocco JL, Bruzzese E (2006) Strategies for the biological control of invasive willows (Salix spp.) in Australia. Aust J Entomol 45:259–267. https://doi.org/10.1111/j.1440-6055.2006.00548.x
Agrawal AA, Kotanen PM (2003) Herbivores and the success of exotic plants: a phylogenetically controlled experiment. Ecol Lett 6(8):712–715
Ali JG, Agrawal AA (2012) Specialist versus generalist insect herbivores and plant defense. Trends Plant Sci 17:293–302
Awmack CS, Leather SR (2002) Host plant quality and fecundity in herbivorous insects. Annu Rev Entomol 47:817–844. https://doi.org/10.1146/annurev.ento.47.091201.145300
Bezemer TM, Harvey JA, Cronin JT (2014) Response of native insect communities to invasive plants. Annu Rev Entomol 59:119–141. https://doi.org/10.1146/annurev-ento-011613-162104
Bochniarz A (2014) Biology and potential harmfulness of cream bordered green pea (Earias clorana L.) Review. Polish J Agron 19:3–10 (in Polish)
Bortiri E, Oh SH, Jiang J, Baggett S, Granger A, Weeks C, Buckingham M, Potter D, Parfitt DE (2001) Phylogeny and systematics of Prunus (Rosaceae) as determined by sequence analysis of ITS and the chloroplast trnL-trnF spacer DNA. Syst Bot 26(4):797–807
Bowers MD (1990) Recycling plant natural products for insect defense. In: Evans DL, Schmidt JO (eds) Insect defenses: adaptive mechanisms and strategies of prey and predators. State University of New York Press, Albany, pp 353–386
Burkart B (2003) Der Einfluss von Schafen, Ziegen und Elchen auf die Vegetation des ehemaligen Panzerschießplatzes Dauban. In: Konold W, Burkart B (eds) Culterra, Schriftenreihe des Institutes für Landespfleg, vol 31. Albert-Ludwigs-Universität Freiburg pp 217–234
Buszko J, Masłowski J (2012) Polish moths. Macrolepidoptera. Koliber & Jarosław Buszko, Janusz Masłowski, Nowy Sącz (in Polish)
Casagrande RA, Dacey JE (2007) Monarch butterfly oviposition on swallow-worts (Vincetoxicum spp.). Environ Entomol 36:631–636
Chen KW, Chen Y (2016) Slow-groth high-mortality: a meta-analysis for insects. Insect Sci. https://doi.org/10.1111/1744-7917.12399
Chupp AD, Battaglia LL (2014) Potential for host shifting in Papilio palamedes following invasion of laurel wilt disease. Biol Invasions 16:2639–2651. https://doi.org/10.1007/s10530-014-0693-2
Colautti RI, Ricciardi A, Grigorovich I, Maclsaac HJ (2004) Is invasion success explained by the enemy release hypothesis? Ecology Lett 7:721–733. https://doi.org/10.1111/j.1461-0248.2004.00616.x
Coley PD, Bateman ML, Kursar TA (2006) The effects of plant quality on caterpillar growth and defense against natural enemies. Oikos 115:219–228. https://doi.org/10.1111/j.2006.0030-1299.14928.x
Cunningham JP, West SA, Zalucki MP (2001) Host selection in phytophagous insects: a new explanation for learning in adults. Oikos 95:537–543. https://doi.org/10.1034/j.1600-0706.2001.950319.x
Czerniakowski ZW (2005) Harmful insects in stool beds willow. Prog Plant Prot 45:77–81 (in Polish)
Czerniakowski ZW (2010) Biodiversity of noxious entomofauna on willow in south-eastern Poland. Fragm Agron 27:19–24 (in Polish)
Dajdok Z, Nowak A, Danielewicz W, Kujawa-Pawlaczyk J, Bena W (2011) NOBANIS—invasive alien species fact sheet - Spiraea tomentosa. https://www.nobanis.org/globalassets/speciesinfo/s/spiraea-tomentosa/spiraea_tomentosa.pdf. Accessed 2 February 2017
Darbyshire SJ (2003) Inventory of Canadian agricultural weeds. Agriculture and Agri-Food Canada, Ottawa
Denno RF, Larsson S, Olmstead KL (1990) Role of enemy-free space and plant quality in host-plant selection by willow. Beetles Ecol 71:124–137. https://doi.org/10.2307/1940253
Digweed SC (2006) Oviposition preference and larval performance in the exotic birch-leafmining sawfly Profenusa thomsonii. Entomol Exp Appl 120:41–49. https://doi.org/10.1111/j.1570-7458.2006.00418.x
Ehrlich PR, Raven PH (1964) Butterflies and plants: a study in coevolution. Evolution 18:586–608. https://doi.org/10.2307/2406212
Faber-Langendoen D (ed) (2001) Plant communities of the Midwest: classification in an ecological context. Association for biodiversity information. Plant Community (Association) Descriptions, Arlington
Feder JL, Reynolds K, Go W, Wang EC (1995) Intra- and interspecific competition and host race formation in the apple maggot fly, Rhagoletis pomonella (Diptera: Tephritidae). Oecologia 101:416–425. https://doi.org/10.1007/BF00329420
Fike J (1999) Terrestrial and palustrine plant communities of Pennsylvania. Pennsylvania Natural Diversity Inventory. Pennsylvania Department of Conservation and Recreation. Bureau of Forestry, Harrisburg
Fortuna TM, Woelke JB, Hordijk CA, Jansen JJ, van Dam NM, Vet LEM, Harvey JA (2013) A tritrophic approach to the preference-performance hypothesis involving an exotic and a native plant. Biol Invasions 15:2387–2401. https://doi.org/10.1007/s10530-013-0459-2
Giertych MJ, Bąkowski M, Karolewski P, Żytkowiak R, Grzebyta J (2005) Influence of mineral fertilization on food quality of oak leaves and utilization efficiency of food components by the gypsy moth. Entomol Exp Appl 117:59–69
Gille A (1950) Le Spiraea tomentosa L. dans la région de Grandby (Comté de Shefford, Québec, Canada): Étude écologique et phytosociologique. Plant Ecol 2:166–196. https://doi.org/10.1007/BF00151675
Greenwood H, O’Dowd DJ, Lake PS (2004) Willow (Salix × rubens) invasion of the riparian zone in south-eastern Australia: reduced abundance and altered composition of terrestrial arthropods. Divers Distrib 10:485–492. https://doi.org/10.1111/j.1366-9516.2004.00104.x
Gripenberg S, Morriën E, Cudmore A, Salminen JP, Roslin T (2007) Resource selection by female moths in a heterogeneous environment: what is a poor girl to do? J Anim Ecol 76:854–865. https://doi.org/10.1111/j.1365-2656.2007.01261.x
Häggström H, Larsson S (1995) Slow larval growth on a suboptimal willow results in high predation mortality in the leaf beetle Galerucella lineola. Oecologia 104:308–315. https://doi.org/10.1007/BF00328366
Haissig BE, Dickson RE (1979) Starch measurement in plant tissue using enzymatic hydrolysis. Physiol Plant 47:151–157. https://doi.org/10.1111/j.1399-3054.1979.tb03207.x
Hansen J, Møller I (1975) Percolation of starch and soluble carbohydrates from plant tissue for quantitative determination with anthrone. Anal Biochem 68:87–94. https://doi.org/10.1016/0003-2697(75)90682-X
Harborne JB (1993) Introduction to ecological biochemistry. Elsevier Academic Press, London
Harvey JA, Fortuna TM (2012) Chemical and structural effects of invasive plants on herbivore-parasitoid/predator interactions in native communities. Entomol Exp Appl 144:14–26. https://doi.org/10.1111/j.1570-7458.2012.01252.x
Harvey JA, Biere A, Fortuna T, Vet LEM, Engelkes T, Morriën E, Gols R, Verhoeven K, Vogel H, Macel M, Heidel-Fischer HM, Schramm K, van der Putten WH (2010) Ecological fits, mis-fits and lotteries involving insect herbivores on the invasive plant, Bunias orientalis. Biol Invasions 12:3045–3059. https://doi.org/10.1007/s10530-010-9696-9
Haukioja E, Neuvonen S (1985) The relationship between size and reproductive potential in male and female Epirrita autumnata (Lep. Geometridae). Ecol Entomol 10:267–270. https://doi.org/10.1111/j.1365-2311.1985.tb00723.x
Hull-Sanders HM, Clare R, Johnson RH, Meyer GA (2007) Evaluation of the evolution of increased competitive ability (EICA) hypothesis: loss of defense against generalist but not specialist herbivores. J Chem Ecol 33:781–799. https://doi.org/10.1007/s10886-007-9252-y
Hwang S-Y, Lindroth RL (1997) Clonal variation in foliar chemistry of aspen: effects on gypsy moths and forest tent caterpillars. Oecologia 111:99–108
Johnson G, Schaal LA (1957) Accumulation of phenolic substances and ascorbic acid in potato tuber tissue upon injury and their possible role in disease resistance. Am Potato J 34:200–209. https://doi.org/10.1007/BF02854948
Kaitaniemi P, Ruohomäki K, Ossipov V, Haukioja E, Pihlaja K (1998) Delayed induced changes in the biochemical composition of host plant leaves during an insect outbreak. Oecologia 116:182–190
Kaplan I, McArt SH, Thaler JS (2014) Plant defenses and predation risk differentially shape patterns of consumption, growth, and digestive efficiency in a guild of leaf-chewing insects. PLoS ONE 9:e93714
Karolewski P, Giertych MJ, Żmuda M, Jagodziński AM, Oleksyn J (2013) Season and light affect constitutive defenses of understory shrub species against folivorous insects. Acta Oecol 53:19–32. https://doi.org/10.1016/j.actao.2013.08.004
Karolewski P, Jagodziński AM, Giertych MJ, Łukowski A, Baraniak E, Oleksyn J (2014) Invasive Prunus serotina—a new host for Yponomeuta evonymellus (Lepidoptera: Yponomeutidae)? Eur J Entomol 111:227–236. https://doi.org/10.14411/eje.2014.026
Karolewski P, Łukowski A, Walczak U, Baraniak E, Mucha J, Giertych MJ (2017) Larval food affects oviposition preference, female fecundity, and offspring survival in Yponomeuta evonymellus. Ecol Entomol 42:657–667. https://doi.org/10.1111/een.12428
Karowe DN (1990) Predicting host range evolution: colonization of Coronilla varia by Colias philodice. Evolution 44:1637–1647. https://doi.org/10.2307/2409343
Keane RM, Crawley MJ (2002) Exotic plant invasions and the enemy release hypothesis. Trends Ecol Evol 17:164–170. https://doi.org/10.1016/S0169-5347(02)02499-0
Keeler MS, Chew FS (2008) Escaping an evolutionary trap: preference and performance of a native insect on an exotic invasive host. Oecologia 156:559–568. https://doi.org/10.1007/s00442-008-1005-2
Kohandani F, Jean G, Goff L, Hance T (2016) Does insect mother know under what conditions it will make their offspring live? Insect Sci 1–9. https://doi.org/10.1111/1744-7917.12300
Koricheva J, Larsson S, Haukioja E, Keinänen M (1998) Regulation plant secondary metabolism by resource availability: hypothesis testing by means of meta-analysis. Oikos 83:212–226
Kott S (2009) Neophytische Spiraea-Arten in der Kernzone “Daubaner Wald” des Biosphärenreservates “Oberlausitzer Heide und Teichlandschaft”. Berichte der Naturforschenden Gesellschaft der Oberlausitz 17:21–36
Lee KP, Roh C (2010) Temperature-by-nutrient interactions affecting growth rate in an insect ectotherm. Entomol Exp Appl 136:151–163
Levesque KR, Fortin M, Mauffette Y (2002) Temperature and food quality effects on growth, consumption and post-ingestive utilization efficiencies of the forest tent caterpillar Malacosoma disstria (Lepidoptera: Lasiocampidae). Bull Entomol Res 92:127–136
Litt AR, Cord EE, Fulbright TE, Schuster GL (2014) Effects of invasive plants on arthropods. Conserv Biol 28:1532–1549. https://doi.org/10.1111/cobi.12350
Łukowski A, Giertych MJ, Zadworny M, Mucha J, Karolewski P (2015) Preferential feeding and occupation of sunlit leaves favors defense response and development in the flea beetle, Altica brevicollis coryletorum—a pest of Corylus avellana. PLoS ONE 10(4):e0126072. https://doi.org/10.1371/journal.pone.0126072
Łukowski A, Giertych MJ, Walczak U, Baraniak E, Karolewski P (2017) Light conditions affect the performance of Yponomeuta evonymellus on its native host Prunus padus and the alien Prunus serotina. Bull Entomol Res 107:208–216. https://doi.org/10.1017/S0007485316000791
Mąderek E, Łukowski A, Giertych MJ, Karolewski P (2015) Influence of native and alien Prunus species and light conditions on performance of the leaf beetle Gonioctena quinquepunctata. Entomol Exp Appl 155:193–205. https://doi.org/10.1111/eea.12298
Mathur V, Ganta S, Raaijmakers CE, Reddy SA, Vet LEM, van Dam NM (2011) Temporal dynamics of herbivore-induced responses in Brassica juncea and their effect on generalist and specialist herbivores. Entomol Exp Appl 139:215–225
Mayhew PJ (1997) Adaptive patterns of host-plant selection by phytophagous insects. Oikos 79:417–428. https://doi.org/10.2307/3546884
Mayhew PJ (2001) Herbivore host choice and optimal bad motherhood. Trends Ecol Evol 16:165–167. https://doi.org/10.1016/S0169-5347(00)02099-1
Mulatu B, Applebaum SW, Coll M (2004) A recently acquired host plant provides an oligophagous insect herbivore with enemy-free space. Oikos 107:231–238. https://doi.org/10.1111/j.0030-1299.2004.13157.x
Muth NZ, Kluger EC, Levy JH, Edwards MJ, Niesenbaum RA (2008) Increased per capita herbivory in the shade: necessity, feedback, or luxury consumption? Ecoscience 15:182–188
Niesenbaum RA, Kluger EC (2006) When studying the effects of light on herbivory, should one consider temperature? The case of Epimecis hortaria F. (Lepidoptera: Geometridae) feeding on Lindera benzoin L. (Lauraceae). Environ Entomol 35:600–606
Nijak K (2009) Pests as causes of destruction in a common willow plantation. Prog Plant Prot 49:1211–1214 (in Polish)
Nijak K (2011) Insect species infesting different clones and cultivars of short rotation willow—a potential risk in crop protection. Prog Plant Prot 51:1563–1566 (in Polish)
Noreĭka R, Smaliukas D (1996) Phytophagous insects as pests of cultivated willow (Salix L.). Ekologija 2:81–87
Noreĭka R, Smaliukas D (2005) Phytophagous insects—pests of industrial plantations of willows Salix L. (Salicaceae) in Lithuania. Ekologija 2:11–14
Nowakowska KM, Halarewicz A (2006) Coleoptera found on neophyte Prunus serotina (Ehrh.) within forest community and open habitat. Electron J Pol Agric Univ. 9(5)
Ohmart CP, Stewart LG, Thomas JR (1985) Effects of food quality, particularly nitrogen concentrations, of Eucalyptus blakelyi foliage on the growth of Paropsis atomaria larvae (Coleoptera: Chrysomelidae). Oecologia 65:543–549. https://doi.org/10.1007/BF00379670
Pearse IS, Hipp AL (2009) Phylogenetic and trait similarity to a native species predict herbivory on non-native oaks. Proc Natl Acad Sci USA 106(43):18097–18102
Price ML, Van Scoyoc S, Butler LG (1978) A critical evaluation of the vanillin reaction as an assay for tannin in sorghum grain. J Agric Food Chem 26:1214–1218. https://doi.org/10.1021/jf60219a031
Remlein-Starosta D, Nijak K (2010) Major diseases and pests of willow grown as energy crop and potential for their control. Prog Plant Prot 50:977–986 (in Polish)
Reschke C (1990) Ecological Communities of New York State. New York Natural Heritage Program. New York State Department of Environmental Conservation, Latham, New York
Schafale MP, Weakley AS (1990) Classification of the Natural Communities of North Carolina. Third approximation. North Carolina Natural Heritage Program. Division of Parks and Recreation, Department of Environment and Natural Resources, Raleigh
Schlaepfer MA, Sherman PW, Blossey B, Runge MC (2005) Introduced species as evolutionary traps. Ecol Lett 8:241–246. https://doi.org/10.1111/j.1461-0248.2005.00730.x
Scriber JM, Slansky F (1981) The nutritional ecology of immature insects. Ann Rev Entomol 26:183–211
Singleton VL, Rossi JA (1965) Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am J Enol Vitic 16:144–158
Stutz S, Štajerová K, Hinz HL, Müller-Schärer H, Schaffner U (2016) Can enemy release explain the invasion success of the diploid Leucanthemum vulgare. Biol Invasions 18(7):2077–2091. https://doi.org/10.1007/s10530-016-1152-z
Sulewska H, Śmiatacz K, Panasiewicz K, Szymańska G, Koziera W (2013) Evaluation of damage and control of cream-bordered green pea (Earias chlorana Hübner) caterpillars in a 4-year old plantation of common willow (Salix viminalis L.). Zemdirbyste-Agric 100:99–104. https://doi.org/10.13080/z-a.2013.100.013
Sunny A, Diwakar S, Sharma GP (2015) Native insects and invasive plants encounters. Arth Plant Int 9:323–331. https://doi.org/10.1007/s11829-015-9384-x
Symes M (1983) Charles Hamilton’s Plantings at Painshill. Garden Hist 11:112–124. https://doi.org/10.2307/1586839
Tahvanainen L, Rytkönen VM (1999) Biomass production of Salix viminalis in southern Finland and the effect of soil properties and climate conditions on its production and survival. Biomass Bioenergy 16:103–117. https://doi.org/10.1016/S0961-9534(98)00074-9
Tallamy DW (2004) Do alien plants reduce insect biomass? Conserv Biol 18:1689–1692. https://doi.org/10.1111/j.1523-1739.2004.00512.x
Tallamy DW, Ballard M, D’Amico V (2010) Can alien plants support generalist insect herbivores? Biol Invasions 12:2285–2292. https://doi.org/10.1007/s10530-009-9639-5
Tokarska-Guzik B, Dajdok Z, Zając M, Zając A, Urbisz A, Danielewicz W, Hołdyński C (2012) Alien plants in Poland with particular reference to invasive species. General Directorate for Environmental Protection, Warsaw (in Polish)
Tremmel M, Müller C (2013) Insect personality depends on environmental conditions. Behav Ecol 24:386–392
USDA—The Plants Database. Species fact sheet - Spiraea tomentosa: https://plants.usda.gov/core/profile?symbol=spto2. Accessed 3 January 2017
van Hengstum T, Hooftman DAP, Oostermeijer JGB, van Tienderen PH (2014) Impact of plant invasions on local arthropod communities: a meta-analysis. J Ecol 102:4–11. https://doi.org/10.1111/1365-2745.12176
Waldbauer GP (1968) The consumption and utilization of food by insects. Adv Insect Phys 5:229–288. https://doi.org/10.1016/S0065-2806(08)60230-1
Wennström A, Hjulström LN, Hjältén J, Julkunen-Tiitto R (2010) Mother really knows best: host choice of adult phytophagous insect females reflects a within-host variation in suitability as larval food. Chemoecology 20:35–42. https://doi.org/10.1007/s00049-010-0040-8
West A (1985) A review of insects affecting production of willows. Information report N-X-232. Canadian Forestry Service, Newfoundland Forestry Centre
Wheeler GS, Massey LM, Southwell IA (2002) Antipredator defense of biological control agent Oxyops vitiosa is mediated by plant volatiles sequestered from the host plant Melaleuca quinquenervia. J Chem Ecol 28:297–315. https://doi.org/10.1023/A:1017982007812
Wiatrowska B (2016) Determinants of the steeplebush (Spiraea tomentosa L.) invasion in the Bory Dolnośląskie Forest. Dissertation, Poznań University of Life Sciences (in Polish)
Wiatrowska B, Danielewicz W (2016) Environmental determinants of the steeplebush (Spiraea tomentosa L.) invasion in the Bory Dolnośląskie Forest. Sylwan 160:696–704 (in Polish)
Williams IS (1999) Slow-growth, high-mortality—a general hypothesis, or is it? Ecol Entomol 24:490–495. https://doi.org/10.1046/j.1365-2311.1999.00217.x
Yoshida K, Murakami M (2012) Larval morphology and feeding behavior in Notodontidae (Lepidoptera) in relation to leaf toughness of host plants. Eur J For Sci 15:45–52
Zając A, Zając M (ed) (2001) Distribution Atlas of vascular plants in Poland. Laboratory of Computer Chorology, Institute of Botany, Jagiellonian University, Cracow (in Polish)
Acknowledgements
This research was financially supported by the statutory activities of the Faculty of Forestry Poznań University of Life Sciences and the Institute of Dendrology Polish Academy of Sciences in Kórnik. We would like to thank M. Bączyk, A. Grzybek, and S. Urbaniak for their technical assistance with the experiments. We are also grateful to the Department of Agronomy Poznań University of Life Sciences for permission to harvest the research material from the experimental surface of Salix viminalis and to the Ruszów Forest District for allowing us to conduct field research at their location.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Livy Williams.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Wiatrowska, B., Łukowski, A., Karolewski, P. et al. Invasive Spiraea tomentosa: a new host for monophagous Earias clorana?. Arthropod-Plant Interactions 12, 423–434 (2018). https://doi.org/10.1007/s11829-017-9592-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11829-017-9592-7