
Abstract
Improving the lung function after experimental allergen challenge by blocking of mast cell (MC) mediators and the capability of MC mediators (including histamine, prostaglandin (PG) D2, and leukotriene (LT) C4) in induction of mucosal edema, bronchoconstriction, and mucus secretion provide evidence that MCs play a key role in pathophysiology of asthma. In asthma, the number of MCs increases in the airways and infiltration of MCs in a variety of anatomical sites including the epithelium, the submucosal glands, and the smooth muscle bundles occurs. MC localization within the ASM is accompanied with the hypertrophy and hyperplasia of the layer, and smooth muscle dysfunction that is mainly observed in forms of bronchial hyperresponsiveness, and variable airflow obstruction. Owing to the expression of a wide range of surface receptors and releasing various cytoplasmic mediators, MCs orchestrate the pathologic events of the disease. MC-released preformed mediators including chymase, tryptase, and histamine and de novo synthesized mediators such as PGD2, LTC4, and LTE4 in addition of cytokines mainly TGFβ1, TSLP, IL-33, IL-4, and IL-13 participate in pathogenesis of asthma. The release of MC mediators and MC/airway cell interactions during remodeling phase of asthma results in persistent cellular and structural changes in the airway wall mainly epithelial cell shedding, goblet cell hyperplasia, hypertrophy of ASM bundles, fibrosis in subepithelial region, abnormal deposition of extracellular matrix (ECM), increased tissue vascularity, and basement membrane thickening. We will review the current knowledge regarding the participation of MCs in each stage of asthma pathophysiology including the releasing mediators and their mechanism of action, expression of receptors by which they respond to stimuli, and finally the pharmaceutical products designed based on the strategy of blocking MC activation and mediator release.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Asthma is characterized by chronic airway inflammation, and hyperresponsiveness (AHR) accompanied by mucus hypersecretion [1]. Triggers of allergic asthma include allergens, fungus (such as Aspergillus fumigatus [2]), viruses (mainly human rhinoviruses (HRV) [3]), and pollutants (including polycyclic aromatic hydrocarbons [4]) [5]. They interact with the airway epithelial cells to initiate the inflammatory response across the airways by releasing of cytokines, particularly IL-25, IL-33, and TSLP [5]. Synergistically with IL-1 and TNF, TSLP stimulates the production of high levels of Th2 cytokines by human MCs [6]. In uncontrolled asthma, MC infiltration to the peripheral airways including the alveolar interstitium occurs. Unlike the healthy subjects, MCs in individuals with asthma express FcεRI and surface bound IgE [7]. The increase in number of MCs in asthmatics is associated with evidence of TH2-skewed inflammation [8] and remodeling with interstitial fibrosis [9]. The participation of MCs in pathogenesis of asthma is supported by the results of tissue biopsies obtained from infants dying of viral bronchiolitis that revealed the presence of large number of tissue resident MCs. It has been reported that these MCs unlike those seen in adult asthmatic individuals did not express surface FcεRI. Development of asthma in children commonly requires both allergic sensitization and viral infection. Considering that sensitization to airborne allergens rarely occurs within the first year of life, there should be a link between MC and viral infection as a predisposing factor for later asthma development [7]. MCs develop from CD34+/CD117+ pluripotent progenitor cells that originate in the bone marrow [10, 11]. The progenitors release into circulation by which access the peripheral tissues via a well-organized integrin/receptor-mediated trafficking. Within the residing tissues, the progenitors differentiate and mature to MCs under the influence of local growth factors, mainly stem cell factor (SCF) [12, 13]. Other MC growth and survival modulators include nerve growth factor (NGF), TGF-β, CXCL12, IL-3, IL-4, IL-9, IL-10, and IL-33 [13]. Human MCs found in connective tissues contain tryptase, chymase, carboxypeptidase, and cathepsin (MCTC), while majority of MCs found in lung and gut express only tryptase (MCT) [14]. Upon IgE-FcεRI-mediated MC activation, subsequent degranulation and release of bioactive mediators occur [15]. MCs produce a wide range of mediators including biogenic amines (histamine and serotonin), serglycin, proteoglycans, proteases (mainly chymase and tryptase), and lipid mediators (platelet-activating factor (PAF), leukotrienes (LTs), and prostaglandins (PGs)) [16]. Additionally, activated MCs release a broad range of pre-stored or de novo synthetized cytokines including GM-CSF, TNFα, IL-3, IL-4, IL-5, IL-6, IL-10, IL-13, and IL-17, chemokines such as CCL2, CCL3, CCL5, and CXCL8, and growth factors including bFGF, NGF, VEGF, TGF-β, and SCF of which the latter acts as the main growth factor for these cells [15]. During immediate allergic reaction (occurring within 10–20 min following allergen exposure), MC released histamine and serotonin cause airway smooth muscle contraction, mucus hypersecretion, and plasma extravasation within the airway wall, that finally result in airway narrowing. The next phase of MC activation occurring within 20–40 min postallergen exposure is determined by the release of the newly produced mediators mainly PGs and LTs that cause further enhancing the allergic airway response [15].
Innate Immune Cells in Asthma (Recruitment and Function)
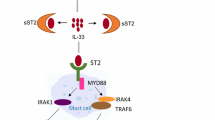
Both innate and specific immune cells actively participate in pathogenesis of asthma. Allergen exposure results in releasing cytokines from airway epithelium mainly IL-33, IL-25, and TSLP which activate ILC-2 cells to proliferate and secrete IL-4 and IL-5 that play a role in induction of IL-13 by Th2 and eosinophil recruitment and activation [17]. ILC2 cells are able to produce IL-13 in an Th2-independent pathway for instance under influence of basophil-derived IL-4 [18, 19]. MC released PGD2 has been reported to activate ILC-2 cells via acting on CRTH2 receptor through which induces the production of type 2 cytokines [20]. T2 cytokines mainly IL-4 and IL-13 cause reduction in junctional complex structure and function of airway epithelial cells in a JAK-dependent manner [21]. Disrupted barrier function possibly promotes allergen sensitization within the airways by accelerating and facilitating the uptake of allergens by subepithelial DCs [21]. Acting via ST2 receptor and MyD88-dependent signaling pathway, IL-33 activates MCs and induces their proliferation [22]. IL-25 acts directly on fibroblasts and endothelial cells to promote airway remodeling and angiogenesis and contributes to production of TSLP and IL-33 in the lung [23]. Eosinophils after being recruited to airways secrete toxic proteins stored in intracellular granules mainly major basic protein (MBP), eosinophil cationic protein(ECP), and reactive oxygen species, which are capable of damaging tissue during allergic inflammation [24, 25]. MBP is known to induce MC degranulation. Moreover, eosinophil released pro-inflammatory mediators mainly LTC4 and LTB4 promote vascular permeability, mucus secretion, and smooth muscle contraction [24]. Several eosinophil surface proteins including CD9, CD11a, CD16, CD25, CD45RO, CD48, CD89, and CD137 have been reported to upregulate in asthma [26]. Eosinophil-released mediators including IL-1, IL-3, IL-5, IL-6, TGF-α, TGF-β, and GM-CSF contribute to airway inflammation. Furthermore, eosinophil-released LTC4, PAF, and 15-HETE (15-hydroxyicosatetraenoic acid) induce acute hyperresponsiveness and ASM hypertrophy [27]. HLMCs activate ILC-2 cells by releasing LTD4 and PGD2 [17]. In return, ILC-2 cells produce IL-9 that is a MC growth factor and promotes IL-4-driven antibody production by B cells and can also induce goblet cell metaplasia [17, 28]. IL-13 induces airway hyperresponsiveness and, in concert with IL-9, promotes mucus production [29]. HLMCs not only release IL-13 but also express the receptor IL-13Rα1. Interestingly, IL-13Rα1 overexpression is reported in asthmatics. IL-13/IL-13Rα1 interaction promotes FcεRI expression on MCs which leads to increased histamine release, MC proliferation, and activation in asthma in an autocrine fashion [30]. Airway epithelial cells contribute in DC recruitment and activation by releasing CCL20 and CCL2 [27]. Moreover, ILC-2-derived IL-13 facilitates the migration of activated lung CD11b+ cDCs to the draining mediastinal lymph nodes (LNs), where they induce Th2 responses [17]. Immature DCs after uptaking the inhaled antigens become mature and migrate to regional LNs to present antigens to naïve T cells which results in differentiation of CD4+ T cells into Th2 cells involved in allergic asthma [31]. Interestingly, some helper T cells capable of producing IL-21 adopt a follicular helper T cell (TFH) subset [17]. TFH cells contribute to producing IL-4 and IL-21, that, along with Th2 cell-derived IL-4, promote class switching in B cells in favor of producing IgE [32]. CD11chi DCs recruit effector Th2 cells to airways through secreting CCL17 and CCL22 [17]. During respiratory syncytial virus (RSV) infection, RSV-infected airway epithelial cells release TSLP which promotes the activation of DCs [33] (Fig. 1).

Upon allergen exposure, air way epithelial cells release IL-25, IL-33, and TSLP and activate ILC-2. DCs after being recruited to lymph nodes act in favor of differentiation of naïve T cells to Th2 cells. TFH cells contribute to Ig class switching in B cells and production of IgE. Antigen-specific IgE molecules sensitize MCs via binding to FcεRI. Further allergen exposures result in MC activation and degranulation. Innate immune cells including eosinophils and DCs are recruited to airways by chemoattractants released by other immune cells
Mast Cell Progenitors from Bone Marrow to Airways: Production and Homing
While SCF, CCL5, CXCL8, CXCL10, CCL11, and CXCL12 are predominant chemokine in MC recruitment to airway epithelium, chemokines including SCF, TGFβ1, CXCL8, CXCL9, CX3CL1, CXCL10, CCL11, and CXCL12 play a role in recruitment of MCs to HASM [34]. Additionally, CXCL9, CXCL10, and CXCL11 act through CXCR3, the most highly expressed HLMC chemokine receptor, and induce a rise in cytosolic-free Ca2+, actin reorganization, and chemotaxis [35, 36]. Human-activated lung MCs release LTB4 that actively attracts the immature MCs via BLT1 receptor to inflammation sites [37]. LTB4-mediated chemoattraction results in MC hyperplasia through which MC progenitors are supplied in the lung tissues [38]. Conversely, PGE2/E-prostanoid (EP)-2 receptor interaction results in inhibiting human lung MC migration [34]. SCF is produced predominantly by epithelial and mesenchymal cells. SCF-CD117 binding induces immature cell proliferation, promotes their chemotaxis to variety of anatomical sites, and suppresses mature MC apoptosis [34]. MCs after being recruited to airways benefit largely from adhesion molecules mainly CADM1 to adhere to human parenchymal lung fibroblasts and HASM cells through homophilic CADM1-CADM1 and CADM1-nectin-3 binding respectively. Human ASM bound SCF mediates MC adhesion via binding to KIT receptor (CD117) [34]. Bronchial smooth muscle (BSM) is infiltrated by MCs upon releasing mediators including TGF-β1, SCF, CXCL10, and CX3CL1 which possess MC chemoattracting activity [39] (Fig. 2, Table 1).

Different chemoattractants are responsible for recruiting MCs to epithelium and ASM. MCs attach to fibroblasts and ASM using homo/heterotypic CADM1, and SCF/CD117 bindings
Role of MC Mediators in Allergic Asthma
Interactions of Mast Cells with Airway Epithelial Cells, Mucous Glands, and Epithelial Goblet Cells
AECs are activated through direct enzymatic activity of the exposed allergens or through activation of a wide range of pattern recognition receptors (PRRs) including TLRs, RIG-I-like receptors (RLRs), NOD-like receptors (NLRs), and C-type lectins. Upon exposure to inhaled allergens, AEC released CCL17 and CCL22 attract and recruit ILC2s, basophils, Tregs, and Th2 through acting on CCR4. Eosinophils and Th2 cells are recruited by AEC released eotaxins CCL11, CCL24, and CCL26 that act via CCR3 receptors. Additionally, AECs are capable of attracting basophils, ILC2s and Th2 cells through releasing PGD2 that binds to CRTH2 receptor [61]. MC-released IL-4 and IL-13 promote the capability of cytokine production by AECs [53]. Both MCs and AECs are able to produce TSLP. AECs owing to expressing TLRs mainly 3, 6, 7, 8, and 9 are able to sense inhaled antigens and in respond to their presence release TSLP. TSLP which is overexpressed in the asthmatic airway promotes the release of Th2 cytokines including IL-4, IL-5, and IL-13 [53]. MCs after localize into the submucosal mucous glands release mediators and cytokines including histamine, PGD2, LTC4, TNFα, chymase, IL-4, IL-6, and IL-13, that consequently promote mucous hypersecretion by hyperplastic submucosal cells and epithelial goblet cells [53]. MC-released IL-13 is the predominant cytokine associated with mucous secretion that promotes the secretion of airway mucus in asthmatics [62]. Excess mucus observed in asthma and COPDs is due to increased biosynthesis of the secretory MUC5AC which is the dominant macromolecule in chemical composition of mucus secreted by airways [63]. IL-13/IL-4Rα interaction activates cytokine receptor-associated Janus kinases (JAKs) which supports the phosphorylation of STAT6. Following dimerization, translocation of phosphorylated STAT6 to the nucleus occurs that suppresses the expression of FOXA2, a transcriptional repressor of MUC5AC [62, 64]. IL-13 signaling promotes transdifferentiation of ciliated to goblet cells [64]. TNFα also is considered as a MC mediator that plays a role in induction of mucous secretion. TNFα/TNFR1 interaction supports receptor trimerization followed by recruitment of multiple signaling proteins to the cytoplasmic domains of TNFR1. TNFR1-associated death domain-containing protein (TRADD) acts as a scaffold for the assembly of downstream signaling complexes, of which receptor-interacting protein1 (RIP1) and TNFR-associated factor2 (TRAF2) are major components. Both RIP1 and TRAF2 are involved in IκB kinase (IKK) activation. IKK promotes the phosphorylation of IκB, after which IκB becomes marked to undergo ubiquitination and degradation by the 26S proteasome. This process is dependent to unmasking a nuclear localization signal and permits subsequent nuclear import of NF-κB [62] (Fig. 3).

Upon allergen exposure, AECs are activated by variety of receptors and release chemoattractant mediators and recruit variety of immune cells to airways. IL-13 and TNF play a crucial role in inducing the mucus production by goblet cells. Both IL-13 and TNF signaling pathways are illustrated
Mast Cell—Airway Smooth Muscle Interactions in Allergic Asthma
The presence of MCs in ASM is considered as a key feature in asthma pathogenesis [65] which is associated to the development of ASM hypertrophy and hyperplasia, ASM dysfunction expressed as BHR, and presence of variable obstruction in airflow [66]. Unlike submucosa residing MCs, the population located in the ASM bundles are always tryptase and chymase positive (MCTC) and their number has been linked to the severity of asthma [59, 67]. Interestingly, MC proteases show different properties when studied in isolation on ASM function. For example, while β-tryptase induces ASM cell proliferation, chymase dramatically reduces it [68]. ASM secretes the three CXCR3 ligands CXCL9, CXCL10, and CXCL11 capable of binding to MC expressed receptor CXCR3 that actively recruit them. SCF/CD117 and TGF-β/TGF-βR also play a role in recruitment of MCs to ASM [66]. Moreover, ASM secreted CXCL10 and CCL5 have a role in the recruitment of MCs via MC expressed CXCR3 and both CCR1 and CCR3, respectively [69]. MC surface expressed CADM-1 facilitates homotypic adhesion between MCs and also heterotypic adhesion with ASM. CADM-1 promotes MC survival and proliferation even in the absence of SCF and is thought to corporate with CD117 in promoting MC survival [57]. CADM-1-mediated MC-ASM adhesion results in ASM activation by MC tryptase, with secretion of TGFβ1 from ASM cells, that upregulates ASM contractile protein expression [57]. PGE2 a lipid mediator of MCs after releasing induces bronchial contractions through interaction with EP1 receptors and relaxations through acting on EP2 or EP4 receptors [70]. Under the function of HLMCs expressed EP2 or EP4 receptors, the concentration of cyclic-AMP raises through activation of adenylyl cyclase. It has been reported that PGE2 works via EP2 receptors to stabilize MCs [71]. Additionally, ASM cells overexpress PAR-2 in asthma. The consequences of PAR-2/MC-derived tryptase include inducing of the calcium mobilization, contraction, and proliferation of human ASM cells [72]. Relatively, overexpression of functional PAR-2 receptors by asthmatic BSM cells accounts for the increased calcium response to PAR-2 stimulation. Repeated PAR-2 stimulations by MC released tryptase promote the proliferation capacity of asthmatic BSM cells [73]. ASM released nerve growth factor (NGF) acts as a MC survival factor that promotes bronchial hyperresponsiveness [74]. Interestingly, ASM release a variety of inflammatory and angiogenic mediators, including eotaxin, GM-CSF, IL-1β, IL-2, IL-5, IL-6, IL-8, IL-10, IL-11, bFGF, PDGF-BB, and VEGF [59]. Recently, it has been reported that ASM-generated pro-MMP-1 after being proteolytically activated by MC-released tryptase plays a role in bronchial hyperresponsiveness. MMP-1 is capable of processing ASM-derived extracellular matrix that enhances ASM proliferation [60]. ASM-derived MMP-1 is induced by collagen I and tenascin C [75] (Fig. 4).

ASM released mediators actively attract MCs to airways. MCs benefit largely from CADM1 and CD117 to attach to ASM. MC Tryptase plays a role in converting pro-MMP-1 to MMP-1 which contributes in bronchial hyperresponsiveness. Tryptase also induces the ASM proliferation, in return ASM-derived TGFβ activates MCs
Mast Cells and Airway Remodeling in Asthma
In asthma, ASM layer infiltration by MCs and secretion of pro-inflammatory and profibrotic mediators are widely accepted to contribute to airway remodeling. Thickening of the asthmatic basement membrane occurs in response to increased deposition of collagen I and III, tenascin, and fibronectin, likely produced by activated myofibroblasts [59]. Subepithelial fibrosis which is another feature of airway remodeling in asthma occurs in the lamina reticularis just below the BM and results in thickening of the BM just below the epithelium [76]. MCs play a predominant role in developing of chronic airway inflammatory changes and remodeling by releasing mediators. Mitogenic properties of MC released tryptase on fibroblasts and to stimulate the synthesis of type 1 collagen in these cells have been reported. Both tryptase and histamine are able to induce ASM proliferation [42]. During airways remodeling TGFβ, another dominant MC mediator involved in airway remodeling promotes epithelial changes and induces subepithelial fibrosis, ASM remodeling, and microvascular changes. TGFβ after being released promotes the differentiation of fibroblasts to myofibroblasts and induces the release of cytokines including fibroblast growth factor-2 (FGF-2) and connective tissue growth factor (CTGF). The latter cytokine enhances the production and deposition of ECM proteins [77]. A number of MC mediators including VEGF, histamine, bFGF, metalloproteinases, IL-8, and proteases are involved in MC-mediated angiogenesis during asthma [40]. MC-derived chymase activates MMP-9 that facilitates the degradation of the ECM and BM, migration of endothelial cells into the interstitial space, and endothelial cell proliferation and differentiation into mature blood vessels [78]. MC-derived proteases, mainly tryptase, and MMPs boost inflammatory responses and airway remodeling in asthma. In turn, ASM-derived mediators, including TGF-β, PGE2, and soluble and membrane-bound SCF modulate the activation state of infiltrating MCs [78].
Mast Cell-Targeted Treatment in Allergic Asthma
There are different therapeutic strategies to target MC-related airway inflammation:
Neutralizing IgE (anti-IgE) and Prevent the Antibody from Linking to the FcεRI
Omalizumab, a clinically approved therapeutic humanized antibody, inhibits the IgE/FcεRI interaction via binding to the Cε3 region on free IgE, prevents MC and basophil activation, and blocks IgE binding to CD23 on B cells and APCs [15, 79, 80]. Moreover, MeDI4212 an antibody with high affinity binds specifically to IgE Cε3 domain and prevents IgE binding to its receptors (FcεRI and CD23) [81].
Neutralizing MC Activator Mediators via Blocking MC Surface Receptors
MCs express a number of receptors that regulate their activation [82]. Tezepelumab (AMG-157) is a fully human neutralizing IgG2 anti-TSLP monoclonal antibody. Tezepelumab inhibits both the early and late allergic responses to a whole long allergen challenge and reduces the number of eosinophils in both blood and sputum of patients with asthma [83, 84]. C-Kit is a surface receptor of SCF expressed on most MCs. It is widely used as a surface marker to identify MCs in tissue. Masitinib is a new kinase inhibitor designed to inhibit c-Kit kinase. It has proven to be effective in mastocytosis and is in a phase III program on patients with severe asthma [85].
Blocking the Signaling Pathways
It is well stablished that cross-linking at FcεRI activates spleen tyrosine kinase (Syk). According to the structure and function, Syk is classified as a ZAP70 family member that is required to mediate MC activation and de novo synthesis of eicosanoids, chemokines, and cytokines [86, 87]. Considering that Syk is located upstream in the cell signaling pathway of multiple immune receptors in human MCs, therapies with Syk inhibitors possibly may be more efficient than drugs that inhibit a single downstream event. In this regard, the Syk inhibitor R343 (Rigel) previously known for its capability to abrogate FcR and BCR signaling has been evaluated by inhaled route in clinical trials for asthma [88].
Blocking the Receptors of Mediators
Blocking the receptors of MC-released mediators could be an effective strategy of controlling the inflammatory effects of MCs in asthma. LTD4 receptor antagonists including “montelukast,” “zafirlukast,” and “pranlukast” are well documented in both asthma and allergic rhinitis [15]. Additionally, the PGD2 receptor “CRTH2” antagonist setipiprant has been reported effective in reducing the late phase reaction after allergen challenge [15]. The pro-inflammatory effects of PGD2 could be observed when it binds to CRTH2 receptor. The receptor is selectively expressed on variety of immune cells including Th2 cells, eosinophils, and basophils [89]. Blocking CRTH2 results in suppressing PGD2 chemotactic activity by which PGD2 recruits effectively circulating eosinophils and basophils to the site of inflammation [89]. Phase 1 clinical trial has revealed the safety selective H4R antagonists including UR-63325, JNJ-39758979, and PF-3893787 in human [27]. Imatinib, a potent inhibitor of c-Kit discoidin domain and platelet-derived growth factor receptors (PDGFR), has been reported to decrease airway hyperresponsiveness, MC counts, and tryptase release in patients with severe asthma [90, 91].
Discussion and Conclusion
It is still not clear why airway remodeling develops in asthma and how such changes contribute to alterations of airway function. In proportion to adult asthma, we also have a poor knowledge regarding childhood asthma. Surprisingly, the frequencies of MCs in the subepithelial mucosa and in the ASM of children who wheeze and with severe asthma as controls show no significant alteration [65]. Moreover, efforts should be done to clarify the exact role of newly discovered MC expressed receptors and released mediators. As an example, mice MC-derived neurotrophin 4 (NT4) has been reported to be in association with persistent changes in ASM innervation and AHR in mice; however, the role of human MC-derived NT4 has been poorly understood in pathogenesis of asthma [92]. Additionally, IFN-γ2 (IL-28A) beyond its role in autoimmunity has been reported to modulate lung DC function to promote T1 immune skewing and suppresses allergic airway after being released from airway MCs in patients with combined rhinitis with asthma [93]. Targeting MC progenitor recruitment may offer an upstream checkpoint to reduce tissue recruited MCs, and the consequences of their presence. The exact molecular mechanism of such recruitment remains unclear most likely it involves integrins and perhaps CADM1, that binds to endothelial CADM1 [48]. Interestingly, the use of IL-37 that binds to IL-18Rα as an anti-inflammatory biological cytokine in suppressing inflammatory cytokines involved in asthma pathogenesis is under investigation. IL-37 capability of suppressing the secretion of pro inflammatory cytokines released from MCs including IL-1, IL-6, IL-8, and TNF-α makes it a promising cytokine to control the MCs [94]. It also increases activated Treg, APC, activated antigen sensitized T cells, and naïve T cells [94]. In humans, the KCa3.1 which is an intermediate conductance Ca2+ activated K+ channel is activated following FcεRI-dependent activation and enhances the influx of Ca2+ (via Orai 1 channels) and histamine release in HLMCs. KCa3.1 blockers such as TRAM-34 have been investigated in mice models of asthma, but still their effectiveness in human asthma has not been proven [57]. Interestingly, attention has been given to immune modulating properties of TLRs especially TLR9 to redirect allergic Th2 responses by triggering Th1 response via TLR activation to control and treat asthma [95]. In recent years, IL-18, a pro-inflammatory cytokine, was introduced as an IFN-γ-inducing factor. IL-18 is emerging to be involved in the pathogenesis of asthma through promoting the production of Th2 cytokines by T cells, NK cells, basophils, and MCs in mice models. Although IL-18 levels have been reported to elevate in patients with asthma, and that human MCs express IL-18R, the exact role of IL-18 with the focus to MCs needs to be investigated in human [96, 97]. Most recently, lysosomotropic agents mainly mefloquine or siramesine that induce the HLMCs apoptosis via permeabilizing the secretory granules of HLMCs and releasing the contents of the granules into the cytosol have been reported to be promising in targeting HLMCs in asthma [98]. Upon acute inflammatory reaction ATP levels increase and adenosine forms through ATP breakdown. ATP and adenosine are capable of activating HLMCs expressed P2Y, P2X, and adenosine receptors [99]. P2X7 selective antagonists possibly could be used for the treatment of MC-mediated chronic inflammatory diseases mainly asthma [99]. One aspect of asthma pathogenesis in human that should be investigated precisely is the role of TLR signaling which has been extensively studied in mice models of asthma. For instance, mediatory role of the BLT2 ligand–BLT2 axis in LPS/TLR4 signaling in producing Th2 cytokines especially IL-13 has been reported [100]. It has been reported that combined stimulation of FcεRI and TLR induces a synergistic cytokine response in MCs, suggesting a contributory role of MCs to allergic exacerbations in the presence of pathogens [101]. Interestingly, in a human in vitro model using LAD2 cells, lipoteichoic acid is reported to reduce the surface expression of FcεRI through TLR2. Yoshioka et al. concluded that TLR2 ligands may be used as a therapy for controlling allergic disorders [102]. Finally, considering the similarities between MCs and basophils including their ability of releasing histamine, leukotrienes, and Th2-related cytokines following IgE-dependent stimulation and the fact that basophils increase in number in tissue and sputum of patients with asthma, investigations should be done to clarify their overlapping roles with MCs in asthma [103]. Investigations to determining the exact role of MCs in pathogenesis of asthma have limitations as any other MC related study; first, unlike animal model, obtaining tissue samples from human airways is limited to postmortem samples or specimen obtained through surgical treatment. Additionally, unlike mice models, there is no transgenic or genetically knockdown human models to participate in MC investigations. MC activation syndrome or mastocytosis may provide opportunity to investigate the role of MCs in pathogenesis of asthma when compared with normal individuals. Although, it seems logical to assume that patients with mastocytosis will tend to have bronchial hyperresponsiveness due to high burden of infiltrated MCs in airway, surprisingly, very few cases have been reported [104].
Abbreviations
- AEC:
-
Airway epithelial cell
- ASM:
-
Airway smooth muscle
- BAL:
-
Bronchoalveolar lavage
- bFGF-2:
-
Basic fibroblast growth factor-2
- BHR:
-
Bronchial hyperresponsiveness
- BM:
-
Basement membrane
- BSM:
-
Bronchial smooth muscle
- EB:
-
Eosinophilic bronchitis
- ECP:
-
Eosinophil cationic protein
- HLMC:
-
Human lung mast cell
- ICAM-1:
-
Intercellular adhesion molecule-1
- LT:
-
Leukotriene
- PAR:
-
Protease-activated receptor
- PDGF:
-
Platelet-derived growth factor
- PG:
-
Prostaglandin
- SCF:
-
Stem cell factor
- VCAM-1:
-
Vascular cell adhesion molecule-1
- CRTH2:
-
Chemoattractant receptor-homologous molecule expressed on TH2 cells
References
Komi DE, Kazemi T, Bussink AP (2016) New insights into the relationship between chitinase-3-like-1 and asthma. Curr Allergy Asthma Rep 16(8):57. https://doi.org/10.1007/s11882-016-0637-2
Singh M, Das S, Chauhan A, Paul N, Sodhi KS, Mathew J, Chakrabarti A (2015) The diagnostic criteria for allergic bronchopulmonary aspergillosis in children with poorly controlled asthma need to be re-evaluated. Acta Paediatr 104(5):e206–e209. https://doi.org/10.1111/apa.12930
Akoto C, Davies DE, Swindle EJ (2017) Mast cells are permissive for rhinovirus replication: potential implications for asthma exacerbations. Clin Exp Allergy 47(3):351–360. https://doi.org/10.1111/cea.12879
Wong TH, Lee CL, Su HH, Lee CL, Wu CC, Wang CC, Sheu CC, Lai RS, Leung SY, Lin CC, Wei YF, Wang CJ, Lin YC, Chen HL, Huang MS, Yen JH, Huang SK, Suen JL (2018) A prominent air pollutant, Indeno[1,2,3-cd]pyrene, enhances allergic lung inflammation via aryl hydrocarbon receptor. Sci Rep 8(1):5198. https://doi.org/10.1038/s41598-018-23542-9
Boita M, Heffler E, Omede P, Bellocchia M, Bussolino C, Solidoro P, Giorgis V, Guerrera F, Riva G, Brussino L, Bucca C, Rolla G (2018) Basophil membrane expression of epithelial cytokine receptors in patients with severe asthma. Int Arch Allergy Immunol 175(3):171–176. https://doi.org/10.1159/000486314
Allakhverdi Z, Comeau MR, Jessup HK, Yoon BR, Brewer A, Chartier S, Paquette N, Ziegler SF, Sarfati M, Delespesse G (2007) Thymic stromal lymphopoietin is released by human epithelial cells in response to microbes, trauma, or inflammation and potently activates mast cells. J Exp Med 204(2):253–258. https://doi.org/10.1084/jem.20062211
Andersson CK, Bergqvist A, Mori M, Mauad T, Bjermer L, Erjefalt JS (2011) Mast cell-associated alveolar inflammation in patients with atopic uncontrolled asthma. J Allergy Clin Immunol 127(4):905–912.e901-907. https://doi.org/10.1016/j.jaci.2011.01.022
Bergqvist A, Andersson CK, Mori M, Walls AF, Bjermer L, Erjefalt JS (2015) Alveolar T-helper type-2 immunity in atopic asthma is associated with poor clinical control. Clin Sci (Lond) 128(1):47–56. https://doi.org/10.1042/cs20140309
Andersson CK, Weitoft M, Rydell-Tormanen K, Bjermer L, Westergren-Thorsson G, Erjefalt JS (2018) Uncontrolled asthmatics have increased FceRI(+) and TGF-beta-positive MCTC mast cells and collagen VI in the alveolar parenchyma. Clin Exp Allergy 48(3):266–277. https://doi.org/10.1111/cea.13092
Elieh-Ali-Komi D, Cao Y (2017) Role of mast cells in the pathogenesis of multiple sclerosis and experimental autoimmune encephalomyelitis. Clin Rev Allergy Immunol 52(3):436–445. https://doi.org/10.1007/s12016-016-8595-y
Elieh Ali Komi D, Rambasek T, Bielory L (2018) Clinical implications of mast cell involvement in allergic conjunctivitis. Allergy 73(3):528–539. https://doi.org/10.1111/all.13334
Elieh Ali Komi D, Grauwet K (2017) Role of mast cells in regulation of T cell responses in experimental and clinical settings. Clin Rev Allergy Immunol 54:432–445. https://doi.org/10.1007/s12016-017-8646-z
Komi DEA, Rambasek T, Wohrl S (2017) Mastocytosis: from a molecular point of view. Clin Rev Allergy Immunol. https://doi.org/10.1007/s12016-017-8619-2
Ali H (2016) Mas-related G protein coupled receptor-X2: a potential new target for modulating mast cell-mediated allergic and inflammatory diseases. J Immunobiol 1(4)
Andersson C, Tufvesson E, Diamant Z, Bjermer L (2016) Revisiting the role of the mast cell in asthma. Curr Opin Pulm Med 22(1):10–17. https://doi.org/10.1097/mcp.0000000000000228
Kubo M (2017) Innate and adaptive type 2 immunity in lung allergic inflammation. Immunol Rev 278(1):162–172. https://doi.org/10.1111/imr.12557
Lambrecht BN, Hammad H (2015) The immunology of asthma. Nat Immunol 16(1):45–56. https://doi.org/10.1038/ni.3049
Zhu J (2015) T helper 2 (Th2) cell differentiation, type 2 innate lymphoid cell (ILC2) development and regulation of interleukin-4 (IL-4) and IL-13 production. Cytokine 75(1):14–24. https://doi.org/10.1016/j.cyto.2015.05.010
Motomura Y, Morita H, Moro K, Nakae S, Artis D, Endo TA, Kuroki Y, Ohara O, Koyasu S, Kubo M (2014) Basophil-derived interleukin-4 controls the function of natural helper cells, a member of ILC2s, in lung inflammation. Immunity 40(5):758–771. https://doi.org/10.1016/j.immuni.2014.04.013
Xue L, Salimi M, Panse I, Mjosberg JM, McKenzie AN, Spits H, Klenerman P, Ogg G (2014) Prostaglandin D2 activates group 2 innate lymphoid cells through chemoattractant receptor-homologous molecule expressed on TH2 cells. J Allergy Clin Immunol 133(4):1184–1194. https://doi.org/10.1016/j.jaci.2013.10.056
Saatian B, Rezaee F, Desando S, Emo J, Chapman T, Knowlden S, Georas SN (2013) Interleukin-4 and interleukin-13 cause barrier dysfunction in human airway epithelial cells. Tissue Barriers 1(2):e24333. https://doi.org/10.4161/tisb.24333
Saluja R, Khan M, Church MK, Maurer M (2015) The role of IL-33 and mast cells in allergy and inflammation. Clin Transl Allergy 5:33. https://doi.org/10.1186/s13601-015-0076-5
Lambrecht BN, Hammad H (2013) Asthma: the importance of dysregulated barrier immunity. Eur J Immunol 43(12):3125–3137. https://doi.org/10.1002/eji.201343730
Lambrecht BN, Persson EK, Hammad H (2017) Myeloid cells in asthma. Microbiol Spectr 5(1). https://doi.org/10.1128/microbiolspec.MCHD-0053-2016
Wilson SJ, Rigden HM, Ward JA, Laviolette M, Jarjour NN, Djukanovic R (2013) The relationship between eosinophilia and airway remodelling in mild asthma. Clin Exp Allergy 43(12):1342–1350. https://doi.org/10.1111/cea.12156
Metcalfe DD, Pawankar R, Ackerman SJ, Akin C, Clayton F, Falcone FH, Gleich GJ, Irani AM, Johansson MW, Klion AD, Leiferman KM, Levi-Schaffer F, Nilsson G, Okayama Y, Prussin C, Schroeder JT, Schwartz LB, Simon HU, Walls AF, Triggiani M (2016) Biomarkers of the involvement of mast cells, basophils and eosinophils in asthma and allergic diseases. World Allergy Organ J 9(7):7. https://doi.org/10.1186/s40413-016-0094-3
Sulaiman I, Lim JC, Soo HL, Stanslas J (2016) Molecularly targeted therapies for asthma: current development, challenges and potential clinical translation. Pulm Pharmacol Ther 40:52–68. https://doi.org/10.1016/j.pupt.2016.07.005
Yu QN, Guo YB, Li X, Li CL, Tan WP, Fan XL, Qin ZL, Chen D, Wen WP, Zheng SG, Fu QL (2018) ILC2 frequency and activity are inhibited by glucocorticoid treatment via STAT pathway in patients with asthma. Allergy 73:1860–1870. https://doi.org/10.1111/all.13438
Elieh Ali Komi D, Sharma L, Dela Cruz CS (2018) Chitin and its effects on inflammatory and immune responses. Clin Rev Allergy Immunol 54(2):213–223. https://doi.org/10.1007/s12016-017-8600-0
Kaur D, Hollins F, Woodman L, Yang W, Monk P, May R, Bradding P, Brightling CE (2006) Mast cells express IL-13R alpha 1: IL-13 promotes human lung mast cell proliferation and Fc epsilon RI expression. Allergy 61(9):1047–1053. https://doi.org/10.1111/j.1398-9995.2006.01139.x
Lee CC, Lin CL, Leu SJ, Lee YL (2018) Overexpression of notch ligand delta-like-1 by dendritic cells enhances their immunoregulatory capacity and exerts antiallergic effects on Th2-mediated allergic asthma in mice. Clin Immunol 187:58–67. https://doi.org/10.1016/j.clim.2017.10.005
Shamji MH, Durham SR (2017) Mechanisms of allergen immunotherapy for inhaled allergens and predictive biomarkers. J Allergy Clin Immunol 140(6):1485–1498. https://doi.org/10.1016/j.jaci.2017.10.010
Garcia-Garcia ML, Calvo C, Ruiz S, Pozo F, Del Pozo V, Remedios L, Exposito N, Tellez A, Casas I (2017) Role of viral coinfections in asthma development. PLoS One 12(12):e0189083. https://doi.org/10.1371/journal.pone.0189083
Bradding P, Arthur G (2016) Mast cells in asthma--state of the art. Clin Expe Allergy 46(2):194–263. https://doi.org/10.1111/cea.12675
Brightling CE, Kaur D, Berger P, Morgan AJ, Wardlaw AJ, Bradding P (2005) Differential expression of CCR3 and CXCR3 by human lung and bone marrow-derived mast cells: implications for tissue mast cell migration. J Leukoc Biol 77(5):759–766. https://doi.org/10.1189/jlb.0904511
Kelsen SG, Aksoy MO, Yang Y, Shahabuddin S, Litvin J, Safadi F, Rogers TJ (2004) The chemokine receptor CXCR3 and its splice variant are expressed in human airway epithelial cells. Am J Physiol Lung Cell Mol Physiol 287(3):L584–L591. https://doi.org/10.1152/ajplung.00453.2003
Weller CL, Collington SJ, Hartnell A, Conroy DM, Kaise T, Barker JE, Wilson MS, Taylor GW, Jose PJ, Williams TJ (2007) Chemotactic action of prostaglandin E2 on mouse mast cells acting via the PGE2 receptor 3. Proc Natl Acad Sci U S A 104(28):11712–11717. https://doi.org/10.1073/pnas.0701700104
Weller CL, Collington SJ, Brown JK, Miller HR, Al-Kashi A, Clark P, Jose PJ, Hartnell A, Williams TJ (2005) Leukotriene B4, an activation product of mast cells, is a chemoattractant for their progenitors. J Exp Med 201(12):1961–1971. https://doi.org/10.1084/jem.20042407
Girodet PO, Ozier A, Trian T, Begueret H, Ousova O, Vernejoux JM, Chanez P, Marthan R, Berger P, Tunon de Lara JM (2010) Mast cell adhesion to bronchial smooth muscle in asthma specifically depends on CD51 and CD44 variant 6. Allergy 65(8):1004–1012. https://doi.org/10.1111/j.1398-9995.2009.02308.x
Zanini A, Chetta A, Saetta M, Baraldo S, D'Ippolito R, Castagnaro A, Neri M, Olivieri D (2007) Chymase-positive mast cells play a role in the vascular component of airway remodeling in asthma. J Allergy Clin Immunol 120(2):329–333. https://doi.org/10.1016/j.jaci.2007.04.021
Lefrancais E, Duval A, Mirey E, Roga S, Espinosa E, Cayrol C, Girard JP (2014) Central domain of IL-33 is cleaved by mast cell proteases for potent activation of group-2 innate lymphoid cells. Proc Natl Acad Sci U S A 111(43):15502–15507. https://doi.org/10.1073/pnas.1410700111
Modena BD, Dazy K, White AA (2016) Emerging concepts: mast cell involvement in allergic diseases. Transl Res 174:98–121. https://doi.org/10.1016/j.trsl.2016.02.011
Gauthier M, Chakraborty K, Oriss TB, Raundhal M, Das S, Chen J, Huff R, Sinha A, Fajt M, Ray P, Wenzel SE, Ray A (2017) Severe asthma in humans and mouse model suggests a CXCL10 signature underlies corticosteroid-resistant Th1 bias. JCI Insight 2(13). https://doi.org/10.1172/jci.insight.94580
Scarpelli MP, Keller S, Tran L, Palmiere C (2016) Postmortem serum levels of IgE and mast cell tryptase in fatal asthma. Forensic Sci Int 269:113–118. https://doi.org/10.1016/j.forsciint.2016.11.001
Lewis RJ, Chachi L, Newby C, Amrani Y, Bradding P (2016) Bidirectional counter regulation of human lung mast cell and airway smooth muscle beta2 adrenoceptors. J Immunol 196(1):55–63. https://doi.org/10.4049/jimmunol.1402232
Parameswaran K, Radford K, Fanat A, Stephen J, Bonnans C, Levy BD, Janssen LJ, Cox PG (2007) Modulation of human airway smooth muscle migration by lipid mediators and Th-2 cytokines. Am J Respir Cell Mol Biol 37(2):240–247. https://doi.org/10.1165/rcmb.2006-0172OC
Singh D, Ravi A, Southworth T (2017) CRTH2 antagonists in asthma: current perspectives. Clin Pharmacol 9:165–173. https://doi.org/10.2147/cpaa.s119295
Arthur G, Bradding P (2016) New developments in mast cell biology: clinical implications. Chest 150(3):680–693. https://doi.org/10.1016/j.chest.2016.06.009
Diamant Z, Mantzouranis E, Bjermer L (2009) Montelukast in the treatment of asthma and beyond. Expert Rev Clin Immunol 5(6):639–658. https://doi.org/10.1586/eci.09.62
Lazarinis N, Bood J, Gomez C, Kolmert J, Lantz AS, Gyllfors P, Davis A, Wheelock CE, Dahlen SE, Dahlen B (2018) Leukotriene E4 induces airflow obstruction and mast cell activation via the CysLT1 receptor. J Allergy Clin Immunol 142:1080–1089. https://doi.org/10.1016/j.jaci.2018.02.024
Kasperska-Zajac A, Brzoza Z, Rogala B (2008) Platelet-activating factor (PAF): a review of its role in asthma and clinical efficacy of PAF antagonists in the disease therapy. Recent Patents Inflamm Allergy Drug Discov 2(1):72–76
KleinJan A (2016) Airway inflammation in asthma: key players beyond the Th2 pathway. Curr Opin Pulm Med 22(1):46–52. https://doi.org/10.1097/mcp.0000000000000224
Carter RJ, Bradding P (2011) The role of mast cells in the structural alterations of the airways as a potential mechanism in the pathogenesis of severe asthma. Curr Pharm Des 17(7):685–698
Agrawal S, Townley RG (2014) Role of periostin, FENO, IL-13, lebrikzumab, other IL-13 antagonist and dual IL-4/IL-13 antagonist in asthma. Expert Opin Biol Ther 14(2):165–181. https://doi.org/10.1517/14712598.2014.859673
Hodsman P, Ashman C, Cahn A, De Boever E, Locantore N, Serone A, Pouliquen I (2013) A phase 1, randomized, placebo-controlled, dose-escalation study of an anti-IL-13 monoclonal antibody in healthy subjects and mild asthmatics. Br J Clin Pharmacol 75(1):118–128. https://doi.org/10.1111/j.1365-2125.2012.04334.x
Reuter S, Stassen M, Taube C (2010) Mast cells in allergic asthma and beyond. Yonsei Med J 51(6):797–807. https://doi.org/10.3349/ymj.2010.51.6.797
Virk H, Arthur G, Bradding P (2016) Mast cells and their activation in lung disease. Transl Res 174:60–76. https://doi.org/10.1016/j.trsl.2016.01.005
Nakae S, Morita H, Ohno T, Arae K, Matsumoto K, Saito H (2013) Role of interleukin-33 in innate-type immune cells in allergy. Allergol Int 62(1):13–20. https://doi.org/10.2332/allergolint.13-RAI-0538
Keglowich LF, Borger P (2015) The three A’s in asthma - airway smooth muscle, airway remodeling & angiogenesis. Open Respir Med J 9:70–80. https://doi.org/10.2174/1874306401509010070
Naveed SU, Clements D, Jackson DJ, Philp C, Billington CK, Soomro I, Reynolds C, Harrison TW, Johnston SL, Shaw DE, Johnson SR (2017) Matrix metalloproteinase-1 activation contributes to airway smooth muscle growth and asthma severity. Am J Respir Crit Care Med 195(8):1000–1009. https://doi.org/10.1164/rccm.201604-0822OC
Thiriou D, Morianos I, Xanthou G, Samitas K (2017) Innate immunity as the orchestrator of allergic airway inflammation and resolution in asthma. Int Immunopharmacol 48:43–54. https://doi.org/10.1016/j.intimp.2017.04.027
Lai H, Rogers DF (2010) New pharmacotherapy for airway mucus hypersecretion in asthma and COPD: targeting intracellular signaling pathways. J Aerosol Med Pulm Drug Deliv 23(4):219–231. https://doi.org/10.1089/jamp.2009.0802
Alevy YG, Patel AC, Romero AG, Patel DA, Tucker J, Roswit WT, Miller CA, Heier RF, Byers DE, Brett TJ, Holtzman MJ (2012) IL-13-induced airway mucus production is attenuated by MAPK13 inhibition. J Clin Invest 122(12):4555–4568. https://doi.org/10.1172/jci64896
Tyner JW, Kim EY, Ide K, Pelletier MR, Roswit WT, Morton JD, Battaile JT, Patel AC, Patterson GA, Castro M, Spoor MS, You Y, Brody SL, Holtzman MJ (2006) Blocking airway mucous cell metaplasia by inhibiting EGFR antiapoptosis and IL-13 transdifferentiation signals. J Clin Invest 116(2):309–321. https://doi.org/10.1172/jci25167
Lezmi G, Galmiche-Rolland L, Rioux S, Jaubert F, Tillie-Leblond I, Scheinmann P, Gosset P, de Blic J (2016) Mast cells are associated with exacerbations and eosinophilia in children with severe asthma. Eur Respir J 48(5):1320–1328. https://doi.org/10.1183/13993003.00947-2016
Bradding P, Walls AF, Holgate ST (2006) The role of the mast cell in the pathophysiology of asthma. J Allergy Clin Immunol 117(6):1277–1284. https://doi.org/10.1016/j.jaci.2006.02.039
Wang G, Baines KJ, Fu JJ, Wood LG, Simpson JL, McDonald VM, Cowan DC, Taylor DR, Cowan JO, Gibson PG (2016) Sputum mast cell subtypes relate to eosinophilia and corticosteroid response in asthma. Eur Respir J 47(4):1123–1133. https://doi.org/10.1183/13993003.01098-2015
Alkhouri H, Hollins F, Moir LM, Brightling CE, Armour CL, Hughes JM (2011) Human lung mast cells modulate the functions of airway smooth muscle cells in asthma. Allergy 66(9):1231–1241. https://doi.org/10.1111/j.1398-9995.2011.02616.x
Manning BM, Meyer AF, Gruba SM, Haynes CL (2015) Single-cell analysis of mast cell degranulation induced by airway smooth muscle-secreted chemokines. Biochim Biophys Acta 1850(9):1862–1868. https://doi.org/10.1016/j.bbagen.2015.05.008
Safholm J, Manson ML, Bood J, Delin I, Orre AC, Bergman P, Al-Ameri M, Dahlen SE, Adner M (2015) Prostaglandin E2 inhibits mast cell-dependent bronchoconstriction in human small airways through the E prostanoid subtype 2 receptor. J Allergy Clin Immunol 136(5):1232–1239.e1231. https://doi.org/10.1016/j.jaci.2015.04.002
Kay LJ, Gilbert M, Pullen N, Skerratt S, Farrington J, Seward EP, Peachell PT (2013) Characterization of the EP receptor subtype that mediates the inhibitory effects of prostaglandin E2 on IgE-dependent secretion from human lung mast cells. Clin Exp Allergy 43(7):741–751. https://doi.org/10.1111/cea.12142
Aubier M, Thabut G, Hamidi F, Guillou N, Brard J, Dombret MC, Borensztajn K, Aitilalne B, Poirier I, Roland-Nicaise P, Taille C, Pretolani M (2016) Airway smooth muscle enlargement is associated with protease-activated receptor 2/ligand overexpression in patients with difficult-to-control severe asthma. J Allergy Clin Immunol 138(3):729–739.e711. https://doi.org/10.1016/j.jaci.2015.12.1332
Allard B, Bara I, Gilbert G, Carvalho G, Trian T, Ozier A, Gillibert-Duplantier J, Ousova O, Maurat E, Thumerel M, Quignard JF, Girodet PO, Marthan R, Berger P (2014) Protease activated receptor-2 expression and function in asthmatic bronchial smooth muscle. PLoS One 9(2):e86945. https://doi.org/10.1371/journal.pone.0086945
Hirst SJ (2003) Regulation of airway smooth muscle cell immunomodulatory function: role in asthma. Respir Physiol Neurobiol 137(2–3):309–326
Rogers NK, Clements D, Dongre A, Harrison TW, Shaw D, Johnson SR (2014) Extra-cellular matrix proteins induce matrix metalloproteinase-1 (MMP-1) activity and increase airway smooth muscle contraction in asthma. PLoS One 9(2):e90565. https://doi.org/10.1371/journal.pone.0090565
Bergeron C, Tulic MK, Hamid Q (2010) Airway remodelling in asthma: from benchside to clinical practice. Can Respir J 17(4):e85–e93
Halwani R, Al-Muhsen S, Al-Jahdali H, Hamid Q (2011) Role of transforming growth factor-beta in airway remodeling in asthma. Am J Respir Cell Mol Biol 44(2):127–133. https://doi.org/10.1165/rcmb.2010-0027TR
Margulis A, Nocka KH, Brennan AM, Deng B, Fleming M, Goldman SJ, Kasaian MT (2009) Mast cell-dependent contraction of human airway smooth muscle cell-containing collagen gels: influence of cytokines, matrix metalloproteases, and serine proteases. J Immunol 183(3):1739–1750. https://doi.org/10.4049/jimmunol.0803951
Davies AM, Allan EG, Keeble AH, Delgado J, Cossins BP, Mitropoulou AN, Pang MOY, Ceska T, Beavil AJ, Craggs G, Westwood M, Henry AJ, McDonnell JM, Sutton BJ (2017) Allosteric mechanism of action of the therapeutic anti-IgE antibody omalizumab. J Biol Chem 292(24):9975–9987. https://doi.org/10.1074/jbc.M117.776476
Mizuma H, Tanaka A, Uchida Y, Fujiwara A, Manabe R, Furukawa H, Kuwahara N, Fukuda Y, Kimura T, Jinno M, Ohta S, Yamamoto M, Matsukura S, Adachi M, Sagara H (2015) Influence of omalizumab on allergen-specific IgE in patients with adult asthma. Int Arch Allergy Immunol 168(3):165–172. https://doi.org/10.1159/000442668
Cohen ES, Dobson CL, Kack H, Wang B, Sims DA, Lloyd CO, England E, Rees DG, Guo H, Karagiannis SN, O'Brien S, Persdotter S, Ekdahl H, Butler R, Keyes F, Oakley S, Carlsson M, Briend E, Wilkinson T, Anderson IK, Monk PD, von Wachenfeldt K, Eriksson PO, Gould HJ, Vaughan TJ, May RD (2014) A novel IgE-neutralizing antibody for the treatment of severe uncontrolled asthma. MAbs 6(3):756–764. https://doi.org/10.4161/mabs.28394
Caslin HL, Kiwanuka KN, Haque TT, Taruselli MT, MacKnight HP, Paranjape A, Ryan JJ (2018) Controlling mast cell activation and homeostasis: work influenced by Bill Paul that continues today. Front Immunol 9:868. https://doi.org/10.3389/fimmu.2018.00868
Peters SP, Busse WW (2017) New and anticipated therapies for severe asthma. J Allergy Clin Immunol Pract 5s(5):S15–s24. https://doi.org/10.1016/j.jaip.2017.07.008
Verstraete K, Peelman F, Braun H, Lopez J, Van Rompaey D, Dansercoer A, Vandenberghe I, Pauwels K (2017) Structure and antagonism of the receptor complex mediated by human TSLP in allergy and asthma. Nat Commun 8:14937. https://doi.org/10.1038/ncomms14937
Dubreuil P, Letard S, Ciufolini M, Gros L, Humbert M, Casteran N, Borge L, Hajem B, Lermet A, Sippl W, Voisset E, Arock M, Auclair C, Leventhal PS, Mansfield CD, Moussy A, Hermine O (2009) Masitinib (AB1010), a potent and selective tyrosine kinase inhibitor targeting KIT. PLoS One 4(9):e7258. https://doi.org/10.1371/journal.pone.0007258
Koziol-White CJ, Jia Y, Baltus GA, Cooper PR, Zaller DM, Crackower MA, Sirkowski EE, Smock S, Northrup AB, Himes BE, Alves SE, Panettieri RA Jr (2016) Inhibition of spleen tyrosine kinase attenuates IgE-mediated airway contraction and mediator release in human precision cut lung slices. Br J Pharmacol 173(21):3080–3087. https://doi.org/10.1111/bph.13550
Moy LY, Jia Y, Caniga M, Lieber G, Gil M, Fernandez X, Sirkowski E, Miller R, Alexander JP, Lee HH, Shin JD, Ellis JM, Chen H, Wilhelm A, Yu H, Vincent S, Chapman RW, Kelly N, Hickey E, Abraham WM, Northrup A, Miller T, Houshyar H, Crackower MA (2013) Inhibition of spleen tyrosine kinase attenuates allergen-mediated airway constriction. Am J Respir Cell Mol Biol 49(6):1085–1092. https://doi.org/10.1165/rcmb.2013-0200OC
Ramis I, Otal R, Carreno C, Domenech A, Eichhorn P, Orellana A, Maldonado M, De Alba J, Prats N, Fernandez JC, Vidal B, Miralpeix M (2015) A novel inhaled Syk inhibitor blocks mast cell degranulation and early asthmatic response. Pharmacol Res 99:116–124. https://doi.org/10.1016/j.phrs.2015.05.011
Hall IP, Fowler AV, Gupta A, Tetzlaff K, Nivens MC, Sarno M, Finnigan HA, Bateman ED, Rand Sutherland E (2015) Efficacy of BI 671800, an oral CRTH2 antagonist, in poorly controlled asthma as sole controller and in the presence of inhaled corticosteroid treatment. Pulm Pharmacol Ther 32:37–44. https://doi.org/10.1016/j.pupt.2015.03.003
Cahill KN, Katz HR, Cui J, Lai J, Kazani S, Crosby-Thompson A, Garofalo D, Castro M, Jarjour N, DiMango E, Erzurum S, Trevor JL, Shenoy K, Chinchilli VM, Wechsler ME, Laidlaw TM, Boyce JA, Israel E (2017) KIT inhibition by imatinib in patients with severe refractory asthma. N Engl J Med 376(20):1911–1920. https://doi.org/10.1056/NEJMoa1613125
Rhee CK, Kim JW, Park CK, Kim JS, Kang JY, Kim SJ, Kim SC, Kwon SS, Kim YK, Park SH, Lee SY (2011) Effect of imatinib on airway smooth muscle thickening in a murine model of chronic asthma. Int Arch Allergy Immunol 155(3):243–251. https://doi.org/10.1159/000321261
Patel KR, Aven L, Shao F, Krishnamoorthy N, Duvall MG, Levy BD, Ai X (2016) Mast cell-derived neurotrophin 4 mediates allergen-induced airway hyperinnervation in early life. Mucosal Immunol 9(6):1466–1476. https://doi.org/10.1038/mi.2016.11
Wang Q, Chen D, Xie H, Lin X, Wang X, Yao Q, Zheng X, Xu C, Chen L, He S, Zhang H (2016) Altered expression of IFN-lambda2 in allergic airway disorders and identification of its cell origins. Mediat Inflamm 2016:5759496. https://doi.org/10.1155/2016/5759496
Conti P, Ronconi G, Caraffa A, Lessiani G, Duraisamy K (2017) IL-37 a new IL-1 family member emerges as a key suppressor of asthma mediated by mast cells. Immunol Investig 46(3):239–250. https://doi.org/10.1080/08820139.2016.1250220
Bezemer GF, Sagar S, van Bergenhenegouwen J, Georgiou NA, Garssen J, Kraneveld AD, Folkerts G (2012) Dual role of Toll-like receptors in asthma and chronic obstructive pulmonary disease. Pharmacol Rev 64(2):337–358. https://doi.org/10.1124/pr.111.004622
Zhang H, Wang J, Wang L, Xie H, Chen L, He S (2018) Role of IL-18 in atopic asthma is determined by balance of IL-18/IL-18BP/IL-18R. J Cell Mol Med 22(1):354–373. https://doi.org/10.1111/jcmm.13323
Wang J, Zhang H, Zheng W, Xie H, Yan H, Lin X, He S (2016) Correlation of IL-18 with tryptase in atopic asthma and induction of mast cell accumulation by IL-18. Mediat Inflamm 2016:4743176–4743114. https://doi.org/10.1155/2016/4743176
Paivandy A, Sandelin M, Igelstrom H, Landelius P, Janson C, Melo FR, Pejler G (2017) Induction of human lung mast cell apoptosis by granule permeabilization: a novel approach for targeting mast cells. Front Immunol 8:1645. https://doi.org/10.3389/fimmu.2017.01645
Wareham KJ, Seward EP (2016) P2X7 receptors induce degranulation in human mast cells. Purinergic Signal 12(2):235–246. https://doi.org/10.1007/s11302-016-9497-4
Lee AJ, Ro M, Cho KJ, Kim JH (2017) Lipopolysaccharide/TLR4 stimulates IL-13 production through a MyD88-BLT2-linked cascade in mast cells, potentially contributing to the allergic response. J Immunol 199(2):409–417. https://doi.org/10.4049/jimmunol.1602062
Suurmond J, Habets KLL, Tatum Z, Schonkeren JJ, Hoen PAC, Huizinga TWJ, Laros JFJ, Toes REM, Kurreeman F (2016) Repeated FcepsilonRI triggering reveals modified mast cell function related to chronic allergic responses in tissue. J Allergy Clin Immunol 138(3):869–880. https://doi.org/10.1016/j.jaci.2016.01.017
Yoshioka M, Fukuishi N, Iriguchi S, Ohsaki K, Yamanobe H, Inukai A, Kurihara D, Imajo N, Yasui Y, Matsui N, Tsujita T, Ishii A, Seya T, Takahama M, Akagi M (2007) Lipoteichoic acid downregulates FcepsilonRI expression on human mast cells through Toll-like receptor 2. J Allergy Clin Immunol 120(2):452–461. https://doi.org/10.1016/j.jaci.2007.03.027
Suzuki Y, Wakahara K (2017) Airway basophils are increased and activated in eosinophilic asthma. Allergy 72(10):1532–1539. https://doi.org/10.1111/all.13197
Sancho-Chust JN, Chiner E, Camarasa A, Llombart M (2009) Recent-onset bronchial asthma as a manifestation of systemic mastocytosis. J Investig Allergol Clin Immunol 19(6):513–515
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Ethical Approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Informed Consent
No informed consent was required to prepare the manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Elieh Ali Komi, D., Bjermer, L. Mast Cell-Mediated Orchestration of the Immune Responses in Human Allergic Asthma: Current Insights. Clinic Rev Allerg Immunol 56, 234–247 (2019). https://doi.org/10.1007/s12016-018-8720-1
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12016-018-8720-1