
Abstract
Strontium isotope ratios in human teeth from the three Swedish prehistoric Stone Age hunter-fisher-gathering societies Norje Sunnansund (Maglemose), Skateholm (Ertebølle) and Västerbjers (Pitted Ware Culture) were analysed with laser ablation to produce data on both individual movement patterns and societal mobility trends. The analyses of teeth from both Skateholm and Västerbjers displayed homogeneous ratios and corresponding mobility patterns, while the data from Norje Sunnansund showed larger variances with heterogenous strontium ratios and varied inter-individual mobility patterns. Correlation with the bioavailable baseline suggests that the size of the geographical areas, where human strontium ratios could have originated, was roughly comparable for all three sites. The teeth measurements were reflected within a 50-km radius of the surrounding landscape and the 25–75% data quartile matched with distances between 3 and 30 km from the sites, suggesting limited mobility ranges among aquatically dependent foragers from southernmost Sweden. By applying ethnographic analogies and site-specific contextual inferences, the results suggest that mobility ranges at Norje Sunnansund were likely not delimited by neighbouring group territories. This changed over time and an increasing territorialisation of the landscape may have influenced movement patterns and caused restrictions to the foraging activities at both Skateholm and Västerbjers.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The perspectives on movement and mobility patterns among foraging societies have become increasingly nuanced for ethnographic societies while remaining more elusive in archaeological contexts. Recent advances in biomolecular analyses, i.e. ancient (a)DNA analyses, have shown extensive, and rapid, movement among Early Holocene foragers. One example is the colonisation of Scandinavia, which can be seen as evidence of foraging mobility (Günther et al. 2018). However, this type of mobility is large scale, showing the mobility of generations rather than that of individuals, and it mainly concerns the colonization of pristine uninhabited lands. At some point, the novel land becomes inhabited, and new types of mobility patterns are likely to emerge. When this happens, territoriality and level of land ownership may influence, limit or restrict hunter-fisher-gatherer movement. Understanding the spectrum of how foraging societies handle the presence of other groups in their close vicinity may provide a frame for interpreting the complexities behind both individual and societal mobility trends.
Contextualizing strontium ratios
Since the 1980s, strontium isotope ratios in teeth have been used to investigate the origin of both humans and artefacts (Ericson 1985; van der Merwe et al. 1990) allowing studies of individual movement and dispersal, networks and population dynamics (Ericson 1985; Gron et al. 2016; Kjällquist and Price 2019; Montgomery 2010; Montgomery et al. 2003; Price et al. 2002, 1994; Sjögren et al. 2009). Provenience studies based on strontium isotopes are possible because the ratio between 87 and 86Sr is dependent on the age and content of the bedrock in a specific area and because these ratios remain unaltered as it is transferred into topsoils and further on into water, plants and animals. The Sr ratios available for biological specimens (bioavailable Sr) are also affected by the composition of the loose soil, precipitation levels, evapotranspiration, aeolian dust, sea spray and the use of fertilizers (Holt et al. 2021). When animals and humans eat and drink, they incorporate and store Sr isotopes from food and water. The different Sr values from the dietary sources at a specific location form the basis for a model of the bioavailable baseline or a Sr isotope region.
Traditional Sr analyses involve comparisons of bulk Sr isotope ratios in a tooth to local Sr baseline data. When tracking mobility, analyses of different teeth, with varied timing of enamel mineralization, from the same individual have been used (Blank et al. 2021; Haverkort et al. 2008). Recent developments in sampling methods allow a finer resolution, and by micro-sampling a tooth, it is possible to analyse small enamel sections and obtain information from a few weeks of spot-specific mineralization instead of averaged data from the entire tooth. By micro-sampling a tooth, multiple times at an equal enamel depth, from tip to cervix, it is possible to study the measurement pattern and obtain chronologically ordered data where each measurement represents a few weeks of strontium uptake incorporated in spot-specific enamel mineralization (Boethius et al. 2022). If correlated with bioavailable strontium ratios, it is thus possible to follow the mobility of both humans and animals (Boethius et al. 2022; Lazzerini et al. 2021).
The Stone Age forager settlements
To investigate if, how and why the mobility pattern changed and developed over time and between different foraging societies, the individual human mobility pattern from three different south Swedish sites has been analysed and connected to ethnographic analogies and site-specific contextual inferences. The sites have been selected to represent three different cultural epochs, separated by around one and a half millennia each, and are comparable through their location in close vicinity to the Baltic Sea (Fig. 1a). The selected sites are as follows:

Map showing the location of the three investigated sites as seen today. Map of Sweden (a) generated with QGIS 3.10 using the Natural Earth data set. Orthophotos (b–d) from Lantmäteriet. Geographical Coordinates: Norje Sunnansund: 56°6′35.7″N 14°39′33″E. Skateholm: 55°23′10.1″N 13°29′1.5″E. Västerbjers: 57°35′21.4″N 18°42′41.9″E
Norje Sunnansund (Fig. 1b), a late Maglemosean site (9600–8600 calibrated (cal) before present (BP) from Blekinge. It is one of the earliest known sites from the Swedish east coast with evidence of a strong fish dependency (Boethius 2018a, b; Boethius and Ahlström 2018), including seasonal targeted mass fish exploitation (Boethius et al. 2021), fish fermentation practises (Boethius 2016), year-round occupation (Boethius 2017) and craft traditions connecting the inhabitants with both north- and southbound cultural traditions and social networks (Damlien et al. 2018; David and Kjällquist 2018; Kjällquist 2020; Kjällquist et al. 2016; Kjällquist and Price 2019).
Skateholm (Fig. 1c), an Ertebølle site (7200–6800 cal BP) from southern Scania. It is a series of Late Mesolithic coastal settlement/burial sites of which the sites Skateholm I and II are most famous for being the largest known Scandinavian Mesolithic cemeteries, for both humans and dogs, with a total of nearly 90 excavated graves (Alexandersen 1988; Eriksson 2003; Larsson 1988, 1990, 1993, 2016; Nilsson Stutz 2003; Persson and Persson 1988). The site is also known for its large species diversity (Jonsson 1988) with year-round-seasonal indicators (Price et al. 2021).
Västerbjers (Fig. 1d), a Pitted Ware Culture (PWC) site (ca 5000–4600 cal BP) from the island of Gotland. It is a mid-Neolithic burial site with over 50 excavated human graves (Janzon 1974; Stenberger 1939) and a zooarchaeological record dominated by boar (Sus scrofa), seal (Phocidae) and dog (Canis familiaris) (Ekman 1974:Table 3; Stenberger et al. 1943). Human stable isotope data (δ13C and δ15N) suggest a protein diet dominated by marine resources (Eriksson 2004), while recent δ13Capatite data suggest a high dependence on C3 sources (Ahlström and Price 2021). Gotlandic PWC sites show year-round seasonal indicators (Olson 2008; Storå, 2001), facilitating suggestions of sedentary foragers at Västerbjers (Ahlström and Price 2021).
Material
Strontium isotope ratios from 35 teeth were analysed and compared. The teeth were selected based on availability and included 5 teeth from Norje Sunnansund, 11 teeth from Skateholm and 19 teeth from Västerbjers. The teeth were analysed using LA‐MC-ICP‐MS with between 5 and 27 individual measurements (mean 15.5) on each tooth. The enamel area, targeted by the laser ablations, was mineralized when the individuals were children or in their early adolescence (Table 1). All strontium isotope analyses on archaeological specimens were made at the Vegacenter, the Museum of Natural History, Stockholm, Sweden.
Baseline expansion
Plant Sr ratios (Holt et al. 2021; Snoeck et al. 2020) or Sr data from archaeological animals with limited mobility, such as mice (Muridae) and voles (Cricetidae) (Kootker et al. 2016), have been suggested the best way to establish regional bioavailable Sr ranges. Sr ratios from water bodies have also provided Sr ratios similar to Sr data from plants (Blank et al. 2018; Maurer et al. 2012) and may serve as a viable option to determine bioavailable baseline ratios (Boethius et al. 2021). Here, 163 individual bioavailable baseline sources were selected based on their ability to represent local, but not spot-specific, strontium ranges. This includes data from plants, water and archaeological mice, rats, voles, hedgehog (Erinaceus europaeus) and least weasel (Mustela nivalis). Data from large mammals, snails, water from wells or limed lakes and humans were not used. The baseline presented here is based on both previously published data (Ahlström Arcini 2018; Ahlström and Price 2021; Boethius et al. 2021; Fraser et al. 2018; Kjällquist and Price 2019; Klassen 2020; Ladegaard-Pedersen et al. 2021; Larsson et al. 2020; Löfvendahl et al. 1990; Sjögren et al. 2009; Wilhelmson and Ahlström, 2015) and data obtained through this study (Supplementary information (S)1: Table S1). The additions presented here comprise water samples from Gotland, five from natural springs and four from freshwater lakes. On the mainland, three archaeological field vole teeth (Microtus agrestis) and six water vole teeth (Arvicola amphibius) were also analysed through LA‐MC-ICP‐MS, with 2 to 10 ablations each. Their average value was used to form the particular baseline data.
Methods
Ethics
The archaeological tooth specimens included in this study were obtained with permission from Blekinge museum (Norje Sunnansund), the Historical Museum at Lund University (human teeth from Skateholm and rodent teeth from Bökeberg and Tågerup) and The National Historical Museums of Sweden (Västerbjers). All analyses of archaeological specimens have been conducted by laser ablations. This causes minimal damage to the investigated specimens, a ca 400 microns long and 150 microns wide cut in the enamel, which is barely visible to the naked eye.
Baseline water 87Sr/86Sranalyses
Water samples were analysed with the Thermal Ionization Mass Spectrometer (TIMS) at the Museum of Natural History, Stockholm, Sweden, following the method outlined in Boethius et al. (2021). The external precision for 87Sr/86Sr as judged from running 987 standards was 17 ppm. Accuracy correction was applied, the 87Sr/86Sr ratio for the NBS 987 standard was 0.710214 ± 13 (n = 12), which was normalized to an NBS987 87Sr/86Sr ratio of 0.710248 following (McArthur 2010), see S2: Table S1.
Laser ablation
The teeth were first checked for prior varnish treatment. When detected on the human teeth from Skateholm, the treated teeth were cleaned with acetone to remove the applied substance. All teeth were soaked and washed in distilled water and cleaned with a soft toothbrush. Following cleaning, the teeth were air-dried at room temperature, after which they were transported to Vegacenter, where they were mounted and fixed in a sample cell on a movable mounting table.
87Sr/86Sr ratios were obtained by measuring the enamel of each tooth using an NWR193 excimer laser ablation system (Elemental Scientific Lasers, Bozeman, USA) connected to a Nu Plasma II multi‐collector ICP mass spectrometer (Nu Instruments Ltd, Wrexham, UK). The teeth were measured on four separate occasions (November 2019, February, March and June 2020). The instrument operating conditions for these occasions are listed in the supplementary information (S2 Tables S2–4) followed by information on the two standards; a spine from a velvet belly lantern shark (Etmopterus spinax), used for normalization, and a tooth from a mountain hare (Lepus timidus), used to verify that all interferences, had been successfully removed (S2 Table S5–8).
Nine rodent teeth were ablated to expand the baseline (S2 Tables S9–17 and Figs. S1–9). The isotope data for each of the 35 ablated human teeth is given in S2 Tables S18–52 along with close-up photographs of the ablation marks (S2 Figs. S10–44). To obtain chronological, ordinal scale, measurements, all ablations were made at equal enamel depths following the development of the tooth, i.e. starting at the tip and moving down towards the cementum–enamel junction (Boethius et al. 2022). To avoid potential enamel surface contamination and to ensure each measurement was done at comparable depths, each tooth was pre-ablated using a laser spot size of 150 μm where the outer ̴10) µm of the enamel were removed (cf. instrumental settings S2 Tables S2–4). Parallel tracks of ca 400 μm were then ablated for Sr analysis, using a 130 μm spot size, on the pre‐ablated enamel surface (see Fig. 2). The results do not show an increased concentration of the rare earth element ytterbium (Yb) or rubidium (Rb), compared to the standards, which could indicate contamination. The often drastic changes in 87Sr/86Sr ratios when dentine was struck are further indications that the enamel has not been contaminated by its depositional soil environment, while the dentine was. Other forms of undetected diagenesis might have affected the enamel, but if that is the case, previous bulk analyses by TIMS have equally failed to identify it (Boethius et al. 2022).

Photographs of three teeth, one from each investigated site, showing the marks left by the laser ablation. Left: first lower molar from Norje Sunnansund (NS:102). Middle: lower canine from Skateholm II (SKII:104). Right: first upper premolar from Västerbjers (VB:41)
In a few cases, the ablations close to the cementum-enamel junction included dentine. This was assessed visually by studying close-up photographs of each ablated tooth (S2 Figs. S1–44) and through divergence in 87Rb/86Sr ratios, as this ratio often changes when dentine or bone is ablated (cf. S2 Tables S9–52). This was observed on two teeth from Norje Sunnansund (NS:100, three ablations and NS:103, one ablation), one time on a tooth from Skateholm (SKI:114) and four times on teeth from Västerbjers (VB:40, two ablations and VB:42, two ablations). In these cases, the measurements were not included in the data interpretation. This was deemed the proper action both because the LA‐MC-ICP‐MS method has been developed for measuring tooth enamel and the reference standards verify measurements on tooth enamel and because dentine has a lower density and more porous structure than enamel (Montgomery 2010) and can be more easily contaminated (Hoppe et al. 2003).
On two of the teeth from Västerbjers, VB:45 and VB:52, the stable 84Sr/86Sr ratio, which was used as further accuracy control, was inaccurate by more than 1000 ppm as established by Thirlwall (1991), which could indicate an insufficient correction of interferences or matrix effects from surface contaminations. The 87Sr/86Sr ratios from both of the teeth fit well with the data from the other teeth and have been included in the analyses while acknowledging that some minor offset might be inflicted on the measurements, as signalled by the slight offset of the measured 84Sr/86Sr ratio.
Isobaric interferences, if present, were corrected by subtracting a gas blank (84Kr) and peak stripping doubly charged REE, Ca-dimers/argides, 87Rb (Glykou et al., 2018). All corrections were applied online, which resulted in interference-free 87Sr/86Sr ratios for each data point (S2 Tables S9–52). Polyatomic interference on m/z 87 has also been reported (Horstwood et al. 2008) and is described as (Ca/Ar)31P16O+. This interference can introduce a significant offset in 87Sr/86Sr, especially for samples with low Sr concentrations, which can be reduced by a thorough low oxide tuning of the gases (Willmes et al. 2016). At Vegacenter, accurate 87Sr/86Sr ratios have been demonstrated down to CSr ~ 7 ppm under optimal tuning conditions (Boethius et al. 2021).
Results
Bioavailable baseline
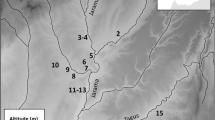
The bioavailable 87Sr/86Sr ratio baseline is disparate in the area of study. The reason for regional variations may be visualized when the data is presented on a bedrock map (Fig. 3) and thus provide a frame for understanding the human data.

Bioavailable 87Sr/86Sr baseline map of southern Sweden displayed on a 1:1 M bedrock map from SGU. Bedrock types covering a larger area are numbered. Each letter corresponds to a baseline sampling point. * indicates averaged data from a specific location where multiple analyses have been made/are available. Explanations of each measurement and its specifics in S1 Table S1
Human87Sr/86Sr isotope data
A total of 532 enamel analyses on human teeth were used in this study. The 87Sr/86Sr ratio range from 0.70950 to 0.72528, depending on the site and specific tooth (Table 2).
The results of the human Sr ratios reflect the diet during childhood or early adolescence and represent individual mobility within or between various Sr isotope regions or the Sr ratios of food from various areas brought back to the settlement (Fig. 4). Four main types of mobility (and different variations of them) could be interpreted from the data. The “low mobility” type, based on more or less horizontal trendlines (e.g. NS:102, SK:105 or VB:50). The “on the move” type, based on trendlines showing a decrease or increase in values (e.g. NS:101 or VB:34). The “back and forth” type, based on curved data with one peak or valley and end values at similar (or moving towards similar) 87Sr/86Sr ratios (e.g. SK:112 or NS:100). The “full transhumance” type, based on a full sinus shaped curve including both a peak(s) and bottom(s) with end values at similar ratios (e.g. NS:103).

Individual mobility during enamel mineralization, indicated by the variation of Sr ratios on each investigated tooth with individual data points from all valid measurements. NS = Norje Sunnansund, SK = Skateholm, VB = Västerbjers. The first number is the individual tooth identity. G followed by Roman or Hindu-Arabic numerals indicates a specific grave. The error bar on each ablation shows the 2SD external precision propagated from external reproducibility (2SD) obtained from the primary standard during the analytical session combined with the within-run precision of each analysis (2SE). Ablations striking dentine excluded; see S2 Figs. S10–44 for information and exact data for each measurement
Strontium variations are most pronounced in the Norje Sunnansund teeth, where data from the different teeth show a combination of high ( ̴ > 0.720), mid-range ( ̴ 0.713–0.720) and low ( ̴ < 0.713) 87Sr/86Sr ratios. As indicated by the sequence of the measurements, the values from the teeth with both the highest and lowest Sr ratios (NS:101, NS:102) are moving towards the mid-ranged ratios within the period of enamel mineralization. The values from the three “mid-ratio teeth” (NS:99, NS:100, NS:103) indicate mobility patterns suggesting a more back and forth/transhumance kind of mobility in geographical areas with roughly similar Sr ratios. None of the teeth from Norje Sunnansund display similar Sr mobility patterns with similar isotope ratios, and no trends are detectable in the dataset. All types of movement patterns are represented, both “low mobility”, “on the move”, “back and forth”, and “full transhumance”.
The teeth from Skateholm, in general, show homogenous and low 87Sr/86Sr ratios. They have, with two main exceptions, the least varied values and mobility trends connected to a homogenous and low 87Sr/86Sr bioavailable baseline. The type of movement pattern represented is thus mainly “low mobility”. One tooth (SK:112) has values that indicate a back-and-forth pattern with the mid tooth ablations elevated above all other human measurements from the site. The other exception is a tooth (SK:113) whose values, although with ratios within the local span, could indicate transhumance, having moved back and forth within the local Sr isotope ranges.
The teeth from Västerbjers, in general, also show homogenous and low 87Sr/86Sr ratios. The individual mobility signals from the Västerbjers teeth are somewhat more elevated than the Skateholm teeth (Fig. 5). The patterns of the values are dominated by indications of low mobility, suggesting a uniform diet from homogenous Sr isotope regions, as well as patterns of back-and-forth mobility, suggesting temporal shifts and subsequent returns to a diet with slightly different Sr ratios. Almost all teeth show similar mobility patterns with the main exception of one tooth (VB:34), where the pattern seems to indicate a move from a different Sr isotope region towards a region with values similar to the other teeth from Västerbjers.

Mobility signals for the various teeth. Illustrated as the mean and standard error of the mean, for the combined enamel ablation data set from each tooth
The similarity between the 87Sr/86Sr ranges among the individuals from Västerbjers and Skateholm is caused by similar bedrock in both regions and corresponds to similar bioavailable baseline ranges on Gotland and in south-western Scania (Fig. 3). Despite similar bioavailable Sr ranges in the two regions, the individual Västerbjers tooth data is more heterogeneous and show somewhat elevated data ranges during enamel mineralization, although not as large as those observed for the Norje Sunnansund individuals (Fig. 5).
Discussion
When studying the various plots of human 87Sr/86Sr ratios (Figs. 4 and 5), the first impression suggest humans of diverse origin at Norje Sunnansund and local individuals at Skateholm and Västerbjers. However, to fully understand the observed human strontium patterns, the data needs to be considered in the context of differences in local bedrock, possible variations in bioavailable baseline and diet.
Both Skateholm and Västerbjers are located in a homogenous geological landscape consisting of sedimentary bedrock dominated by limestone. This is not the case for Norje Sunnansund, which lies in a diverse geological landscape with high local bioavailable variability (Fig. 3). The differences in bioavailable Sr near the three sites are evident if illustrated with arbitrary borders set at a 25- and 50-km radius (Fig. 6a). If relating the bioavailable baseline data to the measured human data, the datasets do, in general, match well (Fig. 6b). This suggests that despite large inter-site variations, the human Sr ratios from all three sites are primarily derived from the surrounding landscape.

Arbitrary set Sr isotope ranges show the bioavailable baselines at a radius set at 25 and 50 km from each site. a Sr isotope ranges illustrated with the distances added to the general baseline map (see S1 Table S1 for exact data of the baselines samples covered by the ranges); b the bioavailable Sr isotope ratios set in relation to the human 87Sr/86Sr ratios. Boxplots showing the 25–75% data range quartiles (the boxes), with the median shown as the horizontal line within the boxes. The whiskers represent the total data range of all measurements in each category. For bedrock information in 6a, see Fig. 3
There is a general, but not perfect, overlap between the local bioavailable Sr isotope ranges and the human Sr data from all sites (Fig. 6b). The 25–75% quartiles of the values from the human teeth are lower than the corresponding baseline 25–75% quartiles for all three sites. Part of this can likely be attributed to the available baseline data and a non-uniform coverage for the areas surrounding the sites. However, lower Sr ratios are present in the close vicinity of each of the sites, through the ratios from the Baltic Sea, whereby possible aquatic influence on the human Sr ratios needs consideration.
Plants have a higher concentration of Sr compared to water and animal flesh, and humans with an omnivorous diet receive the majority of their strontium from the plants they have consumed (Bentley 2006; Montgomery 2010). It is not until meat accounts for more than 50% of the diet that its signals noticeably affect human strontium ratios (Bentley 2006). Through ethnographic data, it is known that high latitude foragers often base their diet on terrestrial hunting and, in particular, fishing (Marlowe 2005). Considering that all three investigated societies have a strong aquatic orientation, it is plausible that resources from the Baltic Sea have also affected the observed ratios. Indeed, recent studies show that flesh from fish may contain large Sr concentrations, sometimes at the same level or exceeding those from plants, and if the entire fish was consumed (including the bones), the Sr concentrations would be even higher. This, when individual Sr data were also correlated with δ13C and δ15N isotope data, has led to suggestions that aquatic resources may, under the right conditions, affect human Sr ratios more than previously suspected (Lahtinen et al. 2020). Applied to the sites investigated here, the Sr data may thus be suggested to have been influenced by a large proportion of the diet stemming from the Baltic Sea, which has lower Sr ratios than the terrestrial areas (Fig. 3).
The heterogeneous mobility patterns observed between the different individuals at Norje Sunnansund and the homogenous patterns observed at Skateholm and Västerbjers are interesting. While the human Sr values are closely related to the local bioavailable Sr ranges at all three sites (Fig. 6b), the observed differences in mobility patterns may suggest a more uniform or “organized” mobility at Skateholm and Västerbjers and less structured mobility at Norje Sunnansund. The inferences are complicated by the highly variable Sr baseline in the areas surrounding Norje Sunnansund and the more homogenous Sr baseline values around both Skateholm and Västerbjers. The different teeth sampled, representing different ages of tooth mineralization, may also complicate comparisons, and these possible sources of error must be considered. The diverse ablation data from the Norje Sunnansund individuals may represent seasonal or temporally limited movement. It may also represent seasonally or temporally obtained food from different Sr isotope regions within a limited area in the ecotone and diverse landscape surrounding the site, combined with food from seal hunting and fishing trips into the Baltic Sea. Chronological changes or personal preferences are also conceivable. Regardless of why the mobility patterns are disparate, it suggests that the foraging ranges, e.g. the hunting grounds, mushroom locations or hazelnut gathering areas, varied temporally, individually, seasonally or chronologically at Norje Sunnansund, while they remained the same at the two other sites. The difference in site-specific inter-individual mobility patterns, between the Maglemose site and the Ertebølle and PWC sites, may indicate a structural change in mobility patterns—going from more unorganized and individual mobility at the earliest site to more organized and structured mobility patterns at the two latter settlements.
Contrasting bioavailable baseline data with multiple Sr measurements from a human tooth enables estimates of where the bioavailable Sr incorporated in the human enamel may have come from. This will enable discussions of mobility patterns, at least for groups with low mobility. Unfortunately, some areas in proximity to the investigated archaeological sites, e.g. Småland in mid-southern Sweden, are still uninvestigated and lack bioavailable Sr data. This complicates the analyses of forager mobility patterns. Nevertheless, if relating the information from the bioavailable baseline map (Fig. 3) to human Sr isotope ratios from each site (Fig. 4), a mobility range around each settlement may be suggested. When correlating human data to the bioavailable Sr dataset (S1 Table S1), the main human mobility ranges can be illustrated as ranges where the measurements obtained from the human enamel could have originated (Fig. 7). When all human Sr values (Fig. 6b, whiskers) are included and related to the values of the baseline map, the range for each site is about 30–50 km (Fig. 7, red ellipses). A smaller Sr-range, corresponding to the 25–75% quartile dataset of the human measurements from each site (Fig. 6b, boxplots), gives mobility ranges of about 3–30 km (Fig. 7, blue ellipses). Although somewhat biased by limitations in available bioavailable baseline data and variations in baseline coverage for different areas, the size differences between these ranges demonstrate that it is possible to obtain information on ancient foraging mobility patterns (Fig. 7).

Model showing two schematically marked possible Sr origin ranges for each of the three archaeological sites. The ellipses mark a range where the current bioavailable baseline Sr data overlap with the human teeth data from each site (cf. Figure 3 and S1 Table S1 for exact data of each baseline measurement). The blue ellipse marks the geographically smallest area for each site where Sr values corresponding to 25–75% of the quartile data range of human Sr (Fig. 6b) could have originated. The red ellipse marks the smallest geographical area for each site where all of the human Sr values could have originated (max/min whiskers in Fig. 6b). Dashed lines indicate a level of uncertainty or insufficient baseline data, and dashed arrows indicate possible Baltic Sea voyages of an unknown extent. a shows the sites Norje Sunnansund and Skateholm, b shows Västerbjers with two alternative blue ellipses; one range on northern Gotland, including only terrestrial baseline data versus one range adjacent to the site that is relevant if human Sr ratios were significantly affected by the lower Baltic Sea Sr values, through a fish and seal oriented diet. c shows the overall geographic location of the illustrated areas. Maps generated with QGIS 3.10 using the Natural Earth data set
Because of the diverse bedrock and the corresponding wide range of bioavailable Sr ratios in the area around Norje Sunnansund, the elevated Sr isotope ranges from the human teeth fit well with bioavailable Sr ratios from the surrounding landscape. The uniform Sr data from the Skateholm individuals also offers indications of limited mobility ranges. However, here, the homogenous bioavailable baseline in the area surrounding the site hampers the efforts to single out specific human mobility patterns, and the human Sr isotope data suggest mainly local mobility without more extensive journeys to areas of different isotope regions. At Västerbjers, the human isotope data overlaps with the general bioavailable baseline of Gotland, but not with the baseline for the local area around Västerbjers itself. This is interesting and might be related to plant foraging activities in the northern part of the island. However, considering elevated human 15 N values and suggestions of a strong aquatic dietary orientation at Västerbjers (Eriksson 2004), it most likely reflects a generally elevated consumption of fish and seals from the Baltic Sea.
Through this, our understanding of diachronic mobility patterns among Stone Age hunter-fisher-gatherers from southern Sweden has taken a step forward. The data shows that the Sr in the human teeth could have originated at roughly similar distances from the sites, indicating limited foraging mobility, at least on some sites, from the end of the Early Mesolithic period and onwards. However, since the inter-individual mobility patterns had changed from heterogeneous at Norje Sunnansund to homogenous at Skateholm and Västerbjers, further considerations and reflections are warranted.
Connecting ancient mobility ranges with ethnographic foraging data
Generalizing the ethnographic foraging spectrum
Ethnographic forager mobility is not optimal for comparisons with mobility patterns among ancient foragers, and direct parallels cannot be established between historical events and archaeological data. Ethnographic accounts do, however, offer some insight into how mobility ranges, territorial restriction, descent lineage ownership and sharing of resources vary with the resource predictability and productivity of a region (Rowley-Conwy and Piper 2016). Ethnographies also illustrate the known range of strategies and human relations connected to foraging mobility and possible restrictions caused by group territories.
Forager territoriality involves systems of both hard and soft boundaries (Grøn, 2015), and there are many aspects of movement patterns and territorial use in the ethnographic foraging spectrum. The specific territorial system practised in a particular region may be of pivotal importance for how the individuals within these societies moved, the distances they travelled from the settlements and what areas of the landscape they visited or did not visit.
At one end of the ethnographic foraging spectrum, a relaxed attitude towards the group’s area or possible territories causes few restrictions to individual mobility. Hunter-gatherers from regions where resources are evenly spread throughout the year, e.g. the Hadza from Tanzania (Marlowe 2010) and the Ju/'hoansi-!Kung from northern Namibia/southern Angola (Lee 1984), do not appear to practise a tight control of specific territories. While certain areas, such as a water hole, might be called one’s own, there would be no hesitation in sharing it with others (Kelly 2013). From a vastly different landscape in northern Canada, Inuit, e.g. the Netsilik, also do not appear to be practising territorial ownership (Balikci 1970). Here, the resources are possibly too scattered and unreliable for the groups to benefit from territorial behaviour (Rowley-Conwy and Piper 2016), and resource sharing is often seen among Inuit populations (Gubser 1965).
Expanding into a form of territorial practise, accounts from the Northern Territories in Australia suggest that Aboriginal territoriality consists “…principally of exclusive access to, and the responsibility to look after, places of spiritual significance; the so-called ‘sacred sites’” (Layton 1986: 22). Since the restrictions do not primarily involve food procurement, this would probably also suggest that the restrictions on individual mobility were few.
Accounts from the North American west coast offer insight into a wide array of more “strict rules” connected to territoriality and elite resource ownership, suggesting seasonal restrictions to individual mobility. Accounts from the Twana suggest seasonal territoriality where people were allowed to move freely through the landscape for roughly half of the year. In time for the salmon migrations in the fall, the landscape and access to particular sections and individual fish traps became limited and connected to the territory of the group residing in the associated winter settlement (Elmendorf 1960). These winter settlements could, around the beginning of the nineteenth century, have included comparably large populations of between several hundred people to over 1,000 individuals (Elmendorf 1993), implying a large foraging dispersal during the warmer months of the year.
Yet other forms of territorial ownership indicate how major territorial restrictions would limit individual mobility. Among the Nootka of Vancouver Island, all land was owned, and the owner of a particular area was entitled to the first yield of the season, after which procurement rights were distributed to the group. Outsiders were prohibited from exploiting the land, unless able to provide proof of kinship (Drucker 1939). Among the Kwakiutl, Boas (1966) describe how both hunting grounds and berry picking grounds were owned and guarded and that trespassers were killed. Accounts of severe punishments for trespassing are numerous among the North American west coast societies (Binford 2001), and correlations between group size, land ownership, interpersonal violence and warfare among aquatically focused foragers in temperate regions have been highlighted (Codding et al. 2019).
Using collated ethnographic data (Binford 2001), it may be possible to draw more general conclusions related to the level of land ownership, population densities and size of occupation area. This may facilitate interpretations of prehistoric mobility by showing the spectrum of how these parameters have affected the mobility pattern of foragers with fish as their primary base of subsistence and who practise some form of landownership (Fig. 8). As shown, the size of the occupied area correlates with increasing populations among foragers practising the least amount of land ownership, while the trend is different when more centralized or elite land ownership is practised. For the latter categories, the size of the occupation area increases more slowly when populations increase, suggesting increases in populations primarily lead to increasing population densities and not area expansions (Fig. 8).

Graph showing area occupied by an ethnolinguistic group (AREA) and the total population size of the ethnolinguistic group (TLPOP). The graph includes all hunter-fisher-gatherer societies that are classified as having fish as their primary subsistence base (SUBSP category 3) and who practise any form of land ownership according to Binford’s (2001) data. Owner 2 is defined as: “the local group claims exclusive use rights over resource location, residential sites, while households may claim special trees and similar features of the landscape”. Owner 3 is defined as: “the local group claims for hunting areas, dominant animals, fishing sites, and animal drive locations, such claims administered by a group leader, but smaller segments may claim exclusive access to resource locations”. Owner 4 is defined as “elite ownership of land and resources” (Binford 2001:426)
The ability to store food may also increase population densities in foraging societies without increasing the size of the territories, as it enables them to maximize the yield from local resource patches (Freeman and Anderies 2015). Interestingly, increases in population densities are often considered a driver for increasing territorial behaviour (Moritz et al. 2020), leading to violence and warfare among non-egalitarian foraging societies. Or, as Kelly puts it, “War appears when mobility is not an option” (Kelly 2013:205). Furthermore, territorial claims to land are, among ethnographic societies, often suggested to be motivated by ancestral lineage (Rowley-Conwy and Piper 2016). Formal disposal areas of the dead have been hypothesized as crucial for claiming land rights and ownership of its resources (Saxe 1970). This suggests that if there is a formal and exclusive disposal area of the dead, the culture likely has a corporate group structure in the form of a lineal descent system (Goldstein 1981). Not all foraging societies claim their right to a particular region through their mortuary practices, but if cemeteries exist, they are often located in visible locations along travel routes or at special locations within the group territory (Conolly 2018; Grøn, 2015).
Understanding the diachronic mobility trends of southern Sweden
The ethnographic accounts serve as an illustration of the diversity among foraging societies, while highlighting that inferences on the diet, storage capacity, territorial markers and high levels of observable violence may facilitate the interpretation of ancient foraging mobility patterns.
Recent estimations, based on archaeological site location and ethnographic population data among terrestrial-oriented foragers, suggest low population densities in the Late Palaeolithic and initial Early Mesolithic Scandinavia (Lundström et al. 2021). However, fish-dependent foraging societies can, given the right circumstances, increase to large numbers of people (Binford 2001; Marlowe 2005). Specific aquatically oriented sites, such as Norje Sunnansund, may, consequently, have harboured comparatively large populations. If the general low population estimations in the Early-Middle Mesolithic hold true, it is plausible that general population densities had not increased significantly by the time Norje Sunnansund was in use. Human populations may, under such circumstances, be centred on fewer sites that not yet have reached their maximum population capacity.
The size of the Norje Sunnandsund human mobility range (Fig. 7) could suggest that the indicated part of the landscape provided the resources needed to sustain the population at the settlement. This suggests that more remote food procurement trips were, in general, not needed. This assumption is enhanced by the indications of large-scale storage practices (Boethius 2016), targeting seasonally abundant fish (Boethius et al. 2021), which would have provided an access to staple food during the leaner months of the year (Boethius 2018b). Low general Early Mesolithic population densities would also indicate that the landscape outside of the Norje Sunnandsund area was largely unoccupied or that mobile, hunting- and gathering-oriented, groups utilized it. Consequently, there might not have been any need for territorial manifestations, as seen among the aquatically oriented foragers in Northwestern America (cf. e.g. Codding et al. 2019). This also fits well with the contextual information from the site. Human remains may (or may not) have been visually displayed at the site (Boethius 2018b:119–124), but, if they were, it was not done separately from other activities, and no uniform way of treating the dead is observable in the archaeological material. Following the Saxe-Goldstein hypothesis, this may indicate a less structured form of a lineage descent system (Goldstein 1981). Less structured territories may be the cause of the disparate mobility patterns observed among the individuals at Norje Sunnansund, as no overarching societal structures and no neighbouring group territories dictated where and when you were allowed to hunt, fish or gather.
The initial low levels of landscape territorialisation may have come to change over time. In areas where foraging groups increased exploitation of predictable and abundantly reoccurring resources, e.g. fish migration and spawning, sedentary lifestyles could gradually have emerged, as is often suggested among aquatically oriented foragers in temperate areas (Kelly 2013), and populations would increase. Sedentary aquatic-dependent societies in high population density regions would, if following the ethnographic data models (Codding et al. 2019), have wanted to maintain territorial boundaries to ensure the procurement of plants and terrestrial animals needed for a functional society. With increasingly more parts of the landscape becoming occupied, remaining mobile groups would have been harder pressed and had more difficulties maintaining a mobile life. This could have led to increasingly aquatically dependent societies, a landscape divided into territories, increasing interpersonal violence and expansion into previously unoccupied areas.
The size of the mobility ranges, as indicated by the strontium data from human teeth (Fig. 7), could be related to the area needed to provide the population with the required resources. With increasing general populations filling the landscape, the various foraging groups would become more densely packed. If considering population increases related to aquatic exploitation in temperate zones (Binford 2001; Kelly 2013; Marlowe 2005), suggestions of larger populations at the end of the Mesolithic period (Lewis et al. 2020) and the expansion of Neolithic farmers into Scandinavia, it suggests that there could have been a different need to manifest territoriality and “claim” their land at the two later sites compared to at Norje Sunnansund. The cemeteries at Skateholm (Larsson 1989) and Västerbjers (Janzon, 1974; Stenberger 1939) are good examples of territorial markers that may connect the right to the land with an ancestral lineage (Layton and Rowley-Conwy 2013; Rowley-Conwy 1998). Territoriality may also be attributed to high levels of interpersonal violence, which is well documented from the Mesolithic period cf. e.g. (Arwidsson 1979; Bennike 1997; Gummesson et al. 2018) as well as among Mid-Neolithic PWC foragers on Gotland (Ahlström and Molnar 2012). At Skateholm, there is even evidence of scalping (Ahlström, 2008), which, considering the close connection between scalping and warfare (Steadman 2008), further implies high levels of territorialisation and, by extension, high population densities (Moritz et al. 2020). The homogenous mobility patterns and uniform Sr ranges observed among the investigated individuals from both Skateholm and Västerbjers may be considered in light of these territorial manifestations, which may imply that overarching societal structures and neighbouring group territories restricted movement. Thus, “territorial systems” may have dictated where individuals were allowed to move and when different resources could safely be exploited.
Conclusions
By analysing Sr isotope ratios on multiple spots of human enamel from 35 forager teeth originating from three south Swedish Stone Age settlements, valuable new information was gained. The sampling via laser ablation was done at comparable depths from the enamel surface. The first measurement was taken closest to the tip of the tooth, and the following measurements were taken in subsequent order along the side of each tooth with the last measurement taken closest to the cementum enamel junction at the cervix. This methodological approach gave measurements where each data point represents a slightly older mineralization age than the previous one.
Tracking the Sr ratios over a temporally limited and chronologically ordered period gave detailed information about the Sr ratios from small parts of the enamel. This information was related to bioavailable Sr ranges from southern Sweden. The data was interpreted by using generalized inferences from ethnographic societies, general archaeological theory and previously published information on the three investigated societies.
This study has shown that from the end of the Early Mesolithic period, some societies had a limited mobility pattern. The human Sr ranges from all three sites matched well with the local baselines. Almost all measurements overlapped with baseline values from within a 50-km radius from the sites, often less, and a more limited Sr ratio, corresponding to 25–75% of the quartile data range for the human Sr values, produced radii of about 3–30 km. The measurement patterns generated for each tooth also provided useful information. The data from Norje Sunnansund displayed heterogeneous inter-individual mobility patterns with different Sr ranges and unique patterns for each tooth. The opposite was true for the data from Skateholm and Västerbjers, where homogenous inter-individual mobility and similar Sr ranges could be observed. The differences were interpreted as an indication that significant changes had occurred during the several millennia of separation between the sites.
The human data from Norje Sunnansund was interpreted as belonging to a fairly sedentary group. No major territorial limitations or boundaries were considered to have affected the movement of the inhabitants at the site. The overlap of human data with the local landscape’s bioavailable Sr ranges at Norje Sunnansund, combined with heterogeneous inter-individual mobility patterns and unique patterns for each tooth, indicated mobility or immobility by choice. The mobility patterns at Skateholm and Västerbjers were differently interpreted. Both of these societies were interpreted as mainly sedentary. However, here the overlap of human data with the local bioavailable baselines and the highly homogenized inter-individual mobility pattern was interpreted as representing foraging activities within limited territories. Late Mesolithic and Middle Neolithic mobility was thus seen as representing forced mobility or immobility. The force in question relates to movement restriction caused by societal systems dictating where and when different resources could be exploited—implying that seasonal restrictions and territorial borders mainly defined where foraging activities were conducted.
All three sites show human Sr ratios influenced by Sr from the Baltic Sea. Apart from highlighting the importance of aquatic resources at the investigated sites, this suggests that Sr input from marine and freshwater diets may affect Sr ratios in omnivorous species. Consequently, aquatically derived Sr may affect both the interpretation of the mobility pattern of human tooth data and the ability to determine local and non-local individuals. Thus, if dietary input from aquatic sources is suspected, aquatic Sr should also be considered as a source of Sr and be included in bioavailable Sr calculations.
The exact nature of how different neighbouring Stone Age foraging societies affected each other remains open for debate. High levels of observed interpersonal violence suggest the relations might not have been entirely frictionless. The ability to investigate individual forager movement patterns and societal mobility ranges, and connect them to inferences on territoriality, diet and storage, is important for our understanding of prehistoric foraging societies. As it currently stands, it is not possible to come any closer to the “real” foraging mobility patterns than this through Sr-analysis on human teeth. Although our current understanding of ancient foraging territories is limited, further expansion of bioavailable baselines and research into the provenience and mobility patterns of the hunted, fished and gathered food is, in the near future, likely to expand the knowledge on these matters.
Data availability
All data is available in the main document, in the supplementary information or, as referred to, in previous publications.
Code availability
Not applicable.
Change history
09 October 2022
Missing Open Access funding information has been added in the Funding Note.
References
Ahlström Arcini C (2018) The Viking Age: a time with many faces. Oxbow books, Oxford
Ahlström T (2008) An early example of scalping from the Mesolithic cemetery Skateholm, Sweden. In: Piek J, Terberger T (eds) Traumatologische und pathologische Veränderungen an prähistorischen und historischen Skelettresten – Diagnose, Ursachen und Kontext. Marie Verlag, Archäologie und Geschichte im Ostseeraum, Rahden, pp 59–66
Ahlström T, Molnar P (2012) The placement of the feathers: violence among sub-boreal foragers from Gotland, central Baltic Sea. In: Schulting R, Fibiger L (eds) Sticks, Stones, and Broken Bones: Neolithic Violence in a European Perspective, Oxford. Oxford University Press, Oxford, pp 17–33
Ahlström T, Price TD (2021) Mobile or stationary? An analysis of strontium and carbon isotopes from Västerbjers. Gotland Sweden. J Archaeol Sci Rep 36:102902
Alexandersen V (1988) Description of the human dentitions from the Late Mesolithic grave-field at Skateholm, Southern Sweden. In: Larsson L (ed) The Skateholm Project. I. Man and Environment, Lund: Regiae Societatis Humaniorum Litterarum Lundensis LXXIX, pp 106–163
Arwidsson G (1979) Stenåldersmannen från Stora Bjärs i Stenkyrka. In: Falck W, Nylén E, Nylén K, Schönbäck B & Svahnström K (eds) Arkeologi på Gotland. Visby: Gatlandica, pp 17–23
Balikci A (1970) The Netsilik Eskimo. Natural History Press, New York
Bennike P (1997) Death in the mesolithic. In: Pedersen L, Fisher A, Aaby B (eds) The Danish Storebælt since the Ice Age: Man, Sea and Forest. AS Storebæltsforbindelsen, Copenhagen, pp 99–108
Bentley RA (2006) Strontium isotopes from the earth to the archaeological skeleton: a review. J Archaeol Method Theory 13:135–187
Binford LR (2001) Constructing frames of reference: an analytical method for archaeological theory building using ethnographic and environmental data sets, Berkeley: University of California Press
Blank M, Sjögren K-G, Knipper C, Frei KM, Malmström H, Fraser M, Svensson EM, Günther T, Yngve H, Jakobsson M, Götherström A, Storå J (2021) Mobility patterns in inland southwestern Sweden during the Neolithic and Early Bronze Age. Archaeol Anthropol Sci 13:64
Blank M, Sjögren K-G, Knipper C, Frei KM, Storå J (2018) Isotope values of the bioavailable strontium in inland southwestern Sweden—a baseline for mobility studies. PLoS ONE 13:e0204649
Boas F (1966) Kwakiutl ethnography, edited by Codere, H. Chicago: University of Chicago Press
Boethius A (2016) Something rotten in Scandinavia: the world’s earliest evidence of fermentation. J Archaeol Sci 66:169–180
Boethius A (2017) Signals of sedentism: faunal exploitation as evidence of a delayed-return economy at Norje Sunnansund, an Early Mesolithic site in south-eastern Sweden. Quatern Sci Rev 162:145–168
Boethius A (2018a) The use of aquatic resources by Early Mesolithic foragers in southern Scandinavia. In: Persson P, Skar B, Breivik HM, Riede F, Jonsson L (eds) The ecology of early settlement in Northern Europe - conditions for subsistence and survival. Sheffield: Equinox, pp 311–334
Boethius A (2018b) Fishing for ways to thrive: integrating zooarchaeology to understand subsistence strategies and their implications among Early and Middle Mesolithic southern Scandinavian foragers, Acta Archaeologica Lundensia Series in 8°, vol. 70. Studies in Osteology 4, Lund: Lund University
Boethius A, Ahlström T (2018) Fish and resilience among Early Holocene foragers of southern Scandinavia: a fusion of stable isotopes and zooarchaeology through Bayesian mixing modelling. J Archaeol Sci 93:196–210
Boethius A, Ahlström T, Kielman-Schmitt M, Kjällquist M, Larsson L (2022) Assessing laser ablation multi-collector inductively coupled plasma mass spectrometry as a tool to study archaeological and modern human mobility through strontium isotope analyses of tooth enamel. Archaeol Anthropol Sci 14:97
Boethius A, Kjällquist M, Kielman-Schmitt M, Ahlström T, Larsson L (2021) Early Holocene Scandinavian foragers on a journey to affluence: mesolithic fish exploitation, seasonal abundance and storage investigated through strontium isotope ratios by laser ablation (LA-MC-ICP-MS). PLoS ONE 16:e0245222
Buikstra, J. E. & Ubelaker, D. H. 1994. Standards for data collection from human skeletal remains, Arkansas Archeological Survey Research Series, Fayetteville, AR: Arkansas Archeological Survey.
Codding BF, Parker AK, Jones TL (2019) Territorial behavior among Western North American foragers: Allee effects, within group cooperation, and between group conflict. Quatern Int 518:31–40
Conolly J (2018) Hunter-gatherer mobility, territoriality, and placemaking in the Kawartha Lakes Region, Ontario. Canadian Journal of Archaeology, 42
Damlien H, Kjällquist M, Knutsson K (2018) The pioneer settlement of Scandinavia and its aftermath. New evidence from western and central Scandinavia. In: Knutsson K, Knutsson H, Apel J, Glørstad H (eds) The Technology of Early Settlement in Northern Europe - Transmission of Knowledge and Culture. Sheffield: Equinox
David É, Kjällquist M (2018) Transmission of knowledge, crafting and cultural traditions, interregional contact and interaction, 7300 Cal BC. In: Glørstad H, Knutsson K, Knutsson H, Apel J (eds) The technology of early settlement in Northern Europe - transmission of knowledge and culture. Equinox Publishing, Sheffield, pp 231–276
Drucker P (1939) Rank, wealth, and kinship in northwest coast society. Am Anthropol 41:55–65
Ekman J (1974) Djurbensmaterialet från stenålderslokalen Ire. Hangvar sn, Gotland. . In: Janzon G (ed) Gotlands mellanneolitiska gravar. Stockholm: Almqvist & Wiksell, pp 212–246
Elmendorf WW (1960) The structure of Twana culture with comparative notes on the structure of Yurok culture by AL Kroeber. Washington State University, Pullman
Elmendorf WW (1993) Twana narratives: native historical accounts of a Coast Salish culture. University of Washington Press, Seattle
Ericson JE (1985) Strontium isotope characterization in the study of prehistoric human ecology. J Hum Evol 14:503–514
Eriksson G (2003) Norm and difference: Stone Age dietary practice in the Baltic region, Stockholm: Jannes Snabbtryck Kuvertproffset HB.
Eriksson G (2004) Part-time farmers or hard-core sealers? Västerbjers studied by means of stable isotope analysis. J Anthropol Archaeol 23:135–162
Fraser M, Sanchez-Quinto F, Evans J, Storå J, Götherström A, Wallin P, Knutsson K, Jakobsson M (2018) New insights on cultural dualism and population structure in the Middle Neolithic Funnel Beaker culture on the island of Gotland. J Archaeol Sci Rep 17:325–334
Freeman J, Anderies JM (2015) The socioecology of hunter–gatherer territory size. J Anthropol Archaeol 39:110–123
Glykou A, Eriksson G, Storå J, Schmitt M, Kooijman E, Lidén K (2018) Intra‐and inter‐tooth variation in strontium isotope ratios from prehistoric seals by laser ablation (LA)‐MC‐ICP‐MS. Rapid Communications in Mass Spectrometry
Goldstein L (1981) One-dimensional archaeology and multi-dimensional people: spatial organization and mortuary analysis. In: Chapman R, Kinnes I, Randsborg K, C. 53 – 69. (ed.) The archaeology of death. Cambridge: Cambridge University Press, pp 53–69
Goodman AH, Armelagos GJ, Rose JC (1980) Enamel hypoplasias as indicators of stress in three prehistoric populations from Illinois. Human biology 515–528
Gron KJ, Montgomery J, Nielsen PO, Nowell GM, Peterkin JL, Sørensen L, Rowley-Conwy P (2016) Strontium isotope evidence of early Funnel Beaker Culture movement of cattle. J Archaeol Sci Rep 6:248–251
Grøn O (2015) Territorial infrastructure, markers and tension in Late Mesolithic hunter-gatherer societies: an ethnoarchaeological approach. In: Bicho N, Detry C, Price TD, Cunha E (eds) Muge 150th: The 150th anniversary of the discovery of mesolithic shell middens. Newcastle: Cambridge Scholars Publishing
Gubser NJ (1965) The Nunamiut Eskimos, hunters of Caribou. Yale University Press, New Haven
Gummesson S, Hallgren F, Kjellström A (2018) Keep your head high: skulls on stakes and cranial trauma in Mesolithic Sweden. Antiquity 92:74–90
Günther T, Malmström H, Svensson EM, Omrak A, Sánchez-Quinto F, Kılınç GM, Krzewińska M, Eriksson G, Fraser M, Edlund H, Munters AR, Coutinho A, Simões LG, Vicente M, Sjölander A, Jansen Sellevold B, Jørgensen R, Claes P, Shriver MD, Valdiosera C, Netea MG, Apel J, Lidén K, Skar B, Storå J, Götherström A, Jakobsson M (2018) Population genomics of Mesolithic Scandinavia: investigating early postglacial migration routes and high-latitude adaptation. PLoS Biol 16:e2003703
Haverkort CM, Weber A, Katzenberg MA, Goriunova OI, Simonetti A, Creaser RA (2008) Hunter-gatherer mobility strategies and resource use based on strontium isotope (87Sr/86Sr) analysis: a case study from Middle Holocene Lake Baikal, Siberia. J Archaeol Sci 35:1265–1280
Holt E, Evans JA, Madgwick R (2021) Strontium (87Sr/86Sr) mapping: a critical review of methods and approaches. Earth Sci Rev 216:103593
Hoppe KA, Koch PL, Furutani T (2003) Assessing the preservation of biogenic strontium in fossil bones and tooth enamel. Int J Osteoarchaeol 13:20–28
Horstwood M, Evans J, Montgomery J (2008) Determination of Sr isotopes in calcium phosphates using laser ablation inductively coupled plasma mass spectrometry and their application to archaeological tooth enamel. Geochim Cosmochim Acta 72:5659–5674
Janzon GO (1974) Gotlands mellanneolitiska gravar. Stockholm university
Jonsson, L. 1988. The vertebrate faunal remains from the Late Atlantic settlement Skateholm in Scania, South Sweden. In: Larsson L (ed) The Skateholm Project I: Man and Environment. LXXIX ed, Lund: Almqvist & Wiksell International, pp 56–88
Kelly R (2013) The lifeways of hunter-gatherers: the foraging spectrum. Cambridge University Press, Cambridge
Kjällquist M (2020) Kulturkontakter i Sydskandinavien under mesolitikum: Hantverkstraditioner, råmaterialval och mobilitet för 9000 år sedan, med utgångspunkt från Norje Sunnansund i Blekinge. Uppsala universitet, Uppsala
Kjällquist M, Boethius A, Emilsson A (2016) Norje Sunnansund. Boplatslämningar från tidigmesolitikum och järnålder. Särskild arkeologisk undersökning 2011 och arkeologisk förundersökning 2011 och 2012, Ysane socken, Sölvesborgs kommun i Blekinge län, Karlskrona: Blekinge museum
Kjällquist M, Price TD (2019) Mesolithic mobility and social contact networks in south Scandinavia around 7000 BCE: lithic raw materials and isotopic proveniencing of human remains from Norje Sunnansund, Sweden. J Anthropol Archaeol 53:186–201
Klassen L (ed) (2020) The Pitted Ware Culture on Djursland: supra-regional significance and contacts in the Middle Neolithic of southern Scandinavia. Aarhus University Press, Aarhus
Kootker LM, van Lanen RJ, Kars H, Davies GR (2016) Strontium isoscapes in the Netherlands. Spatial variations in 87Sr/86Sr as a proxy for palaeomobility. J Archaeol Sci Rep 6:1–13
Ladegaard-Pedersen P, Sabatini S, Frei R, Kristiansen K, Frei KM (2021) Testing Late Bronze Age mobility in southern Sweden in the light of a new multi-proxy strontium isotope baseline of Scania. PLoS ONE 16:e0250279
Lahtinen M, Arppe L, Nowell G (2020) Source of strontium in archaeological mobility studies—marine diet contribution to the isotopic composition. Archaeol Anthropol Sci 13:1
Larsson L (ed) (1988) The Skateholm Project I: man and environment, Lund: Regiae Societatis Humaniorum Litterarum Lundensis LXXIX
Larsson L (1989) Late Mesolithic settlements and cemeteries at Skateholm, southern Sweden. In: Bonsall C (ed) The Mesolithic in Europe, Edinburgh: John Donald, pp 367-378
Larsson L (1990) Dogs in Fraction – Symbols in Action. In: Vermeersch PM, Van Peer P (eds) Contribution to the Mesolithic in Europe. Papers presented at the forth international symposium `The Mesolithic in Europe, Leuven 1990. 5 ed, Leuven University Press: Leuven, pp 153-160
Larsson L (1993) The Skateholm Project: Late Mesolithic coastal settlement in southern Sweden. In: Bogucki P (ed) Case studies in European prehistory. CRC, Ann Arbor, pp 31–62
Larsson L (2016) Some aspects of mortuary practices at the Late Mesolithic cemeteries at Skateholm, southernmost Sweden. In: Grünberg J, Gramsch B, Larsson LJO, Meller H (eds) Mesolithic burials – rites, symbols and social organisation of early postglacial communities. International Conference Halle (Saale), Germany, 18th–21st September 2013. Tagungen des Landesmuseums für Vorgeschichte Halle Band 13/II, Landesamt für Denkmalpflege und Archäologie Sachsen-Anhalt. Halle: Landesmuseum für Vorgeschichte, pp 175-184
Larsson M, Magnell O, Styring A, Lagerås P, Evans J (2020) Movement of agricultural products in the Scandinavian Iron Age during the first millennium AD: 87Sr/86Sr values of archaeological crops and animals in southern Sweden. STAR: Sci Technol Archaeol Res 6:96–112
Layton R (1986) Political and territorial structures among hunter-gatherers. Man 21:18–33
Layton R, Rowley-Conwy P (2013) Wild things in the north? Hunter-gatherers and the tyranny of the colonial perspective. Anthropologie 51:213–230
Lazzerini N, Balter V, Coulon A, Tacail T, Marchina C, Lemoine M, Bayarkhuu N, Turbat T, Lepetz S, Zazzo A (2021) Monthly mobility inferred from isoscapes and laser ablation strontium isotope ratios in caprine tooth enamel. Sci Rep 11:2277
Lee RB (1984) The Dobe! Kung, New York: Holt, Rinehart and Winston
Lewis JP, Ryves DB, Rasmussen P, Olsen J, van der Sluis LG, Reimer PJ, Knudsen KL, McGowan S, Anderson NJ, Juggins S (2020) Marine resource abundance drove pre-agricultural population increase in Stone Age Scandinavia. Nat Commun 11:2006
Lundström V, Peters R, Riede F (2021) Demographic estimates from the Palaeolithic-Mesolithic boundary in Scandinavia: comparative benchmarks and novel insights. Philos Trans R Soc B: Biol Sci 376:20200037
Löfvendahl R, Åberg G, Hamilton PJ (1990) Strontium in rivers of the Baltic Basin. Aquat Sci 52:315–329
Marlowe F (2010) The Hadza: hunter-gatherers of Tanzania. Univ of California Press, Berkeley
Marlowe FW (2005) Hunter-gatherers and human evolution. Evol Anthropol 14:54–67
Maurer A-F, Galer SJG, Knipper C, Beierlein L, Nunn EV, Peters D, Tütken T, Alt KW, Schöne BR (2012) Bioavailable 87Sr/86Sr in different environmental samples — effects of anthropogenic contamination and implications for isoscapes in past migration studies. Sci Total Environ 433:216–229
McArthur J (2010) Strontium isotope stratigraphy. In: Ratcliffe KT, Zaitlin BA (eds) Application of modern stratigraphic techniques: theory and case histories, edited by: , SEPM Spec. Tulsa: SEPM (Society for Sedimentary Geology), pp 129-142
Montgomery J (2010) Passports from the past: Investigating human dispersals using strontium isotope analysis of tooth enamel. Ann Hum Biol 37:325–346
Montgomery J, Evans JA, Neighbour T (2003) Sr isotope evidence for population movement within the Hebridean Norse community of NW Scotland. J Geol Soc 160:649–653
Moritz M, Scaggs S, Shapiro C, Hinkelman S (2020) Comparative study of territoriality across forager societies. Hum Ecol 48:225–234
Nilsson Stutz L (2003) Embodied rituals and ritualized bodies: tracing ritual practices in Late Mesolithic burials: Lund University
Olson C (2008) Neolithic Fisheries: osteoarchaeology of fish remains in the Baltic Sea region. Stockholm University, Archaeology and Ancient Culture, Stockholm
Persson O, Persson E (1988) Anthropological report concerning the interred Mesolithic populations from Skateholm, Southern Sweden. Excavation season 1983–1984. In: Larsson L (ed) The Skateholm Project. I. Man and environment. Lund: Societatis Humaniorum Litterarum Lundensis LXXIX, pp 89-105
Price DT, Larsson L, Magnell O, Borić D (2021) Sedentary hunters, mobile farmers: The spread of agriculture into prehistoric Europe. In: Borić D, Antonović D, Mihailović B (eds) Foraging Assemblages. The Italian Academy for Advanced Studies in America, Columbia University, New York, pp 579–583
Price TD, Burton JH, Bentley RA (2002) The characterization of biologically available strontium isotope ratios for the study of prehistoric migration. Archaeometry 44:117–135
Price TD, Johnson CM, Ezzo JA, Burton JH, Ericson JA (1994) Residential mobility in the prehistoric southwest United States: a preliminary study using strontium isotope analysis. J Archaeol Sci 21:315–315
Reid DJ, Dean MC (2006) Variation in modern human enamel formation times. J Hum Evol 50:329–346
Rowley-Conwy P (1998) Cemeteries, seasonality and complexity in the Ertebølle of southern Scandinavia. In: Zvelebil M, Dennell R, Domanska L (eds) Harvesting the sea, farming the forest: the emergence of Neolithic societies in the Baltic region. Sheffield Academic Press, Sheffield, pp 193–202
Rowley-Conwy P, Piper S (2016) Hunter-gatherer variability: developing the models for the northern coasts. Arctic 69:1–14
Saxe AA (1970) Social dimensions of mortuary practices. University of Michigan
Sjögren K-G, Price TD, Ahlström T (2009) Megaliths and mobility in south-western Sweden. Investigating relationships between a local society and its neighbours using strontium isotopes. J Anthropol Archaeol 28:85–101
Snoeck C, Ryan S, Pouncett J, Pellegrini M, Claeys P, Wainwright AN, Mattielli N, Lee-Thorp JA, Schulting RJ (2020) Towards a biologically available strontium isotope baseline for Ireland. Sci Total Environ 712:136248
Steadman DW (2008) Warfare related trauma at Orendorf, a middle Mississippian site in west-central Illinois. Am J Phys Anthropol 136:51–64. https://doi.org/10.1002/ajpa.20778
Stenberger M (1939) Das Västerbjersfeld - Ein Gräberfeld der Ganggräberzeit auf Gotland. Acta Archaeologica X:60–105
Stenberger M, Dahr E, Munthe H (1943) Das Grabfeld von Västerbjers auf Gotland, Stockholm: Kungl. Vitterhets Historie och Antikvitets Akademien
Storå J (2001) Reading bones: Stone Age hunters and seals in the Baltic. Department of Archaeology, Stockholm University, Stockholm
Thirlwall M (1991) Long-term reproducibility of multicollector Sr and Nd isotope ratio analysis. Chemical Geology: Isotope Geoscience Section 94:85–104
van der Merwe NJ, Lee-Thorp JA, Thackeray JF, Hall-Martin A, Kruger FJ, Coetzee H, Bell RHV, Lindeque M (1990) Source-area determination of elephant ivory by isotopic analysis. Nature 346:744–746
Wilhelmson H, Ahlström T (2015) Iron Age migration on the island of Öland: apportionment of strontium by means of Bayesian mixing analysis. J Archaeol Sci 64:30–45
Willmes M, Kinsley L, Moncel M-H, Armstrong R, Aubert M, Eggins S, Grün R (2016) Improvement of laser ablation in situ micro-analysis to identify diagenetic alteration and measure strontium isotope ratios in fossil human teeth. J Archaeol Sci 70:102–116
Acknowledgements
We are grateful for the financial support from The Swedish Research Council, VR-2019-02975, Birgit och Gad Rausings Stiftelse för Humanistisk forskning, Gyllenstiernska Krapperupsstiftelse, Helge Ax:son Johnssons stiftelse, Ebbe Kocks stiftelse, Lars Hiertas stiftelse, Sahlénsstiftelse and Längmanska stiftelsen. We acknowledge and thank the curators at the Historical Museum at Lund University (Skateholm), the Blekinge Museum (Norje Sunnansund) and the National Historical Museums of Sweden (Västerbjers) for the permission to analyse the teeth in their care. We also wish to thank the reviewers and our editor for the insightful comments. This is Vegacenter publication #055.
Funding
Open access funding provided by Lund University. We are grateful for the financial support from the following:
Adam Boethius: The Swedish Research Council, VR-2019–02,975, https://www.vr.se/ and Birgit och Gad Rausings Stiftelse för Humanistisk forskning, https://www.rausing.org/
Lars Larsson: Gyllenstiernska Krapperupsstiftelse, Helge Ax:son Johnssons stiftelse, Ebbe Kocks stiftelse, Lars Hiertas stiftelse, Sahlénsstiftelse och Längmanska stiftelsen.
The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
Adam Boethius: Conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, project administration, resources, writing—original draft. Review and editing, Skateholm and Norje Sunnansund sampling.
Mathilda Kjällquist: Data curation, baseline sampling, Norje Sunnansund sampling, writing background on Norje Sunnansund. Review and editing.
Torbjörn Ahlström: Data curation, baseline sampling, Västerbjers sampling, writing background on Västerbjers. Review and editing.
Melanie Kielman-Schmitt: Data curation—LA‐MC-ICP‐MS operation—editing.
Lars Larsson: Data curation, writing background on Skateholm. Review and editing. Funding acquisition. Skateholm sampling.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Boethius, A., Kjällquist, M., Kielman-Schmitt, M. et al. Diachronic forager mobility: untangling the Stone Age movement patterns at the sites Norje Sunnansund, Skateholm and Västerbjers through strontium isotope ratio analysis by laser ablation. Archaeol Anthropol Sci 14, 176 (2022). https://doi.org/10.1007/s12520-022-01640-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-022-01640-0