
Abstract
Introduction
Targeting of the proinflammatory cytokine interleukin 17A (IL-17A) or tumor necrosis factor alpha (TNFα) with the monoclonal antibodies (mAbs) ixekizumab or adalimumab, respectively, is a successful therapy for chronic plaque psoriasis. The effects of these treatments on immune cell populations in the skin are largely unknown.
Methods
In this study, we compared the composition of cutaneous, lesional and non-lesional immune cells and blood immune cells in ixekizumab- or adalimumab-treated patients with psoriasis.
Results
Our data reveal that both treatments efficiently downregulate T cells, macrophages and different subsets of dendritic cells (DCs) in lesional skin towards levels of healthy skin. In contrast to lesional skin, non-lesional areas in patients harbor only few or no detectable DCs compared to the skin of healthy subjects. Treatment with neither ixekizumab nor adalimumab reversed this DC imbalance in non-lesional skin of psoriatic patients.
Conclusion
Our study shows that anti-IL-17A and anti-TNFα therapy rebalances the immune cell repertoire of lesional skin in psoriatic patients but fails to restore the disturbed immune cell repertoire in non-lesional skin.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Why carry out this study? |
Chronic plaque psoriasis is a common inflammatory skin disease. Monoclonal antibody therapy is the most effective treatment in moderate-to-severe cases |
Our study compares the impact of two prominent biologicals (ixekizumab and adalimumab) on the immune cell occurrence in the skin and blood |
What was learned from the study? |
Anti-IL-17A and anti-TNFα both regulate the increase in immune cells in diseased skin but are not capable of establishing a sound immune cell profile in uninvolved skin of psoriatic patients, as in healthy persons |
The striking difference in the profile of DCs, especially the low frequency in non-lesional psoriatic skin, suggests that DCs could be involved in the localized onset and distribution of this systemic disease |
Only ixekizumab was able to downmodulate the higher numbers of CD15+CD14− cells in the blood of patients suffering from psoriasis and might therefore play a role in the differentiation of granulocytic myeloid-derived suppressor cells (GMDSCs) |
Introduction
Chronic plaque psoriasis is an inflammatory disease with characteristic skin manifestations and affects 2–3% of individuals in Western countries. The disease burden is considerable, and the risk for comorbidities, including cardiometabolic or mental health disorders, is increased within the psoriatic population but might be reduced by effective treatment [1,2,3]. Psoriasis is understood to be primarily a DC/T cell-driven disease [4,5,6,7]. The prevailing opinion is that epidermal keratinocytes hyperproliferate in response to the massive production of IL-17A by pathogenic CD4+ T-helper 17 cells (Th17) [8,9,10,11]. Th17 expansion in turn is induced by IL-23, which is released by activated myeloid DCs [12, 13]. However, IL-17A is also released by other cells in psoriatic skin, including polymorphonuclear neutrophils (PMNs) and mast cells [14,15,16,17]. The role of these cells in the pathophysiology of psoriasis is poorly defined. TNFα is released by several immune cells, including macrophages/monocytes, natural killer cells (NKs) and DCs [18,19,20]. In psoriasis, elevated TNFα stimulates DCs to produce IL-23 and additionally upregulates the proinflammatory scenery for keratinocytes in synergy with IL-17A [5, 21, 22]. While the knowledge of different immune cell types, particularly T cells and DCs, is profound in lesional skin, less is known about non-lesional skin or the peripheral blood in chronic plaque psoriasis patients [23,24,25,26,27,28,29]. In the study presented here, we analyzed and compared the blockade of IL-17A with TNFα inhibition on the immune cell profile in the skin and blood of psoriasis patients
Methods
Study Design
This was an experimental ex vivo human study conducted at the Department of Dermatology and the Institute of Immunology of the Medical University of Vienna (Vienna, Austria). We investigated the impact of ixekizumab (IXE) and adalimumab (ADA) treatment on skin and blood samples from psoriatic patients. The study was approved by the ethics committee of the Medical University of Vienna (ECS 1642/2017) in accordance with the Helsinki Declaration of 1964 and its later amendments and followed national law and ethical principles in research. It required voluntary participation of patients and healthy controls with written informed consent. Study participation involved the collection of necessary personal data, photography of involved skin areas and skin and blood samples. All patients gave written informed consent to the release of clinical images captured. The individual risk for participants was low, and adverse effects were not observed in the course of the study.
Recruitment of Subjects
Psoriasis patients from the outpatient clinic of the Department of Dermatology, Medical University of Vienna, as well as healthy controls were voluntarily enrolled in the study. All study participants were \(\ge\) 18 and < 80 years of age. All included patients had stable (> 6 months) moderate-to-severe chronic plaque psoriasis involving ≥ 10% of total body surface area (BSA), with a psoriasis area and severity index (PASI) score of ≥ 10 and a Physician Global Assessment (PGA) of ≥ 3 before treatment initiation. Biopsies and blood samples of subjects on therapy with either IXE or ADA were realized within 12 weeks after treatment initiation. Further details on the inclusion and exclusion criteria are listed in the supplementary material.
Collection of Samples and Staining Strategy
Clinical pictures of participants with psoriasis were made at the timepoint of inclusion (Figure S5). In each participant, blood and skin samples were collected. Peripheral blood mononuclear cells (PBMCs) were isolated from blood samples. Six-millimeter (mm) punch biopsies were obtained in duplicate from lesional and non-lesional skin of psoriatic patients, whereas one 6-mm punch biopsy of healthy skin was obtained from controls. Biopsies were split into paraffin and frozen sections. Paraffin-embedded samples were used for histology (Figure S6), immunohistochemistry and archiving, whereas frozen samples were used for immunofluorescence (IF) staining. Based on a set of mAbs, multiplex immunofluorescence (mIF) coexpression analyses were established on acetone-fixed cryosections of lesional psoriatic, non-lesional psoriatic and healthy skin samples, according to a mIF panel, particularly elaborated for this study (Table S2, S3; Figure S7, S8). A detailed description of sample collection and immunohistochemical and immunofluorescence staining protocols are disclosed in the supplementary material section.
Analysis of Skin Samples via TissueFAXS and TissueQuest
Stained skin tissue sections were acquired using the TissueFAXS imaging system (TissueGnostics GmbH, Vienna, Austria) on an Axio Observer Z1 microscope equipped with an LD Plan-Neofluar 20 × /0.4 objective (Zeiss). Data from the acquired photos were processed with TissueQuest image analysis software 6.0 and StrataQuest (TissueGnostics GmbH). Matched isotype controls were included for analysis of background staining. Biopsy specimens were read in a blinded fashion by an independent investigator who was not involved in the collection or staining of the tissue sections. The epidermis and dermis were analyzed separately. In each biopsy, an area of 5 mm2 of the upper dermis and the epidermis (at a length of 5 mm basement membrane) was tagged. Labeled cells within the tagged area were expressed as the number of cells/mm2. Additionally, the percentage and the absolute numbers of labeled cells within the selected area were also gathered. Only cells clearly positive for the antigen were counted. Artifacts, blood vessels and adnexa were excluded from the areas of interest.
Flow Cytometry
Flow cytometric analysis of single-cell suspensions was performed by surface marker staining. PBMCs (2 × 105) were incubated with conjugated mAbs (Table S2) for 30 min at 4 °C. After washing with PBS-BSA, the cells were analyzed by flow cytometry (LSRFortessa™ Cell Analyzer, Becton Dickinson, Franklin Lakes, NJ).
Statistical Analysis
Flow cytometric data were analyzed using FlowJo data analysis software (Becton Dickinson). Further statistical analysis for flow cytometric data and TissueQuest-generated output was performed via GraphPad Prism version 7.00 (GraphPad Software, La Jolla, CA). Therefore, the normality of the data distribution was assessed by applying the Shapiro-Wilk test. For a normal distribution, parametric one- or two-way analysis of variance (ANOVA) was performed, depending on the number of tested conditions. This was followed by Tukey’s pairwise comparisons (Tukey’s post hoc test) with adjusted P values. If a Gaussian distribution was not assumed, the nonparametric Kruskal-Wallis test was applied, followed by Dunn’s correction for multiple comparisons. Probability values of P < 0.05 were considered statistically significant, represented as *P < 0.05, **P < 0.01, ***P < 0.001 and ****P < 0.0001. The results are depicted as the mean ± standard deviation (SD) in scatter plots or bar charts.
Results
Study Population
We enrolled 21 subjects in this study divided into 4 subgroups: 5 healthy controls, 6 untreated psoriasis patients, 5 psoriatic patients under IL-17A inhibition with IXE and 5 individuals under TNFα blockade with ADA within the first 12 weeks of therapy. All groups showed comparable demographic characteristics (Table S1). All participants receiving mAb therapy responded to treatment with at least a PASI75% reduction within the first 6 months. The results presented in Fig. 1 demonstrate a mean PASI reduction of 94% and 87% among IXE- and ADA-treated subjects, respectively, indicating that all patients initially responded to treatment, excluding nonresponders. Further details on the study population, including respective body sites for biopsies, additional scores for disease activity (BSA, PGA), quality of life [Dermatology Life Quality Index (DLQI)] and the histopathologic psoriasis severity score (HPSS) at inclusion, are reported in the supplementary material.

Dynamics of the psoriasis area and severity index (PASI) upon treatment with IL-17A or TNFα inhibitors. Bar charts are depicted as the mean ± standard deviation (SD) with respective single values plotted as points. Psoriatic patients were grouped into subpopulations based on their treatment regimen. PASI scores are shown at study inclusion (I) for all subpopulations. PASI scores under anti-IL-17A (IXE) or anti-TNFα (ADA) therapy are given before treatment initiation (B), at inclusion (I) and within 6 months (R) to confirm treatment response. The percent decrease in the baseline PASI at the timepoint of inclusion and within 6 months are presented for both treatment regimens separately. C Healthy control; PSO psoriasis without treatment; IXE psoriasis under IL-17A inhibition with ixekizumab; ADA psoriasis under TNFα inhibition with adalimumab; ns nonsignificant
Treatment of Psoriasis Patients with Ixekizumab or Adalimumab Reduces T Cells and NK Cells in the Skin
The results of our study demonstrate that CD4+ and CD8+ T cells were found at a higher density in lesional areas of the epidermal and dermal regions of the skin of untreated psoriasis patients than in healthy donors (Fig. 2). Treatment with IXE and ADA reduced the number of CD4+ T cells (Fig. 2a, b) and CD8+ T cells (Fig. 2d, e) in lesional skin to levels observed in healthy donors. An unexpected finding was that the amounts of CD3+Ki67+ T cells in the epidermis of patients treated with ADA were increased (Fig. S2a). The number and percentage of T cells in the peripheral blood of psoriasis patients were not altered compared to those in healthy donors and were not significantly modulated upon treatment of patients with IXE or ADA (Fig. 2c, f). However, T cells isolated from the blood of psoriasis patients expressed higher levels of CD71 (Fig. S2c). The expression of T cell activation marker was significantly lower in patients treated with IXE or ADA.

Impact of ixekizumab and adalimumab treatment on T cells in psoriatic skin and blood. Epidermal and dermal CD3+ T cells, including their combination with CD4+ (a, b) and CD8+ (d, e) cells, are given in numbers/mm2 as bar charts with the mean ± standard deviation (SD) and respective single values plotted as points. The corresponding CD3+ populations in the blood are equally depicted in % of PBMC fraction (c, f). Psoriatic patients were grouped based on their treatment regimen during the induction phase. C Healthy control; PSO psoriasis without treatment; IXE psoriasis under IL-17A inhibition with ixekizumab; ADA psoriasis under TNFα inhibition with adalimumab; ns nonsignificant; L lesional; N non-lesional
Furthermore, untreated psoriatic plaques appeared with more epidermal CD56+ NK cells than healthy skin (Fig. S3a). In the healthy controls, only a few NK cells were present in the epidermis. An increase in epidermal NK cells was not detected under treatment with IXE and ADA (Fig. S3a). Interestingly, the number of dermal NK cells was only reduced by IXE but not upon ADA therapy (Fig. S3b). No relevant changes in CD19+ B cell counts were observed in the skin or blood within the different patient cohorts (Fig. S3d, e, f).
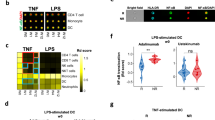
Impact of Treatment with Adalimumab or Ixekizumab on Low-Density CD15+CD14− Cells
Granulocytes (CD15+CD14−) were rare in lesional and non-lesional areas of the dermis in psoriatic patients, and the density and frequency of these cells were not regulated by IXE and ADA treatment (Fig. 3b). In contrast, the percentage of granulocytes was significantly increased in the lesional epidermis of psoriatic patients (Fig. 3a) but not in the non-lesional epidermis (data not shown). Treatment with IXE or ADA reduced granulocytes in the epidermis to baseline levels of healthy controls (Fig. 3a). Analysis of the PBMCs by flow cytometry revealed an increased number of low-density CD15+CD14− cells, potentially resembling granulocytic myeloid-derived suppressor cells (GMDSCs), in patients with psoriasis compared to healthy controls. Interestingly, CD15+CD14− cells were downregulated in IXE- but not ADA-treated patients within the induction phase of therapy (Fig. 3c, d). Thus, ADA and IXE differ in their capacity to downregulate CD15+CD14− cells in psoriasis patients.

Changes in the granulocyte cell system upon IL-17A and TNFα inhibition. Expression of CD15+ granulocytes in lesional skin (a, b), and the PBMC fraction (c, d) of psoriatic patients was analyzed. Data are depicted in either cells/mm2 (skin) or % (PBMCs) as bar charts with the mean ± standard deviation (SD). Single values are plotted as points. Psoriatic patients were grouped into subpopulations based on their treatment regimen during the induction phase. A representative flow cytometry plot of CD15+CD14− cells in a healthy control, an untreated psoriasis patient and a patient under anti-IL-17A (IXE) or anti-TNFα (ADA) therapy is shown under e. Percentages of CD15+CD14− cells are displayed on the right. C Healthy control; PSO psoriasis without treatment; IXE psoriasis under IL-17A inhibition with ixekizumab; ADA psoriasis under TNFα inhibition with adalimumab; ns nonsignificant
Treatment of Psoriasis Patients with Ixekizumab or Adalimumab Does Not Restore the Repertoire of DCs in the Skin
Langerhans cells (LCs) are prototypic CD1a+ cells in the epidermis. The density of LCs in lesional but not in non-lesional areas of psoriatic skin was found to be significantly decreased (Fig. 4a, b). Neither treatment with IXE nor treatment with ADA changed LC numbers in the psoriatic epidermis. In contrast to epidermal LCs, the density of CD1a+ cells was strongly increased in the dermis of lesional skin (Fig. 4c). CD1a+ cells, which could resemble inflammatory DCs, were absent in the dermis of healthy controls as well as in the non-lesional dermis of psoriasis patients (Fig. 4d). A high number of CD1a+ cells in the lesional dermis was not detected in patients upon IXE or ADA treatment (Fig. 4c). Elevated percentages of CD1a+ cells were also detected in the blood of psoriasis patients, which were normalized to levels in healthy controls upon both treatment regimens (Fig. 4e). Other characteristic markers of DCs that were analyzed in this study were CD141, which is expressed on conventional DC-type 1 (cDC1), and CD303, a marker for plasmacytoid DCs (pDCs) (Fig. 4) [30,31,32,33]. The density of CD141+ and CD303+ cells was significantly decreased in non-lesional zones of the dermis of psoriasis patients compared to healthy skin (Fig. 4i, n). This cellular profile was not restored in patients upon treatment with IXE or ADA. The results presented in Figure S4 demonstrate that in contrast to DCs, the levels of CD68+ cells, which are most likely macrophages, in non-lesional skin were similar to those in healthy controls.

Dynamics of dendritic cells in lesional versus non-lesional skin and blood in response to anti-IL-17A or anti-TNFα therapy. Bar charts are depicted as the mean ± standard deviation (SD), including lesional and non-lesional epidermal and dermal CD1a+ (a–d), CD141+ (f–i) and CD303+ (k–n) cells in numbers/mm2. Corresponding CD1a+ (e), CD141+ (j) and CD303+ (o) cells of the PBMC fraction are given in %. Single values are plotted as points. Psoriatic patients were grouped into subpopulations based on their treatment regimen during the induction phase. C Healthy control; PSO psoriasis without treatment; IXE psoriasis under IL-17A inhibition with ixekizumab; ADA psoriasis under TNFα inhibition with adalimumab; ns nonsignificant
The lesional skin of psoriasis patients harbored significantly more CD141+ and CD303+ cells than that of healthy controls (Fig. 4h, m). Here, treatment with IXE or ADA reversed the numbers of both cell types to levels of healthy controls (Fig. 4h, m). In the blood, the numbers of CD303+ cells were increased in psoriasis patients, and expression was absent upon IXE but not adalimumab therapy (Fig. 4o). The expression of CD141+ cells in the blood was similar in psoriatic patients and healthy donors (Fig. 4j). Taken together, IXE and ADA had a comparable impact on the DC compartment of psoriatic patients but did not restore the repertoire of DCs in non-lesional skin or LC in the epidermis.
Discussion
The biologicals IXE directed against IL-17A and ADA against TNFα have significantly improved the handling of moderate to severe psoriasis [34]. In this study, we performed a head-to-head analysis of immune cell distribution and frequency in lesional and non-lesional skin and blood of IXE- and ADA-treated patients with psoriasis. Our data show that both treatment strategies have similar effectiveness in reducing the numbers of key drivers of the disease—T cells, DCs and pDCs—in lesional areas of psoriatic skin. In addition, IXE- but not ADA-treated psoriatic patients exhibited downregulated NK cells in lesional regions of the skin and low-density CD15+CD14− cells in the blood. The effective downregulation of immune cells in diseased skin, demonstrated with anti-IL-17A and anti-TNFα in this study, raises the question of whether treatment with these monoclonal antibodies can decrease inflammatory damage in other tissues apart from the skin. A protective scenario in pre-clinical enthesitis was already shown for patients with psoriasis under adalimumab [35]. Studies on anti-IL-17A agents, also report promising results for patients with psoriatic arthritis [36, 37]. Future studies on the immune cell profile under treatment in tissues involved in psoriasis-associated comorbidities are crucial to identify if the effects are similar to the skin. To be able to decrease tissue damage or even prevent psoriatic comorbidities such as psychiatric, cardiovascular or arthritic involvement would justify early systemic treatment and minimize disease burden.
Psoriasis is typically accompanied by an increase in different types of DCs, in particular CD1a+, cDC1 and pDCs, in the lesional dermis [21, 38,39,40,41,42]. Interestingly, we discovered that the number of DCs in the non-lesional skin was significantly lower, even compared to healthy donors (Fig. 4). The striking difference in the profile of DCs in psoriatic skin suggests that DCs could be involved in the localized onset and distribution of this systemic disease. The auto-induction of psoriatic lesions in xenografts of unaffected psoriatic skin of immune-deficient mice supposed a regulatory assignment of resident immune cells and local immune environments [43]. In this model, pDCs were rapidly recruited to the xenograft and played an important role during the initiation phase of the psoriatic plaque formation [21] by producing type I interferon and promoting functional maturation of myeloid DCs in psoriasis [44, 45]. Thus, the depletion of pDCs in non-lesional psoriatic skin demonstrated in our results might be required to keep the respective skin unaffected in a patient with psoriasis. However, the consequences of the low frequency of DCs in non-lesional psoriatic skin need to be studied in more detail in future studies. The results of our study further demonstrate that both ADA and IXE treatment regimens did not restore the low amounts of DCs in the dermis or LCs in the epidermis in non-lesional or lesional areas of psoriatic skin, respectively. Thus, both biologicals are effective in regulating the increase in immune cells in diseased skin but are seemingly not capable of establishing a sound immune cell profile in uninvolved skin of psoriatic patients as in healthy persons.
The number of CD68+ cells was reduced because of IXE or ADA treatment, suggesting that the increase in macrophages is controlled by both biologics. In contrast to DCs, the number of CD68+ cells in non-lesional skin was comparable to that in the skin of healthy individuals and did not point to a disturbed immune balance in the unaffected skin of psoriasis patients.
Another unexpected finding was the elevated number of low-density CD15+CD14− cells in the PBMC fraction of psoriasis patients, which could resemble GMDSCs. GMDSCs, also described as polymorphonuclear-myeloid-derived suppressor cells (PMN-MDSCs), are a relatively new subset of regulatory myeloid-derived cells of the phagocytic cell system that are generated under inflammatory conditions and inhibit effector T cells [46,47,48,49]. GMDSCs have been detected in various diseases, above all in cancer but also in sepsis or autoimmune disease [50,51,52,53,54,55,56]. Although myeloid suppressor cells in general have shown a remarkable ability to suppress T cell responses in cancer, their role seems to be more heterogenous in autoimmune diseases such as psoriasis [29, 57]. It has been reported that GMDSCs are elevated in the blood of psoriatic patients [58], which could be interpreted as a compensatory mechanism in chronic inflammatory diseases. However, IXE but not ADA was able to downmodulate the higher numbers of CD15+CD14− cells in patients suffering from psoriasis. The fact that GMDSCs in vitro promoted Th17 differentiation matches the observation that anti-IL-17A but not anti-TNFα downmodulated the higher numbers of these cells in the blood of our patients [59]. Thus, IL-17A but not TNFα is seemingly a key factor for the differentiation of GMDSCs in the human immune system. It remains to be determined in future studies whether the CD15+CD14− cells that we found in the PBMCs of our patients are indeed GMDSCs [50, 60, 61]. If so, the presence or absence of GMDSCs before and after IXE therapy could be used as a biomarker to monitor disease progression and therapeutic success.
The mechanisms underlying the treatment of patients with ADA and IXE have been studied and discussed in many studies [62,63,64,65,66]. We observed that neither of the two biologicals induced enhanced apoptosis in immune cells in either the skin or the blood. However, we detected reduced expression of activation markers on T cells, suggesting that inflammatory signals stemming from inflamed tissue may promote T cell activation and probably also T cell proliferation, which is supported by the finding of increased levels of CD71 on T cells in psoriatic patients and the inhibition of CD71 by IXE and ADA.
A limitation of our study was the relatively small study collective of 21 subjects. Biopsies are not a standard procedure at the Medical University of Vienna in the monitoring of psoriasis patients. So, the number of patients included in the study was intended to be small. Additionally, participants were enrolled according to their current treatment status and were not prospectively followed up. Nevertheless, data between patients were generally consistent, arguing against major interpatient variability. Novel methods such as single-cell RNA sequencing (scRNAseq) have significantly advanced our knowledge in the field and led to the description of additional cell types [67,68,69,70,71,72,73]. The findings of expanded T/NK cells in lesional psoriatic skin corroborate recent transciptome studies, which deployed scRNAseq to resolve the cellular heterogeneity of inflammatory skin and confirmed a Tc17/Th17 signature in psoriasis [72,73,74,75]. The increased numbers of dendritic cells in lesional psoriatic skin in our study are a confirmatory finding and in accordance with flow or transcriptome analyses [69, 72]. The low frequency of DCs in non-lesional psoriatic skin demonstrated by our results are unexpected and in a way contrary to previous reports [71]. Follow-up single-cell analyses might focus on non-lesional psoriatic skin compared to healthy and lesional psoriatic skin to further elucidate the gene programs involved in the localized onset of the disease. In summary, the key aspects of the immune cell profile in psoriasis correspond between multiplex immunohistochemistry/immunofluorescence techniques and transcriptome analysis, including scRNAseq. The purpose of our study was, however, not to better characterize immune cells and their subsets in psoriatic skin but to compare the impact of two prominent biologicals (IXE, ADA) on the immune cell occurrence.
Conclusion
Taken together, the results of our study demonstrate that ADA and IXE are both effective in downregulating the increase in prominent cellular drivers of psoriasis but fail to revert the DC deficiencies in non-lesional parts of the skin.
Data Availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author (Julia Tittes) on reasonable request.
References
Rapp SR, et al. Psoriasis causes as much disability as other major medical diseases. J Am Acad Dermatol. 1999;41(3 Pt 1):401–7.
Gelfand JM, et al. Risk of myocardial infarction in patients with psoriasis. JAMA. 2006;296(14):1735–41.
Korman NJ. Management of psoriasis as a systemic disease: what is the evidence? Br J Dermatol. 2020;182(4):840–8.
Gottlieb SL, et al. Response of psoriasis to a lymphocyte-selective toxin (DAB389IL-2) suggests a primary immune, but not keratinocyte, pathogenic basis. Nat Med. 1995;1(5):442–7.
Hawkes JE, Chan TC, Krueger JG. Psoriasis pathogenesis and the development of novel targeted immune therapies. J Allergy Clin Immunol. 2017;140(3):645–53.
Lowes MA, et al. Increase in TNF-alpha and inducible nitric oxide synthase-expressing dendritic cells in psoriasis and reduction with efalizumab (anti-CD11a). Proc Natl Acad Sci U S A. 2005;102(52):19057–62.
Lowes MA, Suárez-Fariñas M, Krueger JG. Immunology of psoriasis. Annu Rev Immunol. 2014;32:227–55.
Harrington LE, et al. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat Immunol. 2005;6(11):1123–32.
Park H, et al. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nat Immunol. 2005;6(11):1133–41.
Teunissen MB, et al. Interleukin-17 and interferon-gamma synergize in the enhancement of proinflammatory cytokine production by human keratinocytes. J Invest Dermatol. 1998;111(4):645–9.
Krueger JG, et al. IL-17A is essential for cell activation and inflammatory gene circuits in subjects with psoriasis. J Allergy Clin Immunol. 2012;130(1):145-54.e9.
Lee E, et al. Increased expression of interleukin 23 p19 and p40 in lesional skin of patients with psoriasis vulgaris. J Exp Med. 2004;199(1):125–30.
Zaba LC, et al. Psoriasis is characterized by accumulation of immunostimulatory and Th1/Th17 cell-polarizing myeloid dendritic cells. J Invest Dermatol. 2009;129(1):79–88.
Lin AM, et al. Mast cells and neutrophils release IL-17 through extracellular trap formation in psoriasis. J Immunol. 2011;187(1):490–500.
Keijsers RR, et al. Cellular sources of IL-17 in psoriasis: a paradigm shift? Exp Dermatol. 2014;23(11):799–803.
Reich K, et al. Evidence that a neutrophil-keratinocyte crosstalk is an early target of IL-17A inhibition in psoriasis. Exp Dermatol. 2015;24(7):529–35.
Res PC, et al. Overrepresentation of IL-17A and IL-22 producing CD8 T cells in lesional skin suggests their involvement in the pathogenesis of psoriasis. PLoS ONE. 2010;5(11): e14108.
Haranaka K, et al. Purification, characterization, and antitumor activity of nonrecombinant mouse tumor necrosis factor. Proc Natl Acad Sci U S A. 1986;83(11):3949–53.
Schottelius AJ, et al. Biology of tumor necrosis factor-alpha- implications for psoriasis. Exp Dermatol. 2004;13(4):193–222.
Bazzoni F, Beutler B. The tumor necrosis factor ligand and receptor families. N Engl J Med. 1996;334(26):1717–25.
Nestle FO, et al. Plasmacytoid predendritic cells initiate psoriasis through interferon-alpha production. J Exp Med. 2005;202(1):135–43.
Uyemura K, et al. The cytokine network in lesional and lesion-free psoriatic skin is characterized by a T-helper type 1 cell-mediated response. J Invest Dermatol. 1993;101(5):701–5.
Keijsers RR, et al. Balance of Treg vs. T-helper cells in the transition from symptomless to lesional psoriatic skin. Br J Dermatol. 2013;168(6):1294–302.
Mehta H, et al. Differential changes in inflammatory mononuclear phagocyte and T-cell profiles within psoriatic skin during treatment with Guselkumab vs. Secukinumab. J Invest Dermatol. 2021;141(7):1707-1718.e9.
Brunner PM, et al. The atopic dermatitis blood signature is characterized by increases in inflammatory and cardiovascular risk proteins. Sci Rep. 2017;7(1):8707.
Kolbinger F, et al. β-Defensin 2 is a responsive biomarker of IL-17A-driven skin pathology in patients with psoriasis. J Allergy Clin Immunol. 2017;139(3):923-932.e8.
Chiricozzi A, et al. Increased expression of interleukin-17 pathway genes in nonlesional skin of moderate-to-severe psoriasis vulgaris. Br J Dermatol. 2016;174(1):136–45.
Wang CQF, et al. IL-17 induces inflammation-associated gene products in blood monocytes, and treatment with ixekizumab reduces their expression in psoriasis patient blood. J Invest Dermatol. 2014;134(12):2990–3.
Soler DC, et al. Increased, but functionally impaired, CD14(+) HLA-DR(-/low) myeloid-derived suppressor cells in psoriasis: a mechanism of dysregulated T cells. J Invest Dermatol. 2016;136(4):798–808.
Kvedaraite E, Ginhoux F. Human dendritic cells in cancer. Sci Immunol. 2022;7(70):eabm9409.
Scheib N, et al. The dendritic cell dilemma in the skin: between tolerance and immunity. Front Immunol. 2022;13: 929000.
Kamata M, Tada Y. Dendritic cells and macrophages in the pathogenesis of psoriasis. Front Immunol. 2022;13: 941071.
Cabeza-Cabrerizo M, et al. Dendritic cells revisited. Annu Rev Immunol. 2021;39:131–66.
Armstrong AW, et al. Comparison of biologics and oral treatments for plaque psoriasis: a meta-analysis. JAMA Dermatol. 2020;156(3):258–69.
Narcisi A, et al. Effects of TNF-α inhibition on pre-clinical enthesitis: observational study on 49 psoriatic patients. J Dermatol Treat. 2022;33(3):1703–6.
Simopoulou T, et al. Secukinumab, ixekizumab, bimekizumab and brodalumab for psoriasis and psoriatic arthritis. Drugs Today (Barc). 2023;59(3):135–67.
Lynde C, et al. Comparative effectiveness of biologics across subgroups of patients with moderate-to-severe plaque psoriasis: results at week 12 from the PSoHO study in a real-world setting. Adv Ther. 2023;40(3):869–86.
Glitzner E, et al. Specific roles for dendritic cell subsets during initiation and progression of psoriasis. EMBO Mol Med. 2014;6(10):1312–27.
Kim TG, Kim SH, Lee MG. The origin of skin dendritic cell network and its role in psoriasis. Int J Mol Sci. 2017;19(1):42.
Zaba LC, et al. Amelioration of epidermal hyperplasia by TNF inhibition is associated with reduced Th17 responses. J Exp Med. 2007;204(13):3183–94.
Chamian F, et al. Alefacept reduces infiltrating T cells, activated dendritic cells, and inflammatory genes in psoriasis vulgaris. Proc Natl Acad Sci U S A. 2005;102(6):2075–80.
Wang A, Bai Y. Dendritic cells: the driver of psoriasis. J Dermatol. 2020;47(2):104–13.
Boyman O, et al. Spontaneous development of psoriasis in a new animal model shows an essential role for resident T cells and tumor necrosis factor-alpha. J Exp Med. 2004;199(5):731–6.
Lande R, et al. Plasmacytoid dendritic cells sense self-DNA coupled with antimicrobial peptide. Nature. 2007;449(7162):564–9.
Farkas A, Tonel G, Nestle FO. Interferon-alpha and viral triggers promote functional maturation of human monocyte-derived dendritic cells. Br J Dermatol. 2008;158(5):921–9.
Dumitru CA, et al. Neutrophils and granulocytic myeloid-derived suppressor cells: immunophenotyping, cell biology and clinical relevance in human oncology. Cancer Immunol Immunother. 2012;61(8):1155–67.
Veglia F, Perego M, Gabrilovich D. Myeloid-derived suppressor cells coming of age. Nat Immunol. 2018;19(2):108–19.
Bergenfelz C, Leandersson K. The generation and identity of human myeloid-derived suppressor cells. Front Oncol. 2020;10:109.
Schmielau J, Finn OJ. Activated granulocytes and granulocyte-derived hydrogen peroxide are the underlying mechanism of suppression of t-cell function in advanced cancer patients. Cancer Res. 2001;61(12):4756–60.
Perez C, et al. Immunogenomic identification and characterization of granulocytic myeloid-derived suppressor cells in multiple myeloma. Blood. 2020;136(2):199–209.
Argyropoulos KV, et al. Tumor-infiltrating and circulating granulocytic myeloid-derived suppressor cells correlate with disease activity and adverse clinical outcomes in mycosis fungoides. Clin Transl Oncol. 2020;22(7):1059–66.
Loeuillard E, et al. Targeting tumor-associated macrophages and granulocytic myeloid-derived suppressor cells augments PD-1 blockade in cholangiocarcinoma. J Clin Invest. 2020;130(10):5380–96.
Janols H, et al. A high frequency of MDSCs in sepsis patients, with the granulocytic subtype dominating in gram-positive cases. J Leukoc Biol. 2014;96(5):685–93.
Rodriguez PC, et al. Arginase I-producing myeloid-derived suppressor cells in renal cell carcinoma are a subpopulation of activated granulocytes. Cancer Res. 2009;69(4):1553–60.
Denny MF, et al. A distinct subset of proinflammatory neutrophils isolated from patients with systemic lupus erythematosus induces vascular damage and synthesizes type I IFNs. J Immunol. 2010;184(6):3284–97.
Tucker SL, Sarr D, Rada B. Granulocytic myeloid-derived suppressor cells in cystic fibrosis. Front Immunol. 2021;12: 745326.
Chen C, et al. Targeting myeloid-derived suppressor cells is a novel strategy for anti-psoriasis therapy. Mediators Inflamm. 2020;2020:8567320.
Ilkovitch D, Ferris LK. Myeloid-derived suppressor cells are elevated in patients with psoriasis and produce various molecules. Mol Med Rep. 2016;14(4):3935–40.
Ferrer G, et al. Myeloid-derived suppressor cell subtypes differentially influence T-cell function, T-helper subset differentiation, and clinical course in CLL. Leukemia. 2021;35(11):3163–75.
Pember SO, et al. Density heterogeneity of neutrophilic polymorphonuclear leukocytes: gradient fractionation and relationship to chemotactic stimulation. Blood. 1983;61(6):1105–15.
Fridlender ZG, et al. Transcriptomic analysis comparing tumor-associated neutrophils with granulocytic myeloid-derived suppressor cells and normal neutrophils. PLoS ONE. 2012;7(2): e31524.
Chan JR, et al. IL-23 stimulates epidermal hyperplasia via TNF and IL-20R2-dependent mechanisms with implications for psoriasis pathogenesis. J Exp Med. 2006;203(12):2577–87.
Chiricozzi A, et al. Scanning the immunopathogenesis of psoriasis. Int J Mol Sci. 2018;19(1):179.
Green L, et al. Clinical and molecular effects of interleukin-17 pathway blockade in psoriasis. J Drugs Dermatol. 2020;19(2):138–43.
Hendriks AG, et al. The effect of adalimumab on key drivers in the pathogenesis of psoriasis. Br J Dermatol. 2014;170(3):571–80.
Tokuyama M, Mabuchi T. New treatment addressing the pathogenesis of psoriasis. Int J Mol Sci. 2020;21(20):7488.
Cytlak U, et al. Differential IRF8 transcription factor requirement defines two pathways of dendritic cell development in humans. Immunity. 2020;53(2):353-370.e8.
Fuentes-Duculan J, et al. A subpopulation of CD163-positive macrophages is classically activated in psoriasis. J Invest Dermatol. 2010;130(10):2412–22.
Kim J, et al. Single-cell transcriptomics applied to emigrating cells from psoriasis elucidate pathogenic versus regulatory immune cell subsets. J Allergy Clin Immunol. 2021;148(5):1281–92.
McGovern N, et al. Human dermal CD14+ cells are a transient population of monocyte-derived macrophages. Immunity. 2014;41(3):465–77.
Nakamizo S, et al. Single-cell analysis of human skin identifies CD14+ type 3 dendritic cells co-producing IL1B and IL23A in psoriasis. J Exp Med. 2021. https://doi.org/10.1084/jem.20202345.
Reynolds G, et al. Developmental cell programs are co-opted in inflammatory skin disease. Science. 2021. https://doi.org/10.1126/science.aba6500.
Zhang B, et al. Single-cell profiles reveal distinctive immune response in atopic dermatitis in contrast to psoriasis. Allergy. 2023;78(2):439–53.
Xia D, et al. Applications of single-cell RNA sequencing in atopic dermatitis and psoriasis. Front Immunol. 2022;13:1038744.
Liu J, et al. Single-cell RNA sequencing of psoriatic skin identifies pathogenic Tc17 cell subsets and reveals distinctions between CD8(+) T cells in autoimmunity and cancer. J Allergy Clin Immunol. 2021;147(6):2370–80.
Acknowledgements
The authors thank Elisabeth Riedl and Christopher Schuster for their valuable scientific input. Additionally, we thank Claus Wenhardt and Johanna Strobl for their expert technical assistance. We thank all study participants for their involvement in the study.
Medical Writing and Editorial Assistance.
We received editorial assistance from an AI program of Research Square during the writing of this article for spelling correction. This was funded by the corresponding author (Julia Tittes).
Funding
This work was conducted as an investigator-initiated trial and supported by funds from Eli Lilly (project no. I1F-OE-O001). The Rapid Service Fee was funded in part by Eli Lilly, the Institute of Immunology, and the Department of Dermatology, Medical University of Vienna.
Author information
Authors and Affiliations
Contributions
Conceptualization: Julia Tittes, Johannes Stöckl; Data curation: Julia Tittes, Jennifer Brell, Pia Fritz, Sarojinidevi Künig; Formal Analysis: Julia Tittes, Pia Fritz; Funding acquisition: Julia Tittes, Johannes Stöckl; Investigation: Julia Tittes, Jennifer Brell, Sarojinidevi Künig; Methodology: Julia Tittes, Jennifer Brell, Pia Fritz, Georg Stary, Julia M. Ressler, Johannes Stöckl; Project administration: Julia Tittes, Johannes Stöckl; Resources: Julia Tittes, Constanze Jonak, Georg Stary, Johannes Stöckl; Software: Julia Tittes, Jennifer Brell, Pia Fritz, Julia M. Ressler; Supervision: Julia Tittes, Johannes Stöckl; Validation: Julia Tittes, Constanze Jonak, Georg Stary, Julia M. Ressler, Johannes Stöckl; Visualization: Julia Tittes, Pia Fritz, Johannes Stöckl; Writing—original draft: Julia Tittes, Johannes Stöckl; Writing—review & editing: Julia Tittes, Constanze Jonak, Georg Stary, Julia M. Ressler, Wolfgang Weninger, Johannes Stöckl.
Corresponding author
Ethics declarations
Conflict of Interest
Other than that mentioned above, Julia Tittes reports consulting fees from Lilly, speaker honoraria from Lilly and Novartis, all outside the submitted work, as well as travel support from Almirall, AbbVie and Bristol-Myers Squibb through institution. Julia M. Ressler received speaker honoraria from Bristol-Myers Squibb, Roche, Amgen and Novartis and travel support by Sanofi, Roche and Bristol-Myers Squibb through institution. All other authors declare no conflicts of interest (COI). The corresponding author, Julia Tittes, has changed her affiliation after the completion of the manuscript. Her new affiliation is the Institute for Clinical Pathology and Molecular Pathology at the University Clinic St.Pölten-Lilienfeld (Karl Landsteiner University).
Ethical Approval
The study was approved in accordance with the Helsinki Declaration of 1964 and its later amendments by the ethics committee of the Medical University of Vienna (ECS 1642/2017) on the 25th of September 2017 and followed national law and ethical principles in research. It required voluntary participation of patients and healthy controls with written informed consent. Additionally, all patients gave written informed consent to the release of clinical images captured.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License, which permits any non-commercial use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc/4.0/.
About this article

Cite this article
Tittes, J., Brell, J., Fritz, P. et al. Regulation of the Immune Cell Repertoire in Psoriasis Patients Upon Blockade of IL-17A or TNFα. Dermatol Ther (Heidelb) 14, 613–626 (2024). https://doi.org/10.1007/s13555-024-01112-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13555-024-01112-4