
Abstract
Upper Miocene sedimentary deposits of Sierra de las Cabras in the Prebetic Ranges (Jumilla, Región de Murcia, Spain) yield a exceptional fossil site of diverse vertebrate ichnofauna, including mammals and birds. This paper presents a detailed study of those ichnites of that site which correspond to mammals. The footprints are located in stratigraphic surfaces found in a thin marly limestone succession that can be assigned to the latest Tortonian to earliest Messinian. The mammalian record consists of 324 footprints (included in trackways, pairs, groups and isolated ichnites) of diverse vertebrates, including: Hippipeda, Rhynoceripeda, Canipeda, Felipeda, Ursipeda, Rodentipeda, as well possible Suipeda. The ichnites were imprinted on wet and cohesive marly carbonate mud, deposited in a semi-arid wetland with relevant freshwater influx. The fossil site, cleaned and protected some years ago but herein studied by the first time, add together the nearby Hoya de la Sima site (Hippipeda, Pecoripeda, Bestiopeda and Paracamelichnium, and probably proboscidean ichnites) to conform an exceptional record of Late Miocene ichnofauna in southern Iberia.
Resumen
Los depósitos sedimentarios del Mioceno superior de la Sierra de las Cabras en la Zona Prebética (Jumilla, Región de Murcia, España) aportan un yacimiento excepcional con diversa icnofauna que incluye mamíferos y aves. Este artículo presenta un estudio detallado de las huellas que corresponden a los mamíferos. Las huellas están en superficies estratigráficas en una sucesión de calizas margosas delgadas que se pueden asignar a la parte terminal del Tortoniense o la basal del Mesiniense. Las pisadas de mamíferos consisten en 324 huellas (incluidas en rastrilladas, pares, grupos e icnitas aisladas) de varios vertebrados que incluyen: Hippipeda, Rhynoceripeda, Canipeda, Felipeda, Ursipeda, Rodentipeda y probablemente Suipeda. Las huellas estan impresas en barro calizo margoso compacto depositado en un ambiente húmedo semiárido con influjo de agua dulce. El yacimiento, limpiado y protegido hace algunos años pero estudiado ahora por primera vez, junto al yacimiento vecino de la Hoya de la Sima (Hippipeda, Pecoripeda, Bestiopeda, Paracamelichnium y probablemente huellas de poroiboscídeos) constituyen un conjunto excepcional de icnofauna del Mioceno en el Sureste de la Península Ibérica.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
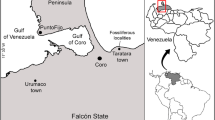
In this paper we study the mammal ichnites found in the Upper Miocene of Sierra de las Cabras near the town of Jumilla (Murcia province, SE Spain). The ichnofossil site, first discovered in 2007 and later cleaned and protected (Herrero, C. 2010; Herrero, E. 2010) by members of the Jeronimo Molina Museum (Jumilla), has yielded over 324 mammal footprints that correspond to a wide range of ichnotaxa, among them: Hippipeda, Rhynoceripeda, Canipeda, Felipeda, Ursipeda, Rodentipeda, as well as possibly Suipeda. The Sierra de las Cabras tracksite joins the well-known Hoya de la Sima site (e.g., Herrero, 1997; Pérez-Lorente et al., 1997, 1999, 2009; Vilas et al., 2006), located about 9 km to the northeast, to form an exceptional record of Late Miocene vertebrate ichnites in the southeast of the Iberian Peninsula (Fig. 1). The Hoya de la Sima site provided diverse vertebrate tracks, inside the protected enclosure (undetermined pecoriped, camelids [Paracamelichnum], Hippipeda); and others outside but on the same levels (? Canipeda /? Felipeda, Ursipeda and possible elephant tracks).

Location of Paleogene and Neogene vertebrate ichnological sites of peninsular Spain. Geological and geographical situation of Sierra de las Cabras tracksite
The site is a succsssion of three stratigraphic beds with diverse types of footprints at their surfaces (numbered 1–3 from base to top), which are however not distributed homogeneously over all of them (Fig. 2). Only the ichnotaxon Felipeda appears in the three layers, but Rodentipeda is only in one of them (in the upper layer or surface 3). Many tracks are deformed because of the physical state of the mud at the time of formation, which can be different depending on the trackway. This means that most of the footprints are not merely stamps (faithful reproduction of the base of the foot), but more complex structures with features that allow us to analyze the dynamics of the movement of the autopods during the phases of the footprint formation. The two Suipeda tracks have been found outside the closure at levels that do not correspond with the three above mentioned.

Schematic plan of the Sierra de las Cabras site (mesh is 60 × 60 cm), showing the distribution of footprints (upper part of the figure) and the location of the three stratigraphic surfaces containing the footprints (lower part)
The precise number of footprints is difficult to estimate because many counted footprints consist of partially or totally overlapping pes-manus pairs, which make it difficult to decide whether they should be counted as one or as two prints. We have counted 324 mammalian footprints (plus 51 avian ones; not considered in this paper) which add 375 prints to those previously described in the near site of Hoya de la Sima (360) making a total of more than 730 footprints in stratigraphic levels of approximately similar age.
Other sites of Paleogene and Neogene vertebrate ichnites in the Iberian Peninsula that will be mentioned throughout the text are located in Fig. 1 and summarized in Suppl. Table 1.
2 Geological framework and age of Sierra de las Cabras tracksite
The Sierra de las Cabras tracksite (CBR) is located eight kilometers westwards from the town of Jumilla in the southern hillside of Sierra de las Cabras (Lat. 38º 28′ 53″, Lon. 1º 24′ 52″, datum: ETRS89; Fig. 1), a WSW-ENE elongated mountain range that mostly consists of folded and thrusted Jurassic carbonates and, to a lesser extent, Middle to Upper Miocene sedimentary rocks. That sierra belongs to the Prebetic Zone, the outermost part of the External Zones of the Betic Cordillera, and resulted from tectonic contraction that mainly occurred during Middle to Late Miocene (e.g., Roca et al., 2006; Rubinat et al., 2013), in the late orogenic stage of the Betic orogen.
The footprint beds are stratigraphically located in a ~ 10 m thick unconformity bounded interval, that consists of thinly bedded, fine grained limestones and marls. In this paper, we assign to this unit a latest Tortonian to earliest Messinian age, based on the following criteria:
-
(1)
The unit uncorformably overlies Serravallian marine deposits (Baena, 1981) that consist of heterozoan bioclastic calcarenites with variable siliciclastic content, with abundant coralline algae, bryozoans, and bivalves. That underlying marine sequence is about 140 m thick in Sierra de las Cabras and presents strong tectonic deformation, showing sub-vertical dip in the site area, contrasting with the gentler dip of the footprint bearing unit.
-
(2)
The unit presents in the fossil site variable SE dip, decreasing upwards and southwards from 60º to 20º, indicating significant syn-sedimentary tectonism, that could be framed in the post-orogenic stage of the Betic Alpine orogenic evolution (e.g., Jabaloy Sánchez et al., 2019; Vera, 2000). That stage developed when rapidly rising new reliefs, and rapidly subsiding basins installed unconformably onto older tectonostratigraphic terranes. During this stage the tectonic frame was quite active rather than calm, and dominated by transtensional tectonics, vertical displacements, diapiric activity of Triassic salts, and volcanism.
-
(3)
A thin volcano-sedimentary bed containing relevant amounts of volcanic ash and other pyroclastic material has been recognized in the stratigraphic levels immediately above the ichnite beds. This bed is very probably related to the K-rich lamproitic volcanism occurring in the nearby area of La Celia (Fúster et al., 1967; Bellon et al., 1977), which has been radiometrically dated as earliest Messinian (6.8 ± 0.4 Ma; Turner et al., 1999; 6.76 ± 0.04 Ma, Duggen et al., 2005); and interpreted as related to transtensive tectonic processes occurring in a late-orogenic framework (Duggen et al., 2005).(
-
(4)
The unit is overlaid by post-orogenic yellowish marls of Pliocene age that cover large areas of the foothills of Sierra de las Cabras (Baena, 1981).
During the late Miocene, the area of Jumilla experienced drastic paleogeographic changes determined by the tectonic closure of the so-called Betic Strait, a marine seaway that connected the Atlantic and the Mediterranean up to the early Tortonian (Martín et al., 2009). With this large-scale regional process, the shallow marine systems with extensive carbonate sedimentation that had prevailed during early Langhian to early Tortonian times gave way to a complex landscape of intramountain basins with dominant continental settings where the marine influence was progressively but rapidly lost (Rossi et al., 2015). The sedimentary deposits bearing the Sierra de las Cabras footprints were deposited in that changing geographic framework.
3 Sedimentary framework and paleoenvironment
The stratigraphic interval that bears the ichnite fossil site consists of cm-scale bedded fine-grained carbonates, marls and marly limestones. There is also some evidence of evaporite minerals, but limited to pseudomorphs after idiomorphic euhedral gypsum crystals, included in carbonate and marly facies. Partial early dolomitization of some of the calcareous facies has been also observed. The ichnites are preserved in the marly limestone beds. Most of these deposits show a typical chalky texture and whitish color.
The main sedimentary facies found in those levels allow us to interpret the paleoenvironment of the Sierra de las Cabras ichnofossil site as a semi-arid wetland system, characterized by shallow ponds, small swamps and marshes, that provided the habitat for a diverse fauna, as revealed by the footprints. Freshwater was probably provided by springs that could formed at the foot of the carbonate hills. Those springs would drain water accumulated during rainy periods in the porous carbonates that form the rocks of the sierra. Sedimentation in those wetlands was dominated by carbonate deposits. These include a variety of freshwater, palustrine and pedogenic carbonates.
Palustrine deposits are mostly represented by micritic facies that contain abundant ostracod shells and small gastropods (Hydrobia sp., usually less than 3 mm high). The presence of calcified green algae and cyanobacteria is also remarkable, forming small bioherms dominated by calcareous tubes possibly attributable to the genus Cladophorites, but a variety of microbial forms can be recognized. Planar to ondulate stromatolite structures are also common in these deposits.
These deposits exhibit extensive evidence of subaerial exposure and pedogenic modification. They include a variety of features such as micro-nodulization, circumgranular-cracking, clotted fabric, pedogenic-ooids, root voids, and Microcodium. These features, and the absence of well-developed pedogenic crusts, are indicative of dominantly “wet” environments in swamp—marsh settings. Pedogenic transformation of original facies ranges from incipient to very intense. Although those habitats were probably wet during most of the time, significant fluctuations in water level should occur, causing episodic emersion. These episodes are evidenced by desiccation cracks and brecciation of semi-lithified sediment.
Water chemistry probably experienced significant changes due to variations in the hydric balance, i.e. changes in freshwater influx vs. evaporation. Negative balance associated to intense evaporative processes eventually determined precipitation of sulphate minerals within the sediment, as revealed by gypsum pseudomorphs present in some levels. Partial dolomitization observed in some of those levels suggests early diagenetic transformation of calcium carbonate by Mg enriched waters.
4 Ichnology
4.1 Method and nomenclature
The footprints are found on the top of carbonate levels. These surfaces are herein named surface 1, surface 2 and surface 3, from base to top. The ichnites were imprinted on highly cohesive, partially consolidated, muddy sediment consisting of marly carbonate mud, that was embedded in water at the time the animals left their tracks. Those places could be flooded, with the sedimentary surface below several centimeters of water, or recently exposed but still saturated with water, as revealed by the presence of desiccation cracks, which are particularly abundant in surface 2.
Besides the vertebrate ichnites, traces of small invertebrates are also present in those surfaces, including horizontal locomotion and grazing traces, probably produced by annelids and gastropods. Microbial mats are present on all those surfaces, and could have notably increased the cohesivity, adherence and viscosity (plastic behavior) of the sedimentary material. Each of the three ichnite surfaces is overlaid by a thin (few centimeters thick) bed of greenish gray marls, that had favored ichnite preservation.
A 60 × 60 cm square grid was outlined with chalk on the studied stratigraphic surfaces with footprints in the CBR site (Fig. 2). The longer lines of the grid are horizontal and coincide with the strike of the stratification planes, the other lines being perpendicular and coincident to the dip. On this mesh an alphanumeric reference system was established, and photographs were taken over all squares, with the aid of a 60 × 60 cm frame with a 5 × 5 cm fine rope mesh. This grid was used as a reference base to locate every feature considered of interest in this study, including ichnites and stratigraphic and structural features.
The images were processed with Adobe Photoshop® and translated on a scaled plane using AutoCAD. The dimension and orientation data are measured directly on the outcrop and on the AutoCAD representation. For 3D processing of the images Adobe Photoshop®, Photosynt®, SynthExport®, MeshLab® and Paraview® software were used.
In this paper, a trackway is considered as a sequence of three or more tracks of the same animal left by locomotion (Thulborn, 1990). Measurements of the morphometric features of tracks and trackways, and the nomenclature system are adopted from classic works (Casamiquela, 1964; Leonardi, 1987; Casanovas et al., 1989). Abbreviations (Table 1) and concepts are based on Haubold (1971), Leonardi (1987) and Pérez-Lorente (2001). In the description of ichnites and surface features, terms from Thulborn (1990), Allen (1997), Fornos et al. (2002), Gatesy (2003) and Requeta et al. (2006–2007) are used. The structures of the ichnites are associated to the T (touch), W (weight) and K (kick-off) phases from Thulborn and Wade (1989).
ª
The reference given to a footprint consists of the site acronym (CBR), followed by the trackway number, a dot, and the footprint number of those included in that trackway (e.g., CBR15.1). For quadrupedal footprints, a lower case letter or subscript (“p” or “m”) is added to the reference for distinguishing pes and manus marks respectively (CBR15.1p is the pes print and CBF15.1 m the manus of the first ichnite pair of the 15 trackway of CBR site). If the two prints are of the same pair, superimposed or separated, the letter “e” (CBR15.1e) is added. The nomenclature of the mammalian prints begins at CBR15 because references CBR1 to CBR14 had been already assigned to avian ichnites from this same site. Those avian ichnites of Sierra de las Cabras site are the focus of the second part of this paper.
We differentiate between “overprint” (footprint printed in whole or in part on another), “superposition” [number of paces taken with the foot before reaching the print of the hand in the same cycle (Peabody, 1959)], and “overlap” (pes ichnite placed in front of the manus print of the same cycle). It should be also noted that the terms “wall” and “hole” are herein used in the ichnological sense (Allen, 1997) [some papers dedicated to the autopodia of present-day horses use “wall” to refer the sector of the hoof between the outer and inner boundaries, from coronary band to the surface resting on the ground].
In this paper the footprint length (l) is measured on the longitudinal axis of the ichnite (pes and manus print lengths are designated by the abbreviation “lp” and “lm” respectively); the print width (a) is measured perpendicular to the longitudinal axis and measures the separation between two lines that are parallel to the axis and tangential to the footprint; the digit axis is defined by a median line that runs through the digit separating the print in two approximately similar halves; the digit length is measured along its axis and goes from the most distal end (including the nail mark) to the corresponding hypex; the reference point of each print (necessary to measure the pace, the stride, …) is the center of the footprint. We study the footprints as marks on the ground (parataxonomy); the trackmaker is a consequence, that is, our measurements are direct, not assumed. In the present data the digit mark is considered from the hypex to the tip of the nail and the outline is taken at the top of the footprint wall, at the point where the wall of the footprint intersects the tracking surface (cf. Pérez-Lorente, 2001, 2015).
4.2 Perissodactyla. Hippipeda Vialov, 1966
118 tracks were reported from Sierra de las Cabras site (Fig. 3). These correspond to 8 trackways, formed by sequences of ichnites each of which has the mark of the manus and that of the overprinted pes (CBR16, CBR22, CBR24, CBR31, CBR32, CBR33, CBR34 and CBR46); 5 groups of pairs (CBR17, CBR25, CBR28, CBR49 and CBR59), each conisting of two manus-pes print associations (two manus and two pes); and 40 isolated prints (up to CBR66 ichnite), each formed by a manus-pes association. Figure 4 illustrates the typical manus pes overprint in these ichnites, as well as the dimensions measured in each ichnite.

Location of Hippipeda tracks and trackways in the Sierra de las Cabras site

Hippipeda from Sierra de las Cabras site. Manus-pes group CBR16.2 used as an example to illustrate methodology. The left image shows the topography of the ichnite, whereas the right drawing shows the print outline, with the manus print (lower, green) and pes print (upper, blue). Measurements of the ichnite: am, width of the manus print; ap, pes print width; lp, pes print length; le, length of the entire ichnite; p^m, angle between the axis of the manus and pes prints
4.2.1 Characterization of the Hippipeda of Sierra de las Cabras
Most of the dual manus-pes prints show the back trace wider, more incomplete and more deformed that the front trace, and therefore the one that was first printed, i.e., the manus print (Suppl. Table 2). The front print (pes), narrower and probably also shorter, overprints and deforms the rear prints (manus).
Each of the tracks (manus or pes) is U-shaped, or V-shaped with the "branches" open at the back (Fig. 5), and with the deepest at the front. In many tracks, two rear marks are also distinguished as two shallow holes (Figs. 4, 5). The main, largest and central mark is attributed to the impression of the middle digit (III) and that of the two posterior holes to two much smaller digits (II and IV). It can be said that there is only one functional digit (III).

Outline of the ichnites. The top and bottom of the print wall are drawn. Green manus lines; blue pes lines: A CBR24.3. U shape of the pes print. Left front wall of the foot curved inwards. The marks of the back digits are distinguished in the manus print. B CBR25.2. V-shaped with the rounded apex of pes print. Collapse of the walls of the manus print. C and D Possible group of separate manus and pes prints of the same animal. E CBR22.6. Total overprint of the pes on the manus print. Scale: 5 cm. F CBR24.2. Sedimentary level fragment displaced by the foot backwards during the “kick-off” phase (Phase K)
The tip of the tracks (of digit III) is round, with the "branches" relatively straight, which open backwards. In some (Fig. 5A, B), the curvature of the hoof “branches” is concave lateral (left front wall of the image). The lateral holes, slightly oval and less deep, are placed next to the “branches”. In some tracks, there is a channel that runs along the inner front edge and the sides (Fig. 5C–E). This channel is interpreted as the hoof base mark. The width of the channel is variable due in part to the fact that the walls of the ichnites have fallen into the hollow. The walls of the hollows are vertical, and we cannot deduce the spatial shape of the hoof, which could be curved (semi-ellipsoidal, semi-oval or hemispherical truncated at the back (Sarjeant & Reynolds, 1999)), nor make estimates on the thickness of the hoof in relation to the size of the track because the channel is always deformed. From the above it can be deduced that they are traces of a quadruped perissodactyl animal whose autopodia have a main digit (III) and two secondary lateral digits (II and IV).
The ichnites appear in three different ways, defined by the position of the pes mark relative to the manus mark.
-
(i)
the pes overlaps the manus: we found only one case (CBR38) in which the pes exceeds the manus print (Fig. 5C, D), that is, the pes is placed in front of the manus print and separated from it.
-
(ii)
the pes overprints the manus: marks in which the mark of the manus is not distinguishable because it is completely overprinted by the pes (Fig. 5E).
-
(iii)
the pes partially covers the manus: these are the most common case, in which the pes print partially conceals the manus print (Fig. 5A, B). In some cases (Fig. 5F) the pes breaks and displaces some sediment of the tracking surface on the manus print.
The length and width of the prints vary considerably. We distinguish two associations (Suppl. Table 2): association A, of the CBR15 to CBR53 ichnites in which the largest dimension of the autopodial mark does not exceed 8 cm in length (Suppl. Table 3); and association B in which we include the ichnites CBR54 to CBR63, larger than 12 cm, the largest being of the order of 14 cm (trackway CBR60).
The pes marks of association A show the following dimensions (Fig. 6: Suppl. Tables 2, 3;): a (print width) from 1.4 to 7.8 cm; l (print length) from 2.4 to 7.8 cm; the average values are: a = 4.4 cm; l = 5.1 cm. The length values of the manus prints were not considered because they are incomplete. The width of manus prints ranges between 4.2 and 10.5 cm, with an average of 6.2 cm, clearly wider than those of the pes.

A Diagram showing the width to length (a/l) relationship of Hippipeda prints of pes. In trackways and pairs, the mean value has been taken. Footprints of the association A (less than 8 cm long) are plotted in black color, and the red ones correspond to those of association B. Dimensions given in centimeters, B evolution of the length of footprints of Equoidea through time, according to Reynolds (2006) but completed with more recent data. The red line shows the average footprint length ot the CBR Hippipeda.The thick line, which goes through the center of the vertical lines, clearly shows a tendency to increase. It should be noted that the anomalously high value for the Lower Miocene (l = 170 mm) has not been considered in the analysis because the measurement very possibly corresponds to a deformed footprint (Astibia et al., 2007)
Some authors (e.g., Reynolds, 2006) have pointed out that the size of animals increases during phylogeny, so their ichnites should also do. When the dimensions of ichnites described in literature are plotted against time, this trend is clearly outlined, as can be observed in Fig. 6 and Suppl. Table 4. The ichnites of Sierra de las Cabras clearly fit within this general trend (Fig. 6).
The ichnites of association B are more irregular, both in the outline and in the size and distribution of individuals in the site. It is possible that some of these tracks correspond to different trackmakers.
The bottom of the tracks (Fig. 7) is commonly flat and has no signs of the frog, or of any flexible pad (in the sense of Sarjeant & Reynolds, 1999), despite the mud probably being soft enough in some sectors of the site to generate some imprint. This absence of structures does not however necessarily imply a flat autopodium, as a relatively elastic behavior of the sediment surface could also impede its formation. That behavior could be favored by the presence of an algal mat cover. It is therefore possible that although the sole of the pes had structures, the tracking surface did not adapt to them due to its elasticity and, consequently, we cannot know if the base of the autopodium was flat or showed some type of shape.

Hoof marks in several footprints. A CBR15; B CBR22.8; C CBR26; D CBR30; E CBR36; F CBR38m. In some of the footprints there are marks of the lateral digits, and structures of collapse of the walls. In B there is part of a bird footprint at the lower end of the image, the horse footprint is posterior to it. In C, collapse structure of the walls, and front channel due to the dragging of the hoof in K-phase
4.2.2 Overprinting and other structures
As has already been said, practically all the ichnites show the two different parts corresponding to the fore and hind autopodium, so the complexity of the structures is greater than in the single autopodium ichnites. In order of formation (phases from Thulborn & Wade, 1989) and considering their origin, the observed structures are the following:
-
(i)
In phase T (touch-down: from the beginning of contact with the ground until the moment of total support) ramps and entry striae are formed due to the forward movement of the base of the autopods. Structures produced by manus are patent in many ichnites. The structures related to the entry of the pes are scarce because this autopodium begins to sink into the front wall of the manus print, that is, the pes begins to descend through the hollow of the manus print.
-
(ii)
In phase W (weight-bearing: maximum pressure phase), some lateral rims are formed by extrusion of mud (Fig. 8B, C). Mud extrusion rims are not evident in all traces because the displaced mud is not of the level of the tracking surface on an elastic sedimentary level, but of a lower level—large but relatively low extrusion zones are formed. In areas where the level is plastic or where the elastic level is broken, the pes penetrates the mud and produces the subsequent collapse of the walls. Probably in this phase, digit III leaves a deeper mark ahead than behind, so that the deepest part corresponds to the distal end of the digit and the shallowest one to the marks of the back digits. It is possible that, at the beginning of phase K, at least in some ichnites the depth of the front part of digit III is accentuated.
-
(iii)
In Phase K (kick-off) two types of structures are distinguished. On elastic ground there are striae, grooves of pes exit, and broken fragments of the tracking level shifted backwards (Fig. 5F). There are also marks of the forward movement of the upper part of the autopodium (Figs. 7C, 8A) and exit scars (Fig. 8). The formation of the scar (Gatesy et al., 1999; Pérez-Lorente and Herrero-Gascón, 2007) occurs because the autopodium penetrates so much in the mud that the forward movement of the lower limb (probably middle phalanx-metapodium sector) is produced inside the mud. This movement leaves a long channel between the anterior and posterior part of the ichnite. The long channel can be partially or totally collapsed, in which case it appears as a narrow channel or as a closed scar between two wider areas (entry and exit of the autopodium). When the channel is completely collapsed (it is a line), the front part of the ichnite is formed by the scar or eyelet of the pes. The exit structures of the autopodium that has been submerged in the mud can clearly be seen in the pes prints. Certain manus-pes groups of small and deep prints have an elongated front part, which we interpret due to the movement of the manus (?) forward during phase K, probably due to the low travel speed of the trackmaker.
-
(iv)
After the exit of the pes or even during the same exit, in some tracks, the walls fall towards the interior of the hollow leaving several types of structures such as the bulging of the walls (Figs. 5B, 7C), the narrowing and collapse of the hollow (Fig. 8A–C), or the breakage and staggering of the walls.

A and B Structures caused by penetration in the mud, corresponding to CBR46.1 and CBR46.3. C penetration structure and passage through the mud (CBR49). 1, pes exit channel; 2, pes exit walls; 3, advance and exit hole of pes; 4, pes print; 5, pes entry depression; 6, mud displaced by the pes; 7 manus entry depression; 8, manus print; 9, back digits (II, IV) of the manus; 10, entry structure of the manus; 11, entry drag marks of the manus; 12, eyelet shape of the narrow cleft structure; 13, narrow cleft mark of upper part of autopodium as it passes and later collapse of the walls; 14, pes print; 15, foot entry structure; 16 ramp of manus entrance
4.2.3 Trackways
The characters of the tracks (pes that is in front of the manus print) are normal in trackways of animals with relatively long limbs compared to the glenoacetabular distance (Pérez-Lorente et al., 2009) and in long stride animals (Kienapfel et al., 2014).
It is not possible to find out what types of gait the trackmakers used in the site due to the similarity in the position of the ichnites in the three trackways. The trackways are narrow to very narrow gauge (Ar / a = 0.46, Pérez-Lorente, 2001) in which Lr < 2a (the trackway width is less than the sum of the width of two tracks, except in CBR46) and the pes print is superimposed on, or overlaps (exceeds) that of the manus. These types of trackways are typical of slow walk and slow trot horses (Kienapfel et al., 2014).
Nor do we recognize left or right tracks due to: (a) the similarity of the soles of the horses (in current horses the four autopods are practically the same); (b) the very short length of the trackways, and their irregularity.
In the same way we have investigated—without results and for the same reasons as in the previous case—the angle p^m to verify if the orientation angle of the manus is greater or less than that of the pes. The placement of the angle p^m does not help in this site to discriminate left and right pairs.
It is very possible that the animals moved slowly, because there are no sliding marks associated with the action on the ground of the faster moving autopods, and because by the depth of the tracks it is deduced that the tracking surface was on soft ground, probably flexible and fragile (fragments of soil broken and displaced by hooves).
Having analyzed the direction of displacement, there is a maximum in the direction of NNE-SSW, with a predominant direction to the North (Fig. 9) This order is consistent with the gregarious behavior of these animals. We cannot know if, in the case of being a pack, it was a large or small group of individuals, although the density of tracks is that printed by a relatively small group. It is also possible that it was made up of individuals of various ages, due to the variation in size of the ichnites.

Diagram showing trajectories of displacement deduced from isolated tracks and trackways orientations. Most frequent trajectories are SSE—NNW with dominant direction towards the North
4.2.4 Ichnotaxonomic determination
The ichnogenus Hippipeda Vialov, 1966 is defined by “Footprints of functionally single-digit hooved herbivores, having a hemiellipsoidal to hemispherical or hemiovoidal hoof wall, with or without traces of a frog. One or two lateral digits (II and IV) may also be impressed in trackways during a fast gait, but these are much smaller than the central impression of digit III. Trackway narrow; stride and pace long” (Sarjeant & Reynolds, 1999).
The CBR15 to CBR63 tracks are the expression of authopod prints such as those included in the ichnogenus Hippipeda. The tracks of Sierra de las Cabras have:
-
(i)
Hippipeda characters: A single functional digit (III), mark of the hoof with a U-shaped or V-shaped, rounded tip and with lateral digit prints (II and IV), smaller than the central impression. Narrow gauge trackways of relatively long pace and stride.
-
(ii)
Other additional characters:
-
(a)
The vertical walls of the prints do not presuppose that the hoof of digit III is semi-ellipsoidal, semi-oval or hemispherical. The bottom of the track is also not curved (not hemispherical, semi-oval or semi-ellipsoidal).
-
(b)
The variation of the most open or closed shape, of straight or slightly curved branches of the mark of the edge of the hoof of digit III in CBR, is attributable to the movement of mud collapse.
-
(c)
The bottom without ornamentation (except the hoof channel) that characterizes the tracks of La Sierra de las Cabras, has no ichnotaxonomic value and is a character that does not serve to deduce the shape of the inside of the track.
-
(d)
The axis of the manus print and the pes print are not parallel, but form an angle (p^m). We could not determine if the angle to the midline of any of the axes (manus or pes orientation) is always greater or less than the other.
-
(e)
The pes print is smaller than that of the manus and is placed overlapping totally or partially in front of it. It should be noted that cases in which the manus prints are larger than pes prints have been reported from fossils of horses (e.g., Aramayo et al., 2015).
-
(a)
Despite the above, there are no criteria to assign them to any known species, nor sufficient criteria to define a new one. Among other things, isolated manus prints, or manus prints from any of the trackways cannot be studied in this site. There is only one case with two tracks (CBR38, Fig. 5C, D) in which one of them may be a complete mark of the manus without overprinting the pes.
The structures that deform the ichnites of this site are well illustrated. They are real (or true) tracks in the sense that they are direct structures (the skin and the hoof of the animal are those that have been in contact with the walls and base of the track hollow) but they are not stamps (faithful reflection of the sole of the pes) In addition, as in other vertebrates, the variability of ichnites is such that it is necessary to be cautious with the characters that could be considered ichnospecific.
It should be noted that only with the CBR small tracks, several species could be proposed if we consider the criteria used in the literature. For example, the circular or straight branches of the walls of digit III have been used to discern among different species, as well as the outward curvature of the branches. Another factor of distrust towards this type of classification is that most of the previous definitions of Hippipeda ichnospecies were based on samples that contain less than three footprints, and in some cases displaced from the original outcrops and without precise location (Hunt & Lucas, 2007).
We assign the CBR15 to CBR53 tracks to the ichnogenus Hippipeda Vialov, 1966, waiting for some new finds in nearby areas to provide complementary data on the manus marks and on the soles of their autopods, that could allow more specific determinations.
4.2.5 Other fossil tracks of Equoidea in Spain
The tracks of Equoidea previously described in Spain (Fig. 10, Suppl. Table 4) range from the Oligocene in Agramunt, (Santamaría et al., 1989–1990) to the Pliocene in Alicante (Lancis & Estévez, 1992). There is representation of traces of the lower Miocene of Navarra (Desojo and Los Arcos sites (Astibia et al., 2007), Cenicero (Díaz-Martínez et al., 2011) and in the upper Miocene of Hoya de la Sima in Murcia (Pérez-Lorente et al.,1999). The tracks of the Oligocene have been attributed to a paleotherid that is a primitive equoidea, and all the others to Hipparion, or Hippipeda. The paleotherid tracks still have three well developed front digits, while in all the others the main digit (III) is obvious.

Compilation of footprints attributed to Equoidea in several sites from Spain. A Plagiolophustipus monfalconensis Santamaría, López, Casanovas 1989. Agramunt, Eocene–Oligocene; B Hippparion footprints from Astibia et al (2007), Desojo site, Lower Miocene; C Hippparion footprints from Astibia et al (2007). Los Arcos site, Lower Miocene; D Hippipeda, this work, Sierra de las Cabras, Upper Miocene; E Hippipeda, Pérez-Lorente et al., 1999; Hoya de la Sima, Upper Miocene; F, Hippparion footprints, Lancis and Estévez (1982), Alicante, Pliocene
The degree of conservation of tracks of Sierra de las Cabras allows us to make observations on the trackmakers to adjust them to an animal pattern with respect to their body proportions. In CBR we are dealing with equids with three digits (II, III and IV) in each of the extremities of which only one (III) is functional. They had more slender hindlimbs than the forelimbs given that the pes print is much narrower and possibly shorter than that of the manus. It also suggests that the limbs were long, possibly longer than the glenoacetabular distance.
4.3 Perissodactyla. Rhynoceripeda Vialov, 1966
4.3.1 Rhynoceripeda footprints
Large tridactyl, mesaxonic tracks were recognized in the Sierra de las Cabras site, having four pad marks. Three of the rounded pads are placed on the front of the tracks and are produced by relatively wide adjacent digits and with a rounded termination; the fourth pad is larger than the previous ones and occupies the central and posterior part. This group of tracks includes those of the CBR67 trackway the isolated tracks CBR68 to CBR81, and those of the CBR82, CBR83 and CBR84 trackways (Fig. 11). Discriminative recognition features are not easily recognizable (Fig. 12) probably because: (i) the physical state of the mud at the time of footprint formation (CBR67.7); (ii) complex pes overprinting on the manus print (CBR67.5, CBR69), and; (iii) post-sedimentary fracturing and consequent ichnite deterioration (CBR67.8, CBR67.9). Despite this, 40 pes and at least 16 manus prints were distinguished in the Sierra de las Cabras site, which adds 56 ichnites to those described so far.

Sierra de las Cabras site map showing the studied surfaces and the reported Rhynoceripeda tracks

Compilation of Paleogene and Neogene tridactyl perissodactyl prints (except Macrauchenichnus). 1) pes print of Lophiopus (Ellenberger, 1980); 2) pes print of Palaeotheriipus (Ellenberger, 1980); 3) pes print of Plagiolophustipus (Ataabadi and Khazee, 2004, cf. Abassi et al., 2015); 4) and 5) perissodactyl prints from Utah (Lockley et al., 2014); 6) perissodactyl print from Colorado (Lockley et al., 2014); 7) right manus of Ronzotherichnus (Demathieu et al., 1984); 8) left pes of Ronzotherichnus (Demathieu et al., 1984); 9) and 10) pes of Rhynoceripeda (Costeur et al., 2009); 11) and 12) perissodactyl tracks (Rajkumar & Klein, 2014); 13) Plagiolophustipus (Santamaría et al., 1989); 14) and 15) Palaeotheriipus (Abbasi et al., 2015); 16) and 17) Plagiolophustipus (Murelaga et al., 2007); 18) group C (Casanovas and Santafé, 1974); 19) and 20) Palaeotheriipus (Gierlinski et al., 2007); 21) and 22) Rhynoceripeda manus and pes of adult individuals (Kordos, 1983); 23) Rhinoceripeda tracks of young individuals (Kordos, 1983); 24) rhinoceros footprint (Antunes et al., 2006); 25) and 26) Dicerothinihcnus, manus-pes couples (Guérin and Demathieu, 1983); 27) perisodactyls prints from China (Lockley et al., 1999); 28) ungulates from Peru (Lockley et al., 1999). CBR67.2, CBR67.6, CBR72 and CBR73, correspond to Rhynoceripeda of Sierra de las Cabras (this work), in which the position and general shape of the digits and prints can be appreciated
Of the pads of each of the three digits, the central is the widest and most advanced. Each of the digits has a single pad attached to that of the adjacent digit, so that the two lateral digits are touching the middle digit. The digits are elongated but placed in two positions: the major axis of the central digit is perpendicular to the track axis, whereas the major axis of the lateral digits is parallel. The heel is at the back of the center pad and is wide and rounded. Several structures found in some of the best preserved tracks, which we consider to be pes prints, were measured as they are those of the overprinted autopodium (Suppl. Table 5). The track length (Suppl. Table 5) ranges between 180 and 325 mm and the width between 207 and 296 mm, the average values being 257 mm (length) and 242 mm (width). The tracks, according to the ratio (l-a)/a value between 0.1 and -0.1 are in the limit between wide and narrow prints since the length is similar to the width. There are no indications about the relative size between the manus and the pes.
The termination and shape of the digits and the footprint shape are typical of an ungulated tridactyl trackmaker.
4.3.2 Rhynoceripeda trackways
The first of the four trackways (CBR67), is irregular and the others (CBR82, CBR83, CBR84) indicate a uniform progression. The measurements (Suppl. Table 5) indicate that the trackways are narrow and concordant in their values: P between 704 and 775 mm; z between 1422 and 1486 mm; Ap, more than 170º (except for CBR84 in which there is only one measurable angle). The measurements coincide with those of a trackway whose tracks are almost on a straight line. The Ar/a ratio of less than 0.5 indicates that the trackway is very narrow gauge.
If the less irregular sectors of the trackway are taken, pace (P, between 370 and 500 mm), trackway deviation (Ar = 81 mm) and trackway width (Lr between 342 and 384 mm) values are obtained. These data serve to classify the trackway as very narrow to wide gauge (Ar/a between 0.24 and 0.73). The width values of the relative trackway deviation (Ar/a) and trackway width indicate that many of the tracks are on the midline (Ar/a < 0.5) and that the furthest is 1/3 of its diameter from the midline. The trackway width is less than or equal to the space occupied by the width of two adjacent tracks.The z/l ratio is so variable that it is not usable.
4.3.3 Comparison with other tridactyl ungulate ichnites and nomenclature
Footrprtnts with features similar to CBR tracks and trackways have been reported for several ichnogenera (Suppl. Table 6). These are differentiated on the basis of the shape and size of digits, metapodial lobes, and digit pads (Abbassi et al., 2015). Tertiary tridactyl ungulates can be separated into non-periodactyls, exclusive of South America and perisodactyls from the rest of the world (Lockley et al., 1999). Previously Tertiary tridactyl ungulates, included the following (Fig. 12):
-
Lophiustipus Ellenberger, 1980, Palaeotheriipus Ellenberger, 1980, Plagiolophustipus Santamaría, López and Casanovas 1989–90. These ichnogenera show different size and digit shape than CBR perisodactyls, and are reported from strata of a different age.
-
Abbassi et al. (2015) and Rajkumar and Klein (2014) also cited perisodactyl ungulates but with long and separate digits.
-
The Argentine and Peruvian ichnites Eumacraucherichnus Aramayo & Manera de Bianco, 1987, and Macraucherichnus Angulo & Casamiquela, 1982 (Aramayo and Manera de Bianco, 2015; Krapovickas et al., 2009; Krapovickas and Vizcaino, 2016; Leonardi, 1994), have a similar shape, but these ichnites are of ungulates (Liptopterna, Macraucheriidae) that did not exist in Europe.
-
The oldest ungulate tracks attributed to rhinocerotids that have a pad on each digit were cited by Casanovas and Santafé (1974) with the name of Group C, Oligocene age (Fig. 12).
-
Ronzhoterichnus Demathieu, Gingsburg, Guérin and Truc 1984, also of Oligocene age, were described later.
-
Other similar tracks spread across different countries (Hungary, France, Namibia, Tanzania, Iran or Syria), with ages between the Miocene and Pleistocene: these are: Rhynoceripeda Vialov, 1966, Kordos (1983) Costeur et al, (2009); Dicerotinichnus Guérin and Demathieu (1993) and morphotypes cited by Ataabadi and Abbassi (2007) and Lockley and Tedrov (2009).
Costeur et al. (2009) quetioned the usage of the name of Ronzhoterichnus that Guérin and Demathieu gave to the French tracks and supported the name of Rhynoceripeda for reasons of priority.
We also assign the tridactyl tracks of ungulates of Sierra de las Cabras to the Rhynoceripeda ichnogenus, following the Costeur et al. (2009) criteria in accordance with the line followed in the ichnotaxonomic nomenclature applied in the Jumilla outcrops.
4.4 Carnivora. Bestiopeda Vialov, 1966
For this ichnological group we follow the criteria proposed by Buynevich (2015) based on Vialov (1966) that includes the traces of all carnivorous mammals within the ichnogenus Bestiopeda Vialov, 1966. According to Vialov, Bestiopeda includes all the traces of magnoorder Ferae (Creodontia or primitive carnivores and Carnivora or evolved carnivores). Because the variety of morphotypes is very large, Buynevich (2015) proposes a code in which the main ichnogenera group Bestiopeda is designated by the capital letter B and each of the different morphotypes, with a lowercase letter that follows the B. This refers to the zoological family or subfamily, in which its modern trackmakers are known at the species level, or to another possible division. In other works, all the mammal carnivorous tracks are grouped in the ichnogenus Bestiopeda (B) and then divided into subgenres, e.g. dog tracks (Canipeda, Bc), felid (Felipeda, Bf), bear (Ursipeda, Bu), weasel (Mustelipeda, Bm), seal and related pinnipeds (Phocipeda, Bp), and so on.
4.4.1 Characterization of Canipeda (Bc) and Felipeda (Bf).
According to Graham et al. (1994) canids (wolves, coyotes, foxes and domestic dogs) and felids (cats, panthers, and others) have similar tracks in general: paraxonic and homopodial tetradactyl, although manus prints tend to be larger than these of the pes prints (Melchor et al., 2018). The ichnites are composed of four small digit pads and a large interdigital pad, which is bigger in felids than in canids. The large pad has received different names in the literature: central (Rodríguez de la Rosa et al., 2012; Scrivner & Botjer, 1986; Thenius, 1967; Neto, 2009); interdigital (Antón et al., 2004; Graham, et al., 1994); metapodial (Reynolds & Milner, 2012; Sarjeant et al., 2002; Sarjeant & Lanston, 1994; Melchor et al., 2018);sole or heel (Panin & Avram, 1962; Antón et al. 1993; Williamson & Lucas, 1996; Remeika, 2001; Neto de Carvalho et al., 2016);or principal (Scrivner & Botjer, 1986).
Both the manus and pes prints are tetradactyl. The posterior margin of the interdigital pad is trilobed in felids and canids. The four digit pads are oval and anteroposteriorly elongated.
The tracks have also received different names including digitigrade (Neto et al, 2016; Remeika, 2001); semiplantigrade (Rodríguez de la Rosa et al., 2012; Sarjeantet al., 2002); semidigitigrade (Sarjeant et al., 2002): between plantigrade and semiplantigrade (Sarjeant et al., 2002); plantigrade (Remeika, 2001; Sarjeant et al., 2002); and subdigitigrade to digitigrade (Melchor et al., 2018). The tracks of canids are commonly longer than wide, unlike felids (Melchor et al., 2018). Also, the anterior margin of the interdigital pad is monolobate in canids and bilobate in felids and generally there are claw marks on the tracks of the canids and not on those of the felids. The support of the feet of these animals on the ground is performed on the distal digit pads (phalanges 2–3) of the four digits and on the metatarsal pad (metatarsal-phalanx 1). The phalanges are not placed parallel to the ground (Sarjeant et al., 2002) so that the digit pads protect the junction between the ungular and the anterior phalanx, and the central pad protects the articulation of the metapodial bones with the proximal part of the first four phalanges. The metapodial bones are elevated in their proximal part so that their distal part is closest to the ground.
Canipeda Panin & Avram, 1962 can be defined with the diagnosis emended by Sarjeant et al. (2002), and recently emended by Melchor et al. (2018) which is translated as follows: Digitigrade to semi-digitigrade, tetradactyl (digits II to V), paraxonic tracks; longer than wide tracks; quadruped and homopodial trackways. Elliptical and separate digit pads, of similar size and with sharp nail marks. The digit pads, one per digit, form an arc in front of a large metapodial pad from which they are well separated and arranged in a bilateral symmetrical pattern along the antero-posterior axis.
For the ichnogenus Felipeda (Bf) we also use the diagnosis of Panin and Avram (1962) emended by Melchor et al (2018): Digitigrade to semi-digitigrade, tetradactyl, paraxonic tracks, generally wider than long tracks; quadruped and homopodial trackways. Rounded and separate digital pads, of similar size and without nail marks. The digit pads form an arc in front of a large metapodial pad from which they are well separated.
4.4.2 Canipeda (Bc) of Sierra de las Cabras
A total of 29 ichnites of the Sierra de las Cabras tracksite are attributed to Canipeda (Bc). Fifteen of these are isolated tracks (CBR86 to CBR100), a pair of tracks (CBR85), and a trackway (CBR101) with twelve tracks (Fig. 13) considered to be from the ichnogenus Canipeda. All the ichnites are present in surface 1 except CBR101 trackway printed on surface 3. Except those of trackway CBR101 (manus and pes prints), the rest we attribute to quadruped animals in which the overprint of the pes on the manus is total.

Sierra de las Cabras site map showing the studied surfaces and the reported trackways, pairs of tracks, and isolated prints of Canipeda
Most of the isolated tracks are deformed by the adjacent tracks of other animals and by the fall of mud into the hollow during the W-K phases. The mud was not very viscous because the autopodia penetrate the ground, but with enough consistency so that the walls of the track combine inwards and completely collapse in some digits.
The best preserved ichnite in the set (CBR92) was used for overall description (Fig. 14). It is a tetradactyl track with a pad on each digit; behind and in the center of the track is a wide interdigital pad. The digit marks are shaped like a drop, rounded proximally and terminated distally at very sharp points. The digit pads are separated from each other and from the interdigital pad. This is the only track in which the morphometric characters are visible, but it shows variable sizes in the digit pads and consequently, any generalization about the size of digit pads is not appropriate from the observations in these ichnites. The digit pads occupy a broad front, transversely to the axis of the track. The width of the track in the central pad is smaller than in the area of the digit pads: the track is narrower in the back half than in the front. Proximally it is bilobed, while in the distal termination it is a kind of angle with a round vertex. The sides of the angle are concave.CBR92 is somewhat longer (l = 121 mm) than wide (a = 106 mm); the length has been measured parallel to the axis of the track, from the heel to the tip of the most protruding nail.

Upper part: photography, drawing and topography of CBR92. Lower part: drawings of CBR86 and CBR88;0 the two prints are similar to Bestiopeda maxima Abel, 1935. The shape is due to the collapse of the walls, very evident in the traces of the digits
The isolated tracks CBR86, CBR88 and CBR98 are similar to the first drawn carnivorous tracks (Bestiopeda maxima Abel, 1935). These are marks defined by a round central zone from which four tips similar to spicules or bradawls protrude (Fig. 14).
The pointed shape of the digit marks suggests digits with sharp termination (nails or claws) and discard rounded terminations. The mud that surrounded the hollow of the ichnites, at the time of their formation, collapses and the contour bulges inwards, so that the digit gaps narrow and the pointed termination of the distal digit marks becomes accentuated. The acuminate termination of the ichnites that we interpret as nail or claw marks, we assign to Canipeda.
The sequence CBR101.1 to CBR101.7 (Fig. 15) forms a continuous trackway of twelve tracks (5 pes and 7 manus?). The measurements (Suppl. Table 7) indicate that the tracks are equidimensional (l = a = 97 mm), the very long pace and stride (respectively 592 and 1156 mm) and the z/l ratio (20) very large, all measurements being congruent with relatively long and thin limbs. CBR101 is a wide gauge trackway (Ar/a = 0.77) and is consistent with the pace angle (Ap = 152º), which is not very open. The orientation of the tracks is negative (sensu Leonardi, 1987). The trackway shows the following features:
-
the pairs of tracks are alternately placed on each side of the midline.
-
the separation of manus and pes from the midline is similar.
-
the closest pairs are of alternate laterality, i.e., alternate manus and pes are closer than manus and pes of the same side.
-
the average distance (Fig. 15, p-m) of the manus and the pes on the same side is 695 mm and the average distance of the manus and the pes of its side (Fig. 15, m-p) is 500 mm. The p-m value is given by the distance that separates the two autopods of the same side when both are resting on the ground; the m-p value is the distance that the pes print overpasses that of the manus on the same side (overlap): the pes overlaps the manus print a distance that is about five times its length.

CBR101 trackway. From left to right: Ichnite reference; trackway prints; trackway main lines; manus pace; pes pace; pes – manus separation on the same side in simultaneous support (p-m); overlap or distance that advances the pes print relative to the manus position (m-p)
Trackways comparable to those from Sierra de las Cabras are produced by camels during amble walking (Pérez-Lorente et al., 2009; Thompson et al., 2007), but in CBR101 the trackway is wide (the separation between the center of the tracks is about 1.5 times the width of the track) and the pes-manus separation of the same side is greater than the pes-manus of alternate sides.
The trackway is assimilable to the type 2–2 of Rezendes (1999), where the pes passes the manus of the same side (500 mm) to a distance almost similar to half of the stride (z = 1156 mm), and also to a moderate escape print (Richarz, 2006) in which the tracks of each side are well separated from the midline and in which the alternate manus and pes prints are closer than the manus and pes on the same side.
Amble walking implies very narrow trackways (the ratio Ar/a ≤ 1 or also Lr ≤ 2a). CBR101 should be produced by a trotting or fast-running animal (Rezendes, 1999; Richarz, 2006) or (pace) amble trot (Thompson et al., 2007). Kienapfel et al. (2014) reported a similar track produced by fast tölt (fast amble) in their Fig. 5b. These authors point out that almost all large mammals with hooves and carnivores make very similar movements.
The height of the acetabulum (h), according to the formula (7,41 × l) by Melchor et al. (2018) is 720 mm. The glenoacetabular distance following the method of Demathieu (1986) and considering that the gait is amble (Richarz, 2006) is 653 mm. This means that the limbs were slightly longer than the glenoacetabular distance. The average relative stride (z/h) (Thulborn, 1990) is 11, so the displacement of the animals includes an aerial phase. Using velocity algorithms proposed by Alexander (1976), Demathieu (1986) and Thulborn (1990), we get 4.8, 3.8 and 2.7 km/h respectively, i.e. a range of estimates probably too large to be reliable.
4.4.3 Comparison with fossil ichnites attributed to canids
The length of the ichnites from Sierra de las Cabras site (Suppl. Table 7), including the loose samples stored in the Jerónimo Molina Museum (Fig. 16) is very large if compared by previously reported ichnites attributed to canids in the literature (Suppl. Table 8). Twelve footprints exceed 100 mm in length and five of them are larger than 120 mm. It should be noted that they are clearly larger than the big footprints reported by Neto et al. (2016) in the Pleistocene of Portugal and attributed to wolves, which average 80 mm in length. The length of the rest of the Tertiary Canipeda footprints previously reported range between 37 and 65 mm.

Four samples from loose fragments found in the Sierra de las Cabras site, kept at the Jerónimo Molina Museum (Jumilla). The two upper images and lower left image are from Canipeda. The lower right of Felipeda. Scale is 10 × 2 cm in all images
The large size of CBR footprints could make them comparable to Carnotipus López et al., 1992, an ichnogenus defined in the Lower Miocene of the Ebro basin (Spain) and whose length (up to the tip of the nails) varies between 108 and 122 mm. However, in this case, the digits are fine and long, narrowing progressively until they reach an acuminate tip; and also, the claw traces are at the same level as the rest of the track do not come out at a level higher than the sole as typically occur in canids.
Melchor et al. (2018) include in Canipeda the traces attributed to herpestids of the Salinas de Añana by Antón et al. (2004), and the tracks found by Rabal-Garcés and Díaz-Martínez (2010) in the Eocene of the Santa Cruz de la Serós. Finally, we ant to note that at the nearby site of the Hoya de la Sima (Jumilla), a trackway with three tracks comparable to those described in Sierra de las Cabras site was reported by Pérez-Lorente et al. (1997) but not attributed to canines or felids because of the lack of clear criteria to discriminate. These tracks are longer (119 mm) than wide (108 mm) and consist of four rounded and small front marks, and a central, rear and larger pad. These tracks are probably closer to Canipeda than Felipeda because of their size and width to length ratio.
4.4.4 Felipeda (Bf) of Sierra de las Cabras
10 tetradactyl tracks (Fig. 17) are distinguished. Two of the tracks form a pair (CBR102), another is an isolated ichnite (CBR104) and the other seven are part of a trackway. They are defined by four rounded and not acuminate digit pads, and a central posterior pad, larger than the other ones. The digit pads are separated from each other in almost all the tracks, and also from the posterior central pad. This pad does not have a uniform shape in all tracks, nor in the tracks of a single trackway (CBR103).

Sierra de las Cabras site map showing the studied surfaces and the reported trackways, pairs of tracks, and isolated prints of Felipeda
The tracks (Suppl. Table 8) are large (> 80 mm long, cf. Reynolds & Milner, 2012), and wider (a = 157 mm) than long (l = 131 mm), except those of trackway CBR103 (a = 95 mm, l = 121 mm). The digit pads are rounded, somewhat elongated and with round distal and proximal endings. The size of the pads is different but there is no pattern of common distribution to all traces. The main pad is much larger than that of the digits (Fig. 18) and with a trapezoidal contour with rounded edges. The front part of the main pad is bilobed, and the back is trilobed in some ichnites (CBR102.2, CBR104). The digit pads are in front and to the sides of the main pad, forming an arc; the proximal part of the medial and lateral digit pads is behind the distal end of the main pad.

Left: pair CBR102, see text for explanation. Right: photography of CBR102.2, drag grooves of the autopodium
There is only one trackway, which is very narrow gauge (Suppl. Table 7. In it, the stride length (1071 mm) is almost double that of the pace (549 mm), according to the 180º value of the pace angle. The z/l ratio (9) is typical of animals with long, thin limbs. Although the trackway is apparently biped, the trackmakers are quadrupeds. The overprinting is total in all footprints with the exception of the CBR104. The glenoacetabular distance ranges between 795 (evolved alternating gait) and 1059 mm (ambler gait) following the scheme by Demathieu (1970).
These tracks are typical Felipeda (Bf). According to the size and weight data of present-day felids (Graham et al., 1994) the trackmaker would be a big felid, probably over 130 kg in weight.
4.4.5 Ursipeda (Bu) of Sierra de las Cabras
They are pentadactyl, plantigrade tracks of a quadruped animal that overprints the pes on the manus print during the march. All tracks are irregular (Figs. 19, 20) and show total or partial overprinting of the pes on the manus print, except CBR109.2 (probable manus print) and CBR106 where the interpretation is more doubtful. There are four trackways (three of which are incomplete CBR107, CBR108 and CBR110 and one complete CBR111); two pairs of tracks (CBR105, CBR109), and uninterpreted marks in which there may be prints of two manus and one pes (CBR106). In trackway CBR108, ichnite CBR108.6 is in a fractured area that may be responsible for its irregular and fragmented outline. Considering the ichnites as overprinted pes-manus groups, a total of 26 ichnites of this morphotype are reported.

Sierra de las Cabras site map showing the studied surfaces and the reported prints and trackways of Ursipeda

Photograph and drawing of ichnites CBR108.5 (above) and CBR108.7 (below). Traces of long claws attached to the digit pads can be observed. Scale bar: 10 cm
Accompanying the ichnites there are abundant structures that denote soft soil, and commonly the toe nails penetrate the mud destroying the marks of the manus prints. In fact there are no complete manus marks, except perhaps CBR109.2, because all the others are overprinted by the pes. The mark of the manus is usually ahead of that of the pes, and deformed by it. There are tracks that have all or part of the front part of the manus from which nail marks are preserved, sometimes with the distal part of the digits. The triangular shape of the pes print is well recognized.
Each digit is represented by a relatively elongated and separated pad as seen in CBR105. The five digits are not always distinguished due to the deformation of the mud. The digits have sharp, thin and long, generally curved nails, each of which is associated with the corresponding digit pad. The nails penetrate the mud, leaving filiform elongated structures. The size of the digit pads and that of the nails cannot be measured on these tracks. Both the envelope of the tip of the nails and the contact between the digit pads with the metatarsal is arched (CBR105.1, CBR108.5).
Separated from the digit pads there is, in both the manus and the pes prints, a wide pad, perhaps somewhat wider than the area occupied by the digits (as the outline on the distal part of the pad slightly protrudes from the area of digits). On the pes, the plantar pad narrows backwards, so that the outline is triangular with a rounded vertex corresponding to the heel. The sole is slightly separated in two parts by a smooth transverse elevation that ends in a lateral recess of the outline.
The width of the prints ranges between 92 and 235 mm and the length between 119 and 346 mm (Suppl. Table 7). It should be noted that these measurements derive from overprinted ichnites in which the outline of the manus is unknown and that of the pes is deformed. The only tracks in which the outline is distinguished with precision are CBR108.4 and CBR108.5 in which the average length (including the nail mark) is 342 mm and the average width of 231 mm (Suppl. Table 7). It is possible that the manus prints are wider than those of the pes (eg: CBR107). Tracks CBR107.2, CBR111.3 and CBR111.4 distinguish pes prints separated from the manus. The least deformed are those of the second trackway in which the average length is 148 mm and the width 155 mm.
Some additional tracks of this type are found in loose samples retrieved from Sierra de las Cabras and conserved in the Jerónimo Molina Museum (Jumilla), with the particularity that in one of the samples an isolated manus mark was found (Fig. 21).

Left: topography of CBR107.2. Right: above, a manus print and (below) a fragment of pes print. Samples found in loose fragments of Sierra de las Cabras tracksite. Scale 10 × 2 cm
4.4.6 Nomenclature and comparison with other ursid ichnites
Three ichnogeneric names have been proposed for fossil traces of ursids (Suppl. Table 9): Platykopus Sarjeant, Reynolds and Kissel-Jones 2002, which is also cited by Abbassi (2010) and Lockley, Reynolds, et al. (2007); Ursichnus Diedrich, 2011, also mentioned by Aramayo et al. (2015); and Ursipeda (Bu) Buynevich, 2015. In the Iberian Peninsula, fossil bear tracks were reported from the Pliocene of Alicante (“ursid tracks” of Lancis & Estévez, 1992).
These tracks of ursids differ from CBR ones in the following aspects (Fig. 22):
-
Platykopus has the phalangian pads with two lobes and they are attached to the central pad in manus and pes (Sarjeant et al., 2002). In general they are smaller tracks than those from CBR.
-
Ursichnus has wide, shovel-shaped nails (Diedrich, 2011. Usually, these ichnites are smaller than those from CBR. It should be noted that they have been described in a very different period (upper Pleistocene)
-
The “ursid traces” of Lancis and Estévez (1992) have a size (with lp slightly smaller than 300 mm) comparable to those from CBR, but do not have ichnotaxonomic characters applicable to this discrimination.

Compilation of Ursid prints from the literature. A–J: Platycopus fossil prints (A and B respectively right pes and right manus, from Sarjeant et al., 2002; C–H are cast copies of the Raymond Alf Museum, from Lockley, Culver, et al., 2007; Lockley, Reynolds, et al., 2007; and J shows the pair br5-br6 of Abassi, 2010). K–M: Ursichnius fossil prints (K is a left manus print, ref. UrsPest-track 2 and L is a pes print, ref. UrsPest-track 1, both from Diedrich, 2011; and M is the manus-pes holotype of Aramayo et al., 2015). N and O Ursid footprints (N is a right manus and O a right pes from Lancis and Estévez (1992). P Ursipeda from this paper, showing fragments of right manus and pes (CBR107.2), with the arrow indicating mud fall
Thus, the CBR ursid ichnites canot be accurately assigned to any of the mentioned ichnogenera. Because in this work we follow classification schemes derived from Vialov (1965, 1966) and subsequent works of other authors we assign the ichnites to the Ursipeda (Bu) in the sense of Buynevich (2015).
4.5 Rodentipeda Vialov, 1966
4.5.1 Rodentipeda of Sierra de las Cabras
There are two biped trackways (CBR112 and CBR113) (Fig. 23), which are paired, i.e., each of them formed by pairs of separate and symmetrical pes prints which are repeated in each step (Fig. 23). In total, 20 footprints are reported (10 footprints in CBR112 and 10 in CBR113). Each pair (Fig. 24) is the trace of two autopods (left and right pes), indicating a displacement by jumps.

Left, Sierra de las Cabras site map showing the studied surfaces and (below) the reported trackways of rodents. Right, CBR112 and CBR113 footprints. The first footprint of the trackways is the left footprint. Observe the forward position of the outer footprint in each step, more evident in CBR113. In this sequence (CBR113), the rear semilunar elevation is placed towards the outside of the midline

Above, image and topography of CBR112.2; the equidistance of the contour lines is 2 mm. Below left, drawing, photograph (center) and contour lines (right) of CBR113.2; equidistance of the contour lines is 2 mm. Scale 10 × 2 cm
In CBR112, the elements of each pair (Figs. 24, 25; Suppl. Table10) are relatively long (l = 47 mm) and narrow (a = 18 mm). Both ichnites show a distal extreme which is rounded and shallow, and a proximal extreme, which is narrower and deeper. The outline of each autopodium has the shape of a drop.

Compilation of fossil trackways of small rodents from the literature: A Ameghinichnus Casamiquela, 1964, Rainforth and Lockley (1996); B Brasilichnium Leonardi, 1994, Rainforth and Lockley (1996); C Brasilichnium Hamblin and Foster (2000); D–H Mammal?, Leonardi (1980, 1994), Lockley, Culver, et al. (2007); I–K Rodent, Lockley, Culver, et al. (2007); L and M Musaltipes Lockley & Milner, 2014, Lockley, Culver, et al. (2007); N–S, Brasilichnium, D’Orazi et al. (2017b); T Musaltipes Lockley & Milner, 2014; U Hypnomyx trackways, Quintana and Arnau (2004); V and W this work. Scale, 10 cm
The footprints of CBR113 (Figs. 24, 25; Suppl. Table 10) have the same morphological characteristics, although they are slightly smaller (l = 30 mm; a = 13 mm). Remarkably, they have a crescent-shaped hollow behind, bounded by a posterior crest (Figs. 23, 24, 25). The straight distance between the two ends of these crescent-shape hollows varies between 39 and 74 mm depending of the print, with an average of 56 mm.
No pads (digit or sole) of any kind are distinguished. No digit marks are distinguished, and only small digits may remain in the wide part of one of the ichnites (CBR113.2. Figure 24). Nail or hoof marks are not seen either. There is no tail mark. The walls of the footprint are vertical or slightly bulging inwards. The penetration of the metatarsal zone indicates that the mud was not very viscous, but the verticality of the deeper walls suggests it was quite cohesive. Some collapse could have happened being responsible for the slight bulging and probable approximation of the walls. The longitudinal axes of both tracks in each pair are subparallel and in turn are parallel or form a very small angle with the midline of the trackway.
In short, they are tiny drop-shape footprints placed in pairs, with the frontal part wider and shallower than the rear. The size end shape are typical of rodents (Lockley, Culver, et al., 2007).
4.5.2 Trackways
CBR112 and CBR113 are two biped trackways with tiny tracks (in the sense of Garcia-Raguel et al., 2009) and paired. The sequence of steps is the periodic repetition of the pairs of footprints that must be pes prints (rear). We consider that the pace (P) is the distance between the geometric center of two pairs of successive tracks; this distance is approximately the same for both the pace and the stride, as it is the separation between the footprints of two alternating pes and two pes of the same side. In both trackways the steps are very long compared to the length of the footprints (Suppl. Table 10). The value of P/l is 8.8 (CBR112) and 15.6 (CBR113). Other characteristics related to the position of the CBR113 tracks in the trackway are: relatively open pace angle (167º); trackway width (Lr) of 87 mm, i.e. almost three times the length of the footprint); and trackway deviation (Ar) of 26 mm, i.e. wide gauge trackway (Ar/a = 0.7; the footprints do not touch the midline).
The trackways indicate oscillating jumps in relation to the trajectory: each pair of footprints moves alternately to the left or to the right of what would be the midline (Figs. 25, 26). The pairs of footprints are nearly symmetrical in the sense that neither of the two ichnites is markedly ahead of the other, but in CBR113, where the oscillating path is more patent, the outer footprint is slightly more advanced than that of the inner one (Figs. 23, 25, 26).

Proposed model for the formation of the semilunar rear structure of CBR113 by a triangular or drop-shaped foot. Left column: position of the pes during the imprint movement, with A–C indicating foot contact; D the position of the foot at the end of T-phase; E the termination of W-phase; and F, the bottom, final structure after K-phase. Right column: position of the body of the trackmaker in successive jumps
Many footprints of jumping trackmakers have an area behind where the tracking surface is deformed due to the characteristics of the sediment, the inclination of the tracking surface, and the progression of the trackmaker (Buck et al., 2016; D’Orazzi et al., 2017a, 2017b). In this sense, it should be noted that all the previously described structures of jumping are in sandy material, many located in dune slopes (deduced from the orientation of the sliding sand when stepping). Both ascending and descending trackways have been reported (Buck et al., 2016; D’Orazzi et al., 2017a, 2017b).
Trackway CBR113 presents similarities with the trackway reported by Hamblin and Foster (2000) in the Jurassic Navajo Sandstone (Fig. 25C). However, the origin of that trackway must be different, as both the type of sedimentary material and the slope of the tracking surface were very different from those of CBR112 and CBR113 which were developed on a muddy horizontal consisting of soft mud with microbial mats. We associate the deformation behind the footprint with the impact of the foot when entering in the mud and the latter impulse to leap forward. During the subsequent partial mud collapse the surface descends leaving the rear crest. The model, following the phase terminology of Thulborn and Wade (1989) is as follows (Fig. 26):
-
During phase T the foot touches, penetrates the ground and begins to form the rim of mud extrusion around the footprint.
-
During phase W the body of the trackmaker advances and the greater pressure falls on the heel, narrower than the sole or front digit section. The heel sinks further. The movement and pressure to start the next jump, move mud backwards and increase the volume of the rear rim.
-
During phase K and after it, part of the mud of the rear rim falls, forming the crescent structure (Figs. 23, 24, 25, 26).
The type of jump in which the contact with the ground of one foot is slightly out of phase with respect to the other, is concordant with D’Orazi et al. (2017a)—the footprints are neither synchronous nor symmetrical. The jumping mode (skipping) can be compared to the “jumping rope” model. Dissymmetry should refer to decoupled feet since one pes is slightly advanced with respect to the other, and in CBR113 the area enclosed in the outline is different on each print in each pair. In CBR113 the footprint of the inner foot is larger and slightly behind with respect to the external footprint. The same pattern is also intuited in CBR112 (Fig. 26). The inner foot is the one that first steps on the ground because it is slightly behind and leaves a larger footprint than that of the external one. We say that the largest footprint is the one that is first printed, because it will be the one that puts the most pressure on the ground.
The rear crescent extrusion that accompanies the footprints of CBR113 is not symmetrical with respect to the footprints or to the main trajectory. The symmetry axis of the crescent is placed in a more external position of the trackway midline than the axis between the pair of footprints. If the mechanism proposed above (Fig. 26) for the formation of these structures is correct, the push of the feet would also be outwards from the midline and thus alternating to one side and the other. The animal would move like a pendulum, transversely to the trajectory, advancing in leaps with slightly out of phase feet support.
Despite the differences between the two trackways (larger steps and footprints and lack of crescent structures in CBR113), we assume that they were left by two specimens of the same taxonomic group because of the biomorphic and morphometric similarities. Also, the extramorphological features, the type and direction of the trackways and the paleoenvironmental situation point more likely to two individuals of the same group than to the two taxonomically different animals.
4.5.3 Jumping trackmakers
As explained, CBR112 and CBR113 are considered to correspond to bipedal jump trackways. It should be mentioned that some reported ichnites with parallel footprints have been attributed to quadruped trackways produces by galloping, in which the manus prints are completely overprinted by the pes. Given the structural differences between the two CBR trackways, if they were quadruped both types of footprints (pes and manus) would be observable in any ichnite, which is not the case.
In the ichnites of CBR112 and CBR113 the back is narrower than the front, reflecting the shape of the foot. This is because at least the metatarsal is printed, which is narrower than the acropodium, or because the foot is narrower in the heel. As already mentioned, these small, narrow metatarsal forms are typical of rodents (Lockley, Culver, et al., 2007).
The footprints are also noticeably deeper in the back area than in the front (Fig. 26). It is likely that this difference in depth is due more to the mechanics of the jump than to the distribution of weight on the sole. We can assume that the spring movement of the proximal joint of the functional foot is responsible for the greater descent of the heel, which in turn is narrower than the front part of the foot.
All bipedal jump trackways with ichnites of these dimensions and proportions have been attributed to vertebrates. Cases are known since the Mesozoic, were Leonardi (1994) initially attributed to dinosaurs certain tridactyl ichnites (Brasilichnium, Fig. 25B), although later. (Rainforth & Lockley, 1996) pointed out that they more probably correspond to other reptiles, therapsids or mammals. Despite the trackmaker attribution becoming very debatable the older ol ichnites are (Jurassic age or earlier), it should be noted that the autopodium in those cases left traces that were wider than long, i.e., clearly different to those from Sierra de las Cabras.
In Spain, fossil trackways of jumping animals have been reported in fossil dunes of Pleistocene age (Fig. 25U), and attributed to Hynomyx footprints (Quintana & Arnau, 2004). In some of the sequences manus marks are also observed, suggesting quadrupedal gait in which the manus are overlapped by the pes. Also, they can be differentiated from those of CBR by the relatively wider shape of the prints.
Most reported jump trackways were found in aeolian dunes or even in volcanic ashes and tuffs, probably arid settings where animals obtain water from solid aliment (Hamblin & Foster, 2000; Rainfort & Lockley, 1996). At the moment the progression by jumps is typical only of certain birds and some very small mammals (rodents), and is not normal in large vertebrates except in kangaroos. There are small jumping rodents that live in forests, meadows, swamps and marshes. According to Lockley, Culver, et al. (2007) there are three families of live rodents which include jumping forms: Zapodidae, Dipodidae and Heteromyidae. Only Zapodidae are able to drink water and live in humid environments (Lockley & Milner, 2014; Lockley, Culver, et al., 2007).
In CBR112 and CBR113, neither the number nor the shape of the digits can be distinguished, nor is there a non-deformed outline of the autopodium. These limitations allo us only to specify on the trackmakers their attribution to Zapodidae by their way of progression, the size and outline of the footprints and the habitat of the site.
4.6 Unidentified tracks
Figure 27 shows the location of 65 ichnites (CBR114 to CBR188) found in Sierra de las Cabras tracksite that show characteristic features that allow them to be associated with any ichnotaxonomic group. These ichnites are heterogeneous, heterometric and do not show any apparent ordering. These do not include either avian or invertebrate (?) traces of the site, which will be the focus of other specific studies.

Sierra de las Cabras site map showing the studied surfaces and the reported unidentified prints
CBR114, CBR115, CBR117 and CBR119 could probably be part of a trackway as they are roughly aligned and show a suitable distance between them; also, they have similar size and rounded shape, and show the same type of circular tension structures inside, produced by the breakdown of partially consolidated sediment (with microbial mats) when the trackmaker stepped on the surface. Ichnites CBR121 and CBR123 are rounded and have comparable size to those assigned to Canipeda as reported before in this paper. Ichnites CBR134 and CBR147 are also the same in size and shape as the aviforms that only print a digit like those of trackways CBR3 and CBR5. Finally, CBR164 and CBR165 could be associated with Ursipeda because of their size and some ornamentation features, although they are a rough surface and are very imperfect. The stretch marks of the upper part of the site, of unknown origin until now, remain to be noted.
4.7 Artiodacyla Suipeda Vialov, 1966
Around the tracksite, we have found some ichnites in several loose pieces of strata coming very probably from the same stratigraphic levels. Despite these tracks being eroded with variable intensity, some of them show some recognizable morphological characters. Among these, there are rounded tracks of various sizes (small ones could correspond to Canipeda and, or Felipeda, and large ones to Rhinoceripeda and Ursipeda).
In one of the pieces, three tetradactyl artiodactyl tracks were found (CBR188, CBR189 and CBR190, Figs. 28, 29). These ichnites have neither the same orientation nor the same size, so they do not belong to the same trackway. The footprints are longer than wide (with length/width ratios of 14//8.8, 8//5.9 and 6.7//4.8 cm, respectively) and show the mark of two elongated and parallel front digits and two smaller back digits. The digit marks show smooth walls and floor, like the ones that are typically produced by hooves. The front prints (which would correspond to the primary hooves) are slightly wider than the rear ones and much longer. These later (secondary hooves), which are present in all three tracks, are oval and shallower than the front ones. In the best preserved track (CBR188) the marks on the back digits (secondary hooves) are not aligned with those on the front ones and are more separated from the footprint axis. (Figs. 28, 29). Pecoripeds, ruminants and suids are among the animals capable of producing this type of tracks. Although the material herein reported does not allow a precise attribution, the fact that separation of the marks produced by secondary hooves is greater than that of the primary hooves, suggest that the trackmaker could probably have been a suid.

CBR189, CBR190, CBR191. Artiodactyl tracks, possible Suipeda

CBR188 contour lines and photography. See text for explanation
5 Conclusions
This paper describes the footprints of seven ichnogenus belonging to Mammalia, found in the Sierra de las Cabras tracksite, a new and exceptional vertebrate ichnite site locate in the Jumilla area (Murcia, SE Spain) in the Prebetic Zone, and chronostratigraphically framed in the uppermost Tortonian to lowermost Messinian interval.
The Sierra de las Cabras site is roughly coetaneous to the nearby Hoya de la Sima tracksite located only several kilometers away but developed under different environmental conditions (saline lakes). Together, the seven mammal ichnogenera of Sierra de las Cabras and the two pecoriped ones and the probable proboscid previously reported from the Hoya de la Sima site, constitute an exceptional record of the vertebrate diversity, and population of the area.
The grouping of footprints and their orientation do not necessarily imply, due to the extension of the sites, that the animals had gregarious behavior, although the likelihood is that there were groups that wandered around these places. The diversity of trajectories both within the trackways of the same ichnotype and the total of those found, speak of groups of animals that roam the area, in principle without a defined direction, and therefore without the presence of defined natural conditions (water line, presence of a pond or any type of natural barrier).While there are groups of animals in which their behavior is gregarious and therefore it is normal to find traces in relatively dense groups (Camelipeda, Hippipeda), the accumulations of other types of footprints in Sierra de las Cabras do not allow us to reach conclusions in this sense, It is likely that the prints of mammals are at least from family groups because they are paired trackways.
The mammal footprints of Sierra de las Cabras are accompanied by other ichnites attributable to birds and arthropods, which will be the subject of successive specific papers.
Availability of data and material
The data and matherial supporting their findings can be found from the text citations and localizations.
References
Abbassi, N. (2010). Vertebrate footprints from the Miocene Upper Red Formation, Shokorchi Area, Zanjan Province, NW Iran. Ichnos, 17, 115–126. https://doi.org/10.1080/10420941003659493
Abbassi, N., Lucas, S. G., & Reza, G. (2015). First report of Oligocene vertebrate footprints from Iran. Palaeogeograohy, Palaeoclimatology, Palaeoecology, 440, 78–89. https://doi.org/10.1016/j.palaeo.2015.08.039
Abel, O. (1935). Vorzeitliche Lebenspuren. Gustav Fischer Jena.
Acosta, G., & Simonetti, J. A. (1999). Guía de huellas de once especies de mamíferos del bosque templado chileno. Boletín Del Museo Nacional De Historia Natural, Chile, 48, 19–27.
Agustí, J., & Antón, M. (2002). Mammoths, sabertooths, and hominids: 65 Million years of mammalian evolution in Europe. Columbia University Press.
Alexander, R. M. (1976). Estimates of speed of dinosaurs. Nature, 261, 129–130.
Allen, J. R. L. (1997). Subfossil mammalian tracks (Flandrian) in the Severn Estuary S.W. Britain: Mechanics of formation, preservation and distribution. Philosophical Transactions of the Royal Society of London, B, 352, 481–551. https://doi.org/10.1098/rstb.1997.0035
Angulo, R. J., & Casamiquela, R. M. (1982). Estudio estratigráfico de las unidades aflorantes en los acantilados de la costa norte del Golfo de San Matías (Río Negro y extremo austral de Buenos Aires) entre los meridianos 62º30’ y 64º30’ W. Mundo Ameghiniano, 2, 20–85.
Anton, M., Lopez, G., & Santamaria, R. (1993). Estudio preliminar de la icnofauna miocena del yacimiento de Salinas de Añana (provincia de Álava). Comunicaciones de las IX Jornadas de Paleontología, Málaga, 23–28.
Antón, M., López, G., & Santamaria, R. (2004). Carnivore trackways from the Miocene site of Salinas de Araña (Alava, Spain). Ichnos, 11, 371–384. https://doi.org/10.1080/10420940490444951
Antunes, M. T., Balbino, A. C., & Ginsburg, G. (2006). Miocene Mammalian footprints in coprolites from Lisbon, Portugal. Annales De Paléontologie, 92, 13–30.
Aramayo, S. A., & Manera de Bianco, T. (1987). Hallazgo de una icnofauna continental (Pleistoceno tardío) en la localidad de Pehuen Có, Provincia de Buenos Aires, Argentina. Parte II: Carnivora, Artiodactyla y Aves. IV Congreso Latinoamericano De Paleontología Actas, 1, 532–547.
Aramayo, S. A., & Manera de Bianco, T. (1996). Edad y nuevos hallazgos de icnitas de mamíferos y aves en el yacimiento paleoicnológico de Pahuén-Co (Pleistoceno tardío), provincia de Buenos Aires, Argentina. 1ª Reunión Argentina de Icnología. Asociación Paleontológica Argentina. Publicación Especial, 4, 47–57.
Aramayo, S. A., Manera De Bianco, T., Bastianelli, N. V., & Melchor, R. N. (2015). Pehuén Co: An updated taxonomic review of a late Pleistocene ichnological site. Palaeogeography, Palaeoclimatology, Palaeoecology, 439, 144–165.
Astibia, H., Del Valle, J., & Murelaga, X. (1994). Icnitas de artiodáctilos (Mammalia) del Paleógeno de Olcoz (Depresión del Ebro, Navarra). Estudios Geológicos, 50, 119–126.
Astibia, H., Pereda, X., Payros, A., Murelaga, X., Berreteaga, A., Baceta, J. I., & Badiola, A. (2007). Bird and mammal footprints from the Tertiary of Navarre (Western Pyrenees). Ichnos, 14, 175–184.
Astibia, H., Rodriguez-Tovar, F., Diaz-Martínez, J. I., Payros, A., & Ortiz, S. (2016). Trace fossils from the Middle and Upper Eocene (Bartonian–Priabonian) molasse deposits of the Pamplona Basin (Navarre, western Pyrenees): Palaeoenvironmental implications. Geological Journal, 52(2), 327–349. https://doi.org/10.1002/gj.2763
Ataabadi, M. M. (2007). Cenozoic mammal footprints of Iran and their significance. In: S. C. Lucas, Spielman, & M. G. Lockley (Eds.) Cenozoic vertebrate tracks and traces. New Mexico Museum of Natural Story and Science Bulletin, 42, 251–259.
Ataabadi, M. M., & Abassi, N. (2007). Affinities and implications of new Miocene mammal footprints from Iran. Vertebrata PalAsiatica, 45, 128–136.
Baena, J. (1981). Mapa Geológico de España E 1:50.000, 2ª serie. Hoja nº 869 (Jumilla). IGME. Secretaría de Publicaciones. Ministerio de Industria. Madrid
Bellon, H., & Brousse, R. (1977). Le magmatisme périméditerranéen occidental. Essai de synthèse. Bulletin Societé Géologique De France, 19, 469–480. https://doi.org/10.2113/gssgfbull.S7-XIX.3.469
Buck, P. V., Ghilardi, A., Peixoto, B. C. P. M., Fernandez, L. B. R., & Fernandes, M. (2016). A new tetrapod ichnotaxon from Botucatu Formation, Lower Cretaceous (Neocomian), Brazil, with comments on fossil track preservation on inclined planes and local paleoecology. Palaeogeography, Palaeoclimatology, Palaeoecology, 466, 21–37. https://doi.org/10.1016/j.palaeo.2016.11.009
Buynevich, I. V. (2015). Recent vertebrate tracks in sandy substrates and their paleoenvironmental implications: Examples from coastal Lithuania. Baltica, 28, 29–40.
Canudo, J. I. (2004). Icnitas de mamíferos en la Formación Peraltilla (Tránsito Eoceno superior-Oligoceno inferior, Paleógeno) en Abiego (Huesca). Yacimiento de Fondota. Dirección General de Patrimonio del Gobierno de Aragón. Internal report. 27 pp.
Casamiquela, R.M. (1964). Estudios icnológicos. Problemas y métodos de la icnología con aplicación al estudio de pisadas mesozoicas (Reptilia, Mammalia) de la Patagonia. Librart, Buenos Aires. 229 p.
Casanovas Cladellas, M.L., Fernández-Ortega, A., Pérez-Lorente, F., & Santafé Llopis, J.V. (1989). Huellas de dinosaurio de La Rioja. Yacimientos de Valdecevillo, La Senoba y de la Virgen del Campo. Instituto de Estudios Riojanos. Ciencias de la Tierra 12, 190 p.
Casanovas Cladellas, M. L., & Santafé Llopis, J. V. (1974). Nota sobre el hallazgo de icnitas de mamíferos en el Terciario catalán. Acta Geológica Hispánica, 9, 45–49.
Casanovas-Cladellas, M. L., & Santafe-Llopis, J. V. (1982). Icnofauna oligocena de Agramunt (Lérida, España). Acta Geológica Hispánica, 17, 113–119.
Clifton, H. E., & Hunter, R. E. (1999). Depositional and other features of the Merced Formation in sea cliff exposures south of San Francisco, California. In D. L. Wagner and S. A. Graham (eds.), Geologic field trips in Northern California: centennial meeting of the Cordilleran Section of the Geological Society of America. California Department of Conservation, Division of Mines and Geology, 119, 89–100.
Costeur, L., Balme, C., & Legal, S. (2009). Early Oligocene mammal tracks form south-eastern France. Ichnos, 16, 257–267.
D’Orazi Porchetti, S., Bertini, R. J., & Langer, M. C. (2017a). Walking, running, hopping: Analysis of gait variability and locomotor skills in Brasilichnium elusivun Leonardi, with inferences on trackmaker identification. Palaeogeography, Palaeoclimatology, Palaeoecology, 465, 14–29. https://doi.org/10.1016/j.palaeo.2016.10.009
D’Orazi Porchetti, S., Bertini, R. J., & Langer, M. C. (2017b). Proposal for ichnotaxonomic allocation of therapsid footprints from the Botucatu Formation (Brazil). Ichnos, 25, 192–207. https://doi.org/10.1080/10420940.2017.1308929
Dávalos-Álvarez, O. G., Nieto-Samaniego, A. F., Alaniz-Álvarez, S. A., Martíınez-Hernández, E., & Ramírez-Arriaga, E. (2007). Estratigrafía cenozoica de la región de Tehuacán y su relación con el sector norte de la falla de Oaxaca. Revista Mexicana De Ciencias Geológicas, 24, 197–215.
Demathieu, G. (1970). Les empreintes de pas de vertébrés du Trias de la bordure nord-est du Massif Central. Cahiers De Paléontologie, 11, 230.
Demathieu, G. (1986). Nouvelles recherches sur la vitesse des vertébrés, auteurs de traces fossiles. Geobios, 19, 327–333.
Demathieu, G., Ginsburg, L., Guerin, C., & Truc, G. (1984). Étude paléontologique, ichnologique et paléoécologique du gisement oligocène de Saignon (bassin d’Apt, Vaucluse). Bulletin Du Muséum National D’histoire Naturelle. Section c, Sciences De La Terre, Paléontologie, Géologie, Minéralogie, 6(2), 153–183.
Diaz-Martínez, I., García Fernández, S., Hernández, J. M., Murelaga, X., & Pérez-Lorente, F. (2011). Icnitas de aves y mamíferos del Mioceno temprano de La Rioja (Cuenca del Ebro, España). Ameghiniana, 48, 139–153. https://doi.org/10.5710/AMGH.v48i2(296)
Díaz-Martínez, I., Hernández, J. M., García Fernández, S., Murelaga, X., & Pérez-Lorente, F. (2012). Uvaichnites riojana: A new crane-like bird ichnotaxon from the lower Miocene of La Rioja (Ebro Basin, Spain). Proceedings of the Geologists’ Association, 123, 464–470.
Diaz-Martínez, I., Suarez-Hernando, O., Martínez-García, B., Hernández, J. M., García Fernández, S., Pérez-Lorente, F., & Murelaga, X. (2015). Early Miocene shorebird-like footprints from the Ebro Basin, La Rioja, Spain: Palaeoecological and palaeoenvironmental significance. Palaios, 30, 424–431. https://doi.org/10.2110/palo.2014.078
Díaz-Martínez, I., Suarez-Hernando, O., Martínez-García, B. M., Larrasoaña, J. C., & Murelaga, X. (2016). First bird footprints from the lower Miocene Lerín Formation, Ebro Basin. Spain. Palaeontologia Electronica, 19, 1.7A, 1–15. https://doi.org/10.26879/604
Diedrich, C. G. (2011). An overview of the ichnological and ethological studies in the Cave Bear Den in Urşilor Cave (Western Carpathians, Romania). Ichnos, 18, 9–26. https://doi.org/10.1080/10420940.2011.552578
Doyle, P., Wood, J. L., & George, G. T. (2000). The shorebird ichnofacies: An example from the Miocene of southern Spain. Geological Magazine, 137, 517–536.
Duggen, S., Hoernle, K. V., Den Bogaard, P., & Garbe-Schönberg, D. (2005). Post-collisional transition from subduction to intraplate-type magmatism in the westernmost Mediterranean: Evidence for continental-edge delamination of subcontinental lithosphere. Journal of Petrology, 46, 1155–1201.
Ellenberger, P. (1980). Sur les empreintes de pas mammifères de l’Eocene súperieur de Garriges-Ste-Eulalie (Gard). Palaeovertebrata, 9, Mémoire Jubilaire Rene Lavocat, 37–78.
Fornos, J. J., Bromley, R. G., Clemensen, L. B., & Rodriguez-Perez, A. (2002). Tracks and trackways of Myotragus balearicus Bate (Artiodactyla, Caprinae) in Pleistocene aeolianites from Mallorca (Balearic Islands, Western Mediterranea). Palaeogeography, Palaeoclimatology, Palaeoecology, 180, 277–313.
Fuster, J. M., Gastesi, P., Sagredo, J., & Fermoso, M. L. (1967). Las rocas lamproíticas del sureste de España. Estudios Geológicos, 23, 53–69.
Garcia, M.-A. (2005). Ichnologie générale de la grotte Chauvet. Bulletin De La Société Préhistorique Française, 102, 103–108. https://doi.org/10.3406/bspf.2005.13341
Garcia Raguel, M., Cuevas, I., Diaz-Martínez, I., & Pérez-Lorente, F. (2009). Fragmentos de roca con huellas de ave en el terciario de Alcanadre (La Rioja). Descripción, estructuras y problemas de identificación. Zubía, 27, 81–158.
Gatesy, S. M. (2003). Direct and indirect tracks features: What sediment did a dinosaur touch? Ichnos, 10, 91–98.
Gatesy, S. M., Middleton, K. M., Jenkins, F. A. J., & Shubin, N. H. (1999). Three-dimensional preservation of foot movements in Triassic theropod dinosaurs. Nature, 399, 141–144.
Gentry, A., Franco, C., & Bustos. D. A. (2011). Mammalian Ichnofauna from the Upper-Pleistocene Deposits of White Sands National Monument, Otero County, New Mexico. Geoscientists in the Parks. National Park Service, Denver, Colorado.
Gierlinski, G., Adach, L., & Niedzwiedzki, G. (2007). A preliminary report on mammal tracks in the Miocene of Poland. In S. G. Lucas, J. A. Spielman, & M.G. Lockley (Eds.) Cenozoic vertebrate tracks and traces, New Mexico Museum of Natural History and Science Bulletin, 42, 5–6.
Graham, R. W., Farlow, J. O., & Vandike, J. E. (1994). Tracking Ice Age felids: identification of tracks of Panthera atrox from a cave in southern Missouri, U.S.A. In K. M. Stewart, & K. L. Seymour (Eds.) Paleoecology and Palaeoenviroments of Late Cenozoic Mammals, 331–345. University of Toronto Press. https://doi.org/10.3138/9781487574154-018
Gromier, E. (1948). La vie des animaux sauvages de l’Afrique. Ed. Payot, Paris.
Gromier, E. (1949). Grands fauves d’Afrique. Ed. Amiot-Dumont, Paris.
Guerin, C., & Demathieu, G. (1993). Empreintes et pistes de rhinocerotidae (Mammalia, Perissodactyla) du gisement Pliocene terminal de Laetoli (Tanzane). Geobios, 26, 497–513.
Hall, N.T. (1965) Petrology of the Type Merced Group, San Fransisco Peninsula, California. PhD Thesis, University of California, Berleley.
Hall, N. T. (1966). Fleishacker Zoo to Mussel Rock (Merced Formation), a Plio-Pleistocene nature walk. California Division of Mines and Geology, Mineral Information Service, 19, S22-25.
Hamblin, A. H., & Foster, J. R. (2000). Ancient animal footprints and traces in the Grand Staircase Escalante National Monument, South Central Utah. Utah Geological Association, 28, 557–568.
Haubold, H. (1971). Ichnia amphibiorum et reptiliorum fossilium. In. O. Kuhn (Ed.). Handbuch der Palaoherpetologie. Gustav Fischer Verlag, 18, 124 p.
Heinz, R. (1933). Ein vorzeitlicher Tránkplatz in der Namibwüste bei Lüderitzbucht (Deutsch-Südwestafrika). Mitteilungen der Geographischen Gesellschaft in Hamburg, XLIII, 267–302.
Hernández Pacheco, F. (1929). Pistas de aves fósiles en el Oligoceno de Peralta de la Sal (Lérida). Memoria De La Real Sociedad Españoña De Historia Natural, 15, 379–383.
Herrero, E. (2010b). Nuevo yacimiento de icnitas de la Sierra de las Cabras (Jumilla, Murcia). Proceedings of the XIX jornadas de patrimonio cultural de la Región de Murcia. 39–40.
Herrero, C. (2010a). Cubierta de protección en el yacimiento de icnitas de la Sierra de las Cabras, Jumilla (Murcia). Proceedings of the XIX jornadas de patrimonio cultural de la Región de Murcia. 41–42.
Herrero, C. (1997). Las huellas fósiles de La Hoya de la Sima. El Picarcho, 93, 21–22.
Hunt, A. P., & Lucas, S. G. (2007). Cenozoic vertebrate trace fossils of North America: ichnofaunas, ichnofacies and biochronology. In S. G. Lucas, J.A. Spielman, & M. G. Lockley, (Eds.) Cenozoic vertebrate tracks and traces, New Mexico Museum of Natural History and Science Bulletin, 42,17–42.
Jabaloy Sánchez, A., Padrón-Navarta, J. A., Gómez-Pugnaire, M. T., Sánchez-Vizcaíno, V., & Garrido, C. J. (2019). Alpine orogeny: Deformation and structure in the Southern Iberian Margin (Betics s.l.). In C. Quesada & J. Oliveira (Eds.), The geology of Iberia: A geodynamic approach regional geology reviews. Cham: Springer. https://doi.org/10.1007/978-3-030-11295-0_10
Johnston, C. C. (1937). Tracks from the Pliocene of west Texas. American Midland Naturalist, 18, 147–152.
Kienapfel, K., Läbe, S., & Preuschoft, H. (2014). Do tracks yield reliable information on gaits?—Part 1: the case of horses. Fossil Record., 17, 59–67.
Koenigswald, W. V., Sander, P. M., & Walders, M. (1996). The upper Pleistocene Bottrop-Welheim (Germany). Acta Zoologica Cracoviensia, 39(1), 253–244.
Kordos, L. (1983). Üjabb adatok az ipolytarnóci alsó-miocén homokkő lábnyomaihoz. Magiar Allami Foldtani Intézet Evi Jelentése, 1985, 453–465.
Krapovickas, V. (2010). El rol de las trazas fósiles de tetrápodos en los modelos de icnofacies continentales en ambientes de climas áridos-semiáridos. Universidad de Buenos Aires.
Krapovickas, V., & Vizcaíno, S. (2016). The Cenozoic radiation of mammals. In M. G. Mángano & L. A. Buatois (Eds.), The trace-fossil record of major evolutionary events (Vol. 2, pp. 371–410). Springer.
Kraprovickas, V., Ciccioli, P. L., Mángano, M. G., Marsicano, C. A., & Limarino, C. O. (2009). Paleobiology and paleoecology of an arid-semiarid Miocene South American ichnofauna in anastomosed fluvial deposits. Palaeogeography, Paleoclimatology, Palaeoecology, 284, 129–152.
Krofel, M. (2007). Golden Jackals (Canis aureus L.) on the Peljesac Peninsula (southern Dalmatia, Croatia). Natura Croatica, 16, 201–204.
Lancis, C., & Estévez, A. (1992). Las icnitas de mamíferos del Sur de Alicante (España). Geogaceta, 12, 60–64.
Leonardi, G. (1980). On the discovery of an abundant ichno-fauna (vertebrates and invertebrates) in the Botucatu formation in Araraquara, São Paulo, Brazil. Anales Academia Brasileira De Ciencias, 52, 559–567.
Leonardi, G. (1981). Brasilichnium elusivum gen. n., sp. n.: Pistas de tetrápode mesozoico guardadas nas coleçoes do Museo Nacional do Rio de Janeiro. Anales Academia Brasileira De Ciências, 53, 793–805.
Leonardi, G. (Ed.). (1987). Glossary and Manual of Tetrapod Footprint Palaeoichnology. Ministerio de Minas e Energia.
Leonardi, G. (1994). Annotated atlas of South America tetrapod footprints (Devonian to Holocene). Ministerio de Minas e Energia.
Lockley, M. G., Reynolds, R. E., Milner, A. R. C., & Varhalmi, G. (2007b). Preliminary overview of mammal and bird tracks from the White Narrows Formation, southern Nevada. In S. G. Lucas, J. A. Spielman, & M. G. Lockley, (Eds.). Cenozoic vertebrate tracks and traces, New Mexico Museum of Natural History and Science Bulletin, 42, 91–96.
Lockley, M. G., Culver, T. S., & Wegweiser, M. (2007a). An ichnofauna of hopping rodent and arthropod trackways from the Miocene of Colorado, In S. G. Lucas,, J. A. Spielman, & M.G. Lockley (Eds.) Cenozoic vertebrate tracks and traces, New Mexico Museum of Natural History and Science Bulletin, 42, 59–66.
Lockley, M. G., Milner, & A. R. C. (2014). The ichnotaxonomy of hopping vertebrate trackways from the Cenozoic of Western USA. In M. G. Lockley, & S. G. Lucas (Eds.) Fossil footprints of Western Nort America. New Mexico Museum of Natural History and Science Bulletin, 62, 489–500.
Lockley, M. G., Ritts, B.-D., & Leonardi, G. (1999). Mammal tracks assemblages from the Early Tertiary of China, Peru, Europe and North America. Palaios, 14, 398–404.
Lockley, M., & Tedrow, A. (2009). Are Bipedopus, Semibipedopus, Lacertipus, Navahopus andBrasilichnium disticnt ichnogenera? Reevaluating Jurassic tracks from the western USA. Journal of Vertebrate Paleontology, 29(3), 136A.
López, G., Mayoral, E., Muñoz, A., Pérez, A., & Santamaréa, R. (1992). Nuevas icnitas de mamíferos carnívoros en el Mioceno Inferior del sector occidental de la Depresión del Ebro (Logroño, La Rioja). Revista De La Sociedad Geológica De España, 5, 31–51.
Lucas, S. G., & Schultz, G. E. (2007). Miocene vertebrate footprints from the Texas Panhandle. New Mexico Museum of Natural History and Science Bulletin, 42, 77–83.
Martín, J. M., Braga, J. C., Aguirre, J., & Puga-Bernabéu, A. (2009). History and evolution of the North-Betic Strait (Prebetic Zone, Betic Cordillera): A narrow, early Tortonian, tidal-dominated, Atlantic-Mediterranean marine passage. Sedimentary Geology, 216, 80–90.
McCrea, R. T., & Sarjeant, W. A. S. (2001). New ichnotaxa of bird and mammal footprints from the Lower Cretaceous (Albian) Gates Formation of Alberta. In K. Carpenter, D. H. Tanke, & M. W. Skrepnick (Eds.), Mesozoic Vertebrate Life (pp. 453–478). University of Indiana Press.
Mcneil, P., Hills, L., Kooyman, B., & Tolman, S., (2007). Significance of latest Pleistocene tracks, trackways, and trample grounds from southern Alberta. In S. G. Lucas, J. A. Spielman, & M. G. Lockley (Eds.) Cenozoic vertebrate tracks and traces, New Mexico Museum of Natural History and Science Bulletin, 42, 209–224.
Melchor, R. N., Feola, S. F., & Manera de Bianco, T. (2018). Canid paleoichnology: Taxonomy, review and producers of Canipeda from the late Pleistocene of Argentina. Ichnos, 26, 85–107. https://doi.org/10.1080/10420940.2018.1447466
Milan, J., Clemensen, L. B., Buchardt, B., & Noe-Nygaard, N. (2007). A late Holocene tracksite in the Lodbjerg dune system, Northwest Jylland, Denmark. In S. G. Lucas, J. A. Spielman, & M. G. Lockley (Eds.). Cenozoic vertebrate tracks and traces, New Mexico Museum of Natural History and Science Bulletin, 42, 241–250
Morales, J., Senut, B., & Pickford, M. (2011). Crocuta dietrichi from Meob, Namibia: Implications for the age of the Tsondab Sandstone in the coastal part of the Namib Desert. Estudios Geológicos, 67, 207–215. https://doi.org/10.3989/egeol.40463.184
Morgan, G. S., & Williamson, T. E. (2007). Middle Miocene (Late Barstovian) mammal and bird tracs from the Benavidez Ranch local fauna. Zia Formation, Albuquerque basin, Sandoval County, New Mexico. In S. G. Lucas, J. A. Spielman, & M. G. Lockley (Eds.) Cenozoic vertebrate tracks and traces, Bulletin of New Mexico Museum of Natural History and Science Bulletin, 42, 319–330.
Murelaga, X., Astibia, H., Baceta, J. I., Almar, Y., Beamud, B., & Larrascaña, J. C. (2007). Fósiles de pisadas de aves en el Oligoceno de Etaio (Navarra, Cuenca del Ebro). Geogaceta, 41, 139–142.
Murelaga, X., Baceta, J. I., Astibia, H., Badiola, A., & Pereda, X. (2000). Icnitas de perisodáctilos en el Olicoceno de Navarra: Posición estratigráfica y sistemática. Geogaceta, 27, 15–17.
Mustoe, G. E. (2002). Eocene bird, reptile, and mammal tracks from the Chuckanut Formation, Northwest Washington. Palaios, 17(4), 403–413. https://doi.org/10.1669/0883-1351(2002)017%3c0403:EBRAMT%3e2.0.CO;2
Neto, C. (2009). Vertebrate tracksites from the Mid-Late Pleistocene eolianites of Portugal: The first record of elephants tracks in Europe. Geological Quarterly, 53, 407–414.
Neto, C., & Figueiredo, & S., Belo, J. (2016). Vertebrate tracks and trackways from the Pleistocene eolianites of SW Portugal. Comunicaçoes Geológicas, 103(1), 101–116.
Panin, N., & Avram, E. (1962). Noe urme de pas de vertebrate in Miocenul Subcarpatilor Ruminestkya. Studie Si Cercetari De Géologie, Géophyzica, Si Géografie, Serie De Géologie, 7, 455–484.
Payros, A., Astibia, H., Cearreta, A., Pereda-Suverbiola, X., Murelaga, X., & Badiola, A. (2000). The Upper Eocene South Pyrenean coastal deposits (Liedena Sandstone, Navarre): Sedimentary facies, benthic foraminifera and avian ichnology. Facies, 42, 107–132.
Peabody, F.E., (1959). Trackways of living and fossil salamanders. University of California Publications in Zoology 63, 71 p.
Pérez-Lorente, F. (2001). Paleoicnología. Los dinosaurios y sus huellas en La Rioja. Fundación Patrimonio Paleontológico de La Rioja.
Pérez-Lorente, F. (2015). Dinosaur footprints and trackways of La Rioja. In J. O. Farlow (Ed.), Life of the past. Indiana Univerity Press.
Pérez-Lorente, F., Herrero, C., Herrero, E., & Montoya, P. (2009). Paracamelichnum jumillensis n.ichnogen. n.ichnosp., Upper Miocene Camelidae Ichnites from the Hoya de la Sima site (Murcia, Spain). Ichnos, 16, 208–219.
Pérez-Lorente, F., & Herrrero-Gascón, J. (2007). El movimiento de un dinosaurio deducido de una rastrillada terópoda con estructuras de inmersión de los pies en el barro y de arrastre de cola (Formación Villar del Arzobispo. Galve, Teruel, España). Revista Española De Paleontología, 22, 157–174. https://doi.org/10.7203/sjp.22.2.20421
Perez-Lorente, F., Rodriguez, T., Mancheño, M. A., Serrano, F., & Romero, M. (1997). Pisadas fósiles de mamíferos en el Mioceno Superior de la Hoja de la Sima (Jumilla, Murcia, España). Memorias De Arqueología, 12, 15–28.
Perez-Lorente, F., Serrano, F., Rodríguez, T., Mancheño, M. A., & Romero, M. (1999). Pisadas fósiles de mamíferos en el mioceno superior de La Hoya de la Sima (Jumilla, Murcia. España). Revista Española De Paleontología, 14, 257–267.
Prats, M., & López, G. (1995). Sintesi de la icnofauna del sinclinal de Agramunt, entre el meridiá d’Agramunt i el riu Segre (Prov. de Lleida). Paleontologia i Evolució, 28–29, 247–267.
Quintana, J., & Arnau, P. (2004). Descripció dels rastres i les petjades d’Hypnomys Bate, 1918 (Mammalia: Gliridae) de la cova de sa Duna (Alaior, Menorca). Endins, 26, 7–14.
Rabal, R., & Diaz-Martínez, I. (2010). Primeras icnitas de mamífero carnívoro en el Paleógeno de Aragón (España). Cidaris, 30, 259–264.
Rainforth, E. C., & Lockley, M. G. (1996). Tracks of diminutive dinosaurs and hopping mammals from the Jurassic of north and south America. In: M. Morales (Ed.). Continental Jurassic Symposium Museum of Northern Arizona Bulletin, 60, 265–269.
Rajkumar, H. S., & Klein, H. (2014). First perissodactyl footprints from Flysch deposits of the Barail Group (Lower Oligocene) of Manipur, India. Journal of Earth System Science, 123, 413–420. https://doi.org/10.1007/s12040-013-0396-6
Remeika, P. (2001). The Fish Creek Canyon ichnofauna: a Pliocene (Blancan) vertebrate footprints assemblage from Anza-Borrego Desert State Park, California. In V. Santucci & L. Maclelland (Eds.) Proceedings of the 6th fossil resource conference. 55–75. Technical Report, National Park Service, Geologic Resources Division.
Remeika, A. P. (1999). Identification, stratigraphy and age of Neogene vertebrate footprints from the Vallecito-Fish Creek Basin, Anza-Borrego Desert State Park, California. San Bernardino County Museum Association Quarterly, 46, 37–45.
Requeta, L. E., Hernández Medrano, N., & Pérez-Lorente, F. (2006–07). La Pellejera: Descripción y aportaciones. Heterocronía y variabilidad de un yacimiento con huellas de dinosaurio de La Rioja (España). Zubía Monográfico, 18–19, 21–114.
Reynolds R.E. (2001). Wolves of Shoshone, Southern Death Valley. In R.E. Reynolds (Eds.). The changing face of the east Mojave desert. 2001. Desert symposium. California State University, Desert Studies Consortium, Abstracts, 58–60
Reynolds, R. E. (2006). Horse hoof prints in the fossil record. In R.E. Reynolds (Ecd.). Making tracks across the Southwest. 2006 Desert symposium field guide and abstracts from proceedings. California State University, Desert Studies Consortium & LSA Associates, Inc. 25–28.
Reynolds, R. E., & Milner, A. R. C. (2007). Preliminary description of mamma trackways from te Middle Miocene (Late Barstovian Nalma) Enterprise reservoir sediments in southwestern Utah. In S. G. Lucas, J. A. Spielman, & M. G. Lockley (Eds.). Cenozoic vertebrate tracks and traces, of New Mexico Museum of Natural History and Science Bulletin, 42, 261–266
Reynolds, R. E., & Milner, A. R. C. (2012). Early Neogene cat tracks from California and Utah. In R. E. Reynolds (Ed.) Searching for the Pliocene: Southern Exposures. 2012 Desert symposium, California State University.152–159.
Reynolds, R. E., Roeder, M. A., Wake, T. A., & Smith, G. R. (2016). Freshwater fauna and flora from Bouse basal marl in Little Chemehuevi Wash. In R. E. Reynolds (Ed.) Investigations along the Lower Colorado River. Desert Symposium Field Guide and Proceedings California State University 112–120.
Rezendes, P. (1999), Tracking and the art of seeing: how read animal racks and sign. 2nd Edition. Collins Reference. 336 p.
Richarz, K. (2002). Steinbachs Naturführer. Mosaik Verlag.
Roberts, D. L. (2008). Last Interglacial Hominid and Associated Vertebrate Fossil Trackways in Coastal Eolianites, South Africa. Ichnos, 15, 190–207. https://doi.org/10.1080/10420940802470482
Roberts, G. (2009). Ephemeral, Subfossil Mammalian, Avian and Hominid Footprints within Flandrian Sediment Exposures at Formby Point, Sefton Coast, North West England. Ichnos, 16, 33–48. https://doi.org/10.1080/10420940802470730
Roca, E., Sans, M., & Koyi, H. (2006). Polyphase deformation of diapiric areas in models and in the eastern Prebetics (Spain). American Association of Petroleum Geologists Bulletin, 90, 115–136.
Rodriguez De La Rosa, R.A., & Guzmán,J.R. (2012). Huellas de aves y de mamíferos del Neógeno de Jalisco, México. Paleontología mexicana, LXII, 147–158.
Rossi, C., Vilas, L., & Arias, C. (2015). The Messinian marine to nonmarine gypsums of Jumilla (Northern Betic Cordillera, SE Spain): Isotopic and Sr concentration constraints on the origin of parent brines. Sedimentary Geology, 328, 96–114.
Rubinat, M., Roca, E., Escalas, M., Queralt, P., Ferrer, O., & Ledo, J. J. (2013). The influence of basement structure on the evolution of the Bicorb-Quesa Diapir (eastern Betics Iberian Peninsula): Contractive thin-skinned deformation above a pre-existing extensional basement fault. International Journal of Earth Sciences, 102, 25–41. https://doi.org/10.1007/s00531-012-0789-9
Santamaria, R., Lopez, G., & Casanovas, M. L. (1989–90). Nuevos yacimientos con ichnitas de mamíferos del Oligoceno en los alrededores de Agramunt (Lérida, España). Paleontologia y evolución. 23, 141–152.
Santucci, V. L., Tweet, J., Bustos, D., Nyborg, T., & Hunt, A. P. (2014). An inventory of Cenozoic fossil vertebrate tracks and burrows in National Park Service areas. In M. G. Lockley & S. G. Lucas (Eds.) Fossil footprints of western North America. New Mexico Museum of Natural History and Science Bulletin, 62, 469–488.
Sarjeant, W. A. S., & Reynolds, R. E. (1999). Camel and horse footprints from the Miocene of California. In R.E. Reynolds ed. 1999 Desert Research Symposium. Abstracts from Proceedings. San Bernardino County Museum Association Quarterly, 46, 3–18
Sarjeant, W. A. S., & Reynolds, R. E. (2001). Bird footprints from the Miocene of California. In R. E. Reynolds (Ed.). The changing face of the Mohave desert. Abstracts of the 2001 Desert Symposium. California State University. 21–40.
Sarjeant, W. A. S, Reynolds, R. E., & Kissell-Jones, M. M. (2002). Fossil creodont and carnivore footprints from California, Nevada, and Wyoming. In R.E. Reynolds (Ed.) Between the basins: exploring the Western Mojave and southern Basin and Range Province. Abstracts from the 2002 Desert Symposium. California State University. 37–50.
Sarjeant, W. A. S., & Langston, W. (1994). Vertebrate footprints and invertebrate traces from the Chadronian (Late Eocene) of West Texas. Texas Memorial Museum Bulletin, 36, 1–86.
Scrivner, P. J., & Bottjer, D. J. (1986). Neogene avian and mammalian tracks from Death Valley National Monument, California: Their context, clasification and preservation. Palaeogeography, Palaeoclimatology, Palaeoecology, 57, 285–331.
Solecki, R. S., & Solecki, R. L. (1966). New data from Yabroud, Syria, Preliminary report of the Columbia University archaeological investigations. Annales Archéologiques Árabes Syriennes, 16, 121–153.
Stuart, C., & Stuart, T. (2006). Field guide to the larger mammals of Africa. Struik publishers.
Thenius, E. (1967). Saugetierfährten aus den Rohrbacher Konglomerat (Pliozän) von Neiderösterreich. Annalen Der Naturhistorischen Museums in Wien, A, 71, 363–379.
Thompson, M. E., White, R. S., & Morgan, G. S. (2007). Pace versus trot: can medium speed gait be determined from fossil trackways? In S. G. Lucas, J. A. Spielman, & M. G. Lockley (Eds.) Cenozoic vertebrate tracks and traces, New Mexico Museum of Natural History and Science Bulletin, 42, 309–314.
Thulborn, A. (1990). Dinosaur tracks. Chapman and Hall.
Thulborn, R. A., & Wade, M. (1989). A footprint as a history of movement. In D. D. Gillette & M. G. Lockley (Eds.), Dinosaurs Tracks and Traces (pp. 51–56). Cambridge Univ.
Turner, S. P., Platt, J. P., George, R. M. M., Kelley, S. P., Pearson, D. G., & Nowell, G. M. (1999). Magmatism associated with orogenic collapse of the Betic-Alboran domain, SE Spain. Journal of Petrology, 40, 1011–1036.
Vera, J. A. (2000). El Terciario de la Cordillera Bética: Estado actual de conocimientos. Revista De La Sociedad Geológica De España, 12, 345–373.
Vialov, O. S. (1965). Stratigrafiya neogenovix molass Predkarpatskogo progiba, Kiev. Naukova Dunka. pt.K.
Vialov, O. S. (1966). Sledy zhiznedeyatel’nosti organizmov I ikh paleotologicheskoe znachenie. Akademii Nauk Ukranian.
Vilas, L., Arias, C., Rozycki, A., Herrero, C., & Martínez-Abellán, R. (2006). El yacimiento de icnitas de vertebrados de la Hoya de la Sima (Mioceno terminal). Jumilla, Murcia, España. De Re Metallica, 6–7, 1–6.
Williamson, T. E., & Lucas, S. G. (1996). Mammal footprints from the Miocene-Pliocene Ogalla Formation, eastern New Mexico. New Mexico Geology, 18, 1–5.
Acknowledgements
We are very grateful to the Jerónimo Molina Museum of Ethnography and Natural Sciences of Jumilla town and the Jumilla City Council for the attention received during our stay and for the help provided to carry out the work. Also we greatefull to the anonimous reviewers for their corrections and suggestions.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. There has been no funding for this work nor is it included in a funded project.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visithttp://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Herrero, C., Herrero, E., Martín-Chivelet, J. et al. Vertebrate ichnofauna from Sierra de las Cabras tracksite (Late Miocene, Jumilla, SE Spain). Mammalian ichnofauna. J Iber Geol 48, 241–279 (2022). https://doi.org/10.1007/s41513-022-00192-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41513-022-00192-5