
Abstract
Biological nitrogen fixation (BNF) can help replenish available nitrogen (N) in cropland and reduce the use of chemical N fertilizers, with diazotrophs playing an important role. However, the response of diazotroph community and BNF activity in biochar amendment soil, especially in the deep soil horizon, are poorly understood. In this study, soil samples were collected from topsoil (0–20 cm) and subsoil (20–40 cm) in the field experiment (established in 2013) comprising treatments with no chemical fertilizer (CK), chemical fertilizer (NPK), biochar (BC), and biochar plus chemical fertilizers (BNPK). Here, we investigated the diazotroph community using real-time PCR and high-throughput sequencing of the nifH gene, and assessed the soil N2 fixation rate (RNfix) using acetylene reduction assay (ARA). Results showed that in the topsoil, the treatments with biochar significantly increased nifH gene copies and RNfix, which was consistent with the increased soil organic matter (SOM), total carbon-to-nitrogen ratio (C/N), dissolved organic carbon (DOC) and pH. In the subsoil, applying chemical fertilizers (NPK) strongly decreased RNfix, but had no effect on diazotroph abundance; in contrast, biochar application (BC) had no effect on RNfix, but suppressed the growth of bacteria and diazotrophs while increasing the abundance of Rhizobiales order. Diazotroph and bacterial gene copies were significantly and positively correlated in both top- and sub-soil, and they were mainly influenced by SOM and total nitrogen (TN). In addition, soil nitrate nitrogen (NO3−–N) was the major factor in shaping the vertical stratification of diazotroph community structure. Although nifH gene abundance was significantly correlated with RNfix in the topsoil, the structure equation modeling (SEM) showed the highest correlation between diazotroph community structure and RNfix. Hence, we suggested that soil carbon and nitrogen sources were the key factors correlated with changes in the vertical pattern of diazotroph abundance. Biochar induced the dominant diazotroph community succession and increased soil carbon content and pH, which contributed to the BNF activity. Changes in the BNF activity were driven by the variation in diazotroph community structure.
Highlights
-
Biochar significantly increased 16S rRNA and nifH gene copies in the topsoil.
-
Soil carbon and nitrogen sources were the key factors correlated with the diazotroph abundance rather than its specific functional traits.
-
Biochar induced the dominant diazotroph community succession and increased Rhizobiales, which contributed to the BNF activity.
-
Changes in the BNF activity were driven by the variation in diazotroph community structure.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Nitrogen (N) is an essential macronutrient and a limiting resource for crop yield and quality (Liu et al. 2020). To meet the demand for food for a growing population, countries face the challenges of maintaining agricultural productivity and environmental safety in applying chemical fertilizers (Chen et al. 2019; Hu et al. 2019), especially regarding the overuse of N fertilizers (Liu et al. 2013), such as soil acidification (Lin et al. 2018), N leaching (Liu et al. 2020) and ammonia volatilization (Verhoeven and Six 2014). Rational use of N implies not only the utilization of chemical N fertilizers, but also a consideration of the alternative approach of supplying plant-available N from the N-rich atmosphere via biological nitrogen fixation (BNF) (Gul and Whalen 2016). The previous report showed BNF is responsible for supplying 2.95 million tonnes of N annually to pulse crops and about 18.5 million tonnes of N annually to oilseed legumes in the world via the action of free-living and symbiotic diazotrophs (Gul and Whalen 2016). Biochar, as an organic carbon (C)-rich soil amendment (Lehmann et al. 2011), can serve to buffer nutrient availability and retain moisture (Hagemann et al. 2017; Li et al. 2019a), while mediating soil microbial activity, including promoting BNF processes (Mia et al. 2014; Gul and Whalen 2016), which could contribute to reducing N fertilizer application (Chen et al. 2019).
Diazotrophs act as contributors to BNF, changing the atmospheric N2 into ammonium via nitrogenase (Kumar et al. 2020). The nifH gene, encoding a subunit of nitrogenase, is highly conserved across the bacterial and archaeal taxa and thus is widely used as a molecular marker to investigate the size and composition of diazotroph communities (Lin et al. 2018; Liu et al. 2019a; Wang et al. 2016). Lin et al. (2018) studied the characteristics of diazotroph communities in a soil amendment experiment with different additives applied to topsoil and concluded that nifH gene abundance could indirectly indicate BNF activity (Reardon et al. 2014). However, the results of Zhang et al. (2020) contradict this, as they found a poor relationship between soil nitrogenase activity and nifH gene abundance in pristine peatlands. Moreover, some studies are ambiguous; for example, Tang et al. (2018) and Wang et al. (2016) revealed significant decreases in the nifH abundance in forest and wheat soils, respectively. Since high concentration of oxygen inactivates nitrogenase (Lin et al. 2018), anaerobic conditions, such as in subsoil, usually favor the activity of diazotrophs (Kumar et al. 2020; Limmer and Drake 1996), but the decline in abundance of the diazotrophs with soil depth implies that the relationship between diazotroph growth and the BNF activity would not be strong. Nevertheless, the variable diazotroph community composition may be largely responsible for this, such as the high exogenous energy dependence of heterotrophic diazotroph growth (Weber and King 2012) and the direct association of rhizosphere diazotrophs with plant roots (Garrido-Oter et al. 2018). Therefore, the exploration of diazotroph community structure would be fundamental to the investigation of BNF activity variation.
Microbial community is innately sensitive to environmental changes (Liu et al. 2019c), so it is important to understand how fertilization modifies the soil environment and induces changes in the diazotroph community and BNF. The well-developed porous structure and a high degree of surface aromatization form the basis of biochar capacity to improve soils and regulate soil microbial communities (Chen et al. 2019; Lehmann et al. 2011). Biochar enables uptake of nutrients from chemical fertilizers, organic matter mineralization, and atmospheric deposition to supply microbial demand. And, it has been reported that biochar application could increase the abundance of soil diazotrophs (Oram et al. 2014), enhance nitrogenase activity (Mia et al. 2014), change the structure of diazotroph community (Liu et al. 2019c) and induce some microbial taxa succession, such as the Burkholderiales (Chen et al. 2018). Chemical fertilization usually has a negative effect on BNF processes, such as decreasing the abundance of nifH gene (Wang et al. 2017) and the BNF rate (Tang et al. 2017). Generally, these results are mostly attributed to the multiple secondary effects arising from variability in the soil parameters, such as C (energy) and N contents (Weber and King 2012; Zhang et al. 2020), nitrate nitrogen (NO3−–N), ammonium nitrogen (NH4+-N), and available phosphorus (AP) and potassium (AK) down the soil depth (Lin et al. 2018; Mia et al. 2014; Oram et al. 2014), soil pH (Lin et al. 2018) and other factors (e.g. salt ions and micronutrients) (Yang et al. 2016), and soil physical properties (e.g. soil moisture and oxygen partial pressure) (Wang et al. 2016). In addition, different diazotroph communities, e.g. free-living and symbiotic diazotrophs, responded differentially to specific soil variables (Che et al. 2018). Although many factors may influence diazotroph communities, the specific responses of diazotroph communities to biochar and chemical fertilization, and the ecological drivers of BNF variation, remain poorly understood.
The effects of biochar additions on microbiota in topsoil have been reported widely (Gul and Whalen 2016; Chen et al. 2019). However, the strong effects of soil depth on the diazotroph community and activity have been ignored (Liu et al. 2020), leaving the response of diazotroph communities in arable subsoil is unclear. It could be anticipated that the microbiota in different soil layers would not show a consistent response to fertilization practices (Azeem et al. 2021; Li et al. 2019a; Gul and Whalen 2016; Hu et al. 2019). As an important commercial oil crop, peanut (Arachis hypogaea L) could form the symbiosis with various rhizobia (Furlan et al. 2017) and also yield considerable amounts of diazotrophs into soil, thus constituting an important source of N supply. To investigate the responses of biochar, chemical fertilizer and their combined application on soil properties, diazotroph communities and activity in bulk soil, we analyzed topsoil (0–20 cm) and subsoil (20–40 cm) samples collected in a peanut field trial established in May 2013 using real-time quantitative PCR, Illumina Miseq sequencing and the acetylene (C2H2) reduction assay (ARA). Specifically, we hypothesized that, (i) the size, structure and activity of the diazotroph community would respond differently to soil depths as well as treatments, such as biochar, chemical fertilizer and their combination; and (ii) the relationship between diazotroph activity and diazotroph community composition would be closer than that with its abundance.
2 Materials and methods
2.1 Experimental site and design
The experiment was set up at the National Peanut Industry Technology System base in Shenyang Agricultural University (41°49 'N and 123°34 'E), Liaoning Province, China, in May 2013. The tested soil is classified as Alfisol derived from quaternary loess. The experimental site climate is detailed in Chen et al. (2020). The field trial comprised four treatments in a randomized block design with three replicates. Each replicate plot covered an area of 2 m2 and was separated by a 20 cm wide concrete wall embedded 2 m into the soil. The four treatments were: (i) CK, no fertilizer; (ii) NPK, NPK chemical fertilizer only; (iii) BC, biochar only; (iv) BNPK, BC + NPK chemical fertilizer. The chemical fertilizers applied were urea, calcium superphosphate and potassium sulphate. The biochar was produced from corn straw by pyrolysis at 450℃ under limited oxygen and was supplied by Liaoning Biochar Engineering & Technology Research Center (Shenyang, Liaoning, China). In the BC and BNPK treatments, 1500 kg ha−1 of biochar was applied annually. Biochar contained 1.25% N, 0.88% P2O5, 2.70% K2O and 41.99% C, at pH 10.06 (measured in 2018). Both the NPK and BNPK treatments were supplied with the same rates of 225 kg N ha−1 yr−1, 112.5 kg P2O5 ha−1 yr−1 and 112.5 kg K2O ha−1 yr−1, and the BNPK treatment was supplemented with the sub-optimal rates of N, P, and K through the chemical fertilizers.
In early May (2013–2018), biochar was evenly spread onto the soil surface and immediately tilled into the soil for ridging; a furrow depth of 10–15 cm was opened in the middle of the wide-ridge (70 cm wide) for fertilization prior to sowing. Peanut cv Fuhua No. 12 was sown by placing two seeds per hole on the both sides of the ridge to give 7 cm plant-to-plant spacing and 30 cm row-to-row spacing, and was harvested in September. All treatments had the same field management.
2.2 Soil sampling and analyses
Soil sampling was conducted on September 24th for the control treatment and October 1st for the other treatments, 2018. For the experimental treatments, seven 1.5 cm diameter soil cores (randomly collected in each plot) were segmented into four 10 cm layers down to 40 cm layer. We mixed the top two layers and the two bottom layers to get the topsoil (0–20 cm) and subsoil (20–40 cm). The collected soil was sieved (2 mm); one portion of the mixed soil samples was air-dried, another portion was stored at 4 ℃ for chemical measurements, and the third portion was stored at −80 ℃ and used for DNA extraction and the soil N2 fixation rate (RNfix) assay.
Fresh soil samples (10 g) were extracted with 50 mL of 2 M KCl, and NH4+–N and NO3−–N were measured using a continuous flow analyzer (SEAL AutoAnalyzer3 HR, Germany). Separated fresh soil samples (10 g) were extracted with 50 mL of 0.5 M K2SO4 for soil dissolved organic carbon (DOC) and nitrogen (DON) determination using a TOC analyzer (Multi N/C® 3100, Analytik Jena AG, Germany). Air-dried soil samples were extracted for AP and AK with 0.5 M NaHCO3 and 1.0 M ammonium acetate, respectively, and measured by colorimetry (Epoch™, BioTek, USA) and flame photometry (M410, Sherwood, England), respectively (Bao 2000). pH was measured using a glass electrode in a 1:5 soil to water ratio. SOM and TN were determined according to Chen et al. (2020) and measured using an elemental analyzer (Vario ELIII, Elementar, Germany).
2.3 The N2 fixation rate assay
The RNfix was determined by the ARA (Liu et al. 2019c). In the presence of C2H2, nitrogenase enzyme produces ethylene (C2H4), so the measurement of C2H4 production is a measure of nitrogenase activity (Belnap 2002). When soil samples were transported to laboratory, 10 g of fresh soil was placed into a 110 mL penicillin bottle sealed with a butyl rubber stopper, and 10% of the headspace (11 mL) was substituted with pure C2H2 after a 24 h pre-incubation. All the samples were incubated at 25 ℃ in dark for 48 h (Liu et al. 2019c). The C2H4 concentrations were determined by gas chromatography (Shimadzu GC 2010PLUS, Japan). The RNfix was reported in nmol C2H4 kg−1 dry weight soil (dws) h−1.
2.4 Soil DNA extraction and real-time PCR
0.25 g of frozen soil was used for extracting total DNA using a PowerSoil® DNA Isolation kit (MoBio Laboratories, Inc., USA). The DNA concentration and purity were determined using a NanoDrop 2000 UV–vis spectrophotometer (Thermo Scientific, Wilmington, USA), and DNA quality was checked by 1% agarose gel electrophoresis.
Two genes, nifH and 16S rRNA, were subjected to real-time quantitative PCR (qPCR) in a Rotor-Gene Q real-time PCR system (Qiagen, Germany). The primer sets nifH-F/nifH-R (5'-AAA GGY GGW ATC GGY AAR TCC ACC AC-3’/5'- TTG TTS GCS GCR TAC ATS GCC ATC AT) (Rosch et al. 2002) and 338F /806R (5'- ACT CCT ACG GGA GGC AGC AG -3'/5'- GGA CTA CHV GGG TWT CTA AT-3') (Xu et al. 2016) were used to amplify nifH and 16S rRNA, respectively. The qPCR methodology was following Wang et al. (2016), as detailed in supporting information Table S1. Melting curves and agarose gel electrophoresis were used to examine the specificity of the amplified products.
2.5 Illumina MiSeq sequencing and bioinformatics analysis
The diazotroph community structure was determined via high-throughput sequencing of nifH gene. The amplicons were recovered from a 2% agarose gel, further purified using an AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA), and quantified using QuantiFluor™-ST (Promega, USA) according to the manufacturer’s protocol. Purified products were pooled in equimolar amounts and paired-end sequenced (2 × 300) on an Illumina MiSeq platform (Illumina, San Diego, CA, USA) according to the standard protocols. The raw sequence data were submitted to the NCBI Sequence Read Archive (https://www.ncbi.nlm.nih.gov/sra/) under BioProject PRJNA738640.
The resulting fastq files were demultiplexed, quality-filtered by Trimmomatic and merged by FLASH using the standard procedures. In brief, the reads were truncated at any site receiving an average quality score < 20 over a 50-bp sliding window. Primers were matched allowing 2 nucleotide mismatches, and reads containing ambiguous bases were removed. Sequences with overlaps longer than 10 bp were merged according to their overlapping sequences. Operational taxonomic units (OTUs) were clustered with a 90% similarity cutoff using UPARSE (version 7.1 http://drive5.com/uparse/), and chimeric sequences were identified and removed using UCHIME (Lin et al. 2018). The taxonomy of each nifH gene sequence was analyzed by the RDP Classifier algorithm (http://rdp.cme.msu.edu/) against the FGR database (http://fungene.cme.msu.edu/) using confidence threshold of 60%. The normalized OTU table was used in the downstream analysis.
2.6 Statistical analysis
Data were statistically analyzed using the SPSS 19.0 software package (IBM, Armonk, NY, USA) for Windows. Differences in all the data were assessed by one-way ANOVA followed by the Tukey’s HSD test at P ≤ 0.05.Multivariate ANOVA was used to identify the effects of biochar, chemical fertilizer, soil depth, and their interactions. The relationships between nifH and 16S rRNA genes abundance (log-transformed), RNfix, diazotroph community compositions and diversity indices, and soil properties were subjected to the Spearman's correlation analysis. Data were normalized when needed to meet the analysis assumptions. An OTU-based normalized table was used to perform structural analysis, when not specifically stated.
R software (Version 4.0.0) was employed to perform the following analyses. The relative influences of soil properties on nifH and 16S rRNA genes abundance were quantitatively estimated by conducting aggregated boosted tree analysis (ABT) using the “gbm” package with 5000 trees for boosting, tenfold cross-validation, and three-way interactions (Lin et al. 2018). Correlation among the beta diversity of diazotroph community structure was calculated using the Bray–Curtis distance matrix via principal coordinate analysis (PCoA) in the “vegan” package. The significance of the diazotroph community difference among samples was tested by the analysis of similarities (ANOSIM) based on 9999 permutations (Liu et al. 2020). The relationships between the diazotroph community (as well as nifH gene copies, RNfix) and soil properties were determined by Redundancy Discrimination Analysis (RDA) and the Mantel test (Bai et al. 2015; Lin et al. 2018). The analysis of multivariate regression tree (MRT) was used to describe the relationships between the diazotroph community and soil properties using the “mvpart” and “MVPARTwrap” packages. Structural equation modeling (SEM), using the “lavaan” package, was performed to explore the interactive effects of diazotroph abundance and community and soil variables on RNfix based on the vertical distribution pattern. To simplify the SEM model, we reduced the dimensionality of soil variables to remove potential collinearity. The first axis score of PCoA, which represents diazotroph community, was incorporated into the model (Zhang et al. 2020). Goodness-of-fit for the model was examined by the χ2 test (P > 0.05) and root mean squared error of approximation (RMSEA < 0.05); the models with satisfactory fits were compared using the Akaike's Information Criterion (AIC) (Zhang et al. 2020).
3 Results
3.1 Soil properties
According to two-way ANOVAs (Table 1), biochar (BC) had a significant positive effect on SOM, pH, and C/N ratio in both soil layers (increasing them by 8.6–30.6%, 4.1–9.2% and 11.9–31.0%, respectively) and DOC and DON in subsoil (P < 0.05) (increases of 8.0% and 6.0%, respectively) compared with the CK; in contrast, chemical fertilizer (NPK) negatively affected these properties (P < 0.05). The NPK significantly increased AP (149.8–381.0%) and AK (8.0–54.0%). There was a significant negative effect of the chemical fertilizer × biochar interaction (NPK × BC, i.e. BNPK) on NO3−–N in two soil layers, as well as a positive effect on SOM, C/N ratio, DON, AP, and AK in subsoil (P < 0.05), with the BNPK having the highest TN (0.79 g kg−1) in 0–20 cm. When soil depth (Depth) was included in a multivariate ANOVA, the depth significantly decreased all soil parameters (P < 0.05) except NH4+-N (Table 1).
3.2 16S rRNA and nifH genes abundance
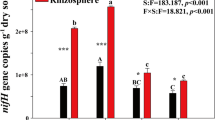
The abundance of 16S rRNA and nifH genes ranged from 1.2 × 109 to 4.6 × 109 copies g−1 and 6.6 × 105 to 2.6 × 106 copies g−1 in dws, respectively (Fig. 1a). Gene copies in the topsoil were higher than in the subsoil in all the treatments (P < 0.05). Multivariate ANOVAs revealed that except for the NPK, the other factors, such as BC, depth and their interactions had a significant effect on the abundance of these two genes (P < 0.05, Table 2). In the topsoil, the BC had more 16S rRNA and nifH gene copies (by 45% and 128%, respectively) and the BNPK had more nifH gene copies (by 71%) compared with CK. However, 16S rRNA and nifH gene copies in the BC significantly declined in the subsoil, with 48% and 27% decrease, respectively (P < 0.1).

Abundance (copy numbers g−1 dry weight soil) of 16S rRNA and nifH genes in topsoil and subsoil under different treatments (a). Bars represent ± standard errors of the means (n = 3). Significant differences (P < 0.05) among the treatments within same soil layer are indicated by different uppercase letters and the two soil layers within the same treatment by different lowercase letters. The asterisks denote the significance level: * P < 0.05; ** P < 0.01; *** P < 0.001. Relationships between the abundance of the 16S rRNA and nifH genes (b). CK: no fertilizer; NPK: chemical fertilizer; BC: biochar; BNPK: biochar plus chemical fertilizer
Significant positive relationships between 16S rRNA and nifH gene copies were detected in both topsoil (R2 = 0.66, P ≤ 0.001) and subsoil (R2 = 0.84, P ≤ 0.001) (Fig. 1b). The relative influence of the vertical distribution of the soil properties on 16S rRNA and nifH gene copies was determined by the ABT model (Fig. 2), which showed TN (accounting for 25.8% of the variation) and SOM (27.4%) as the major factors determining the abundance of 16S rRNA and nifH genes, respectively. Furthermore, Spearman’s correlations were performed to understand a differential response of these genes to soil properties in each soil layer. The results revealed that SOM, C/N and pH had a positive effect on 16S rRNA and nifH gene copies in the topsoil (P < 0.05), whereas TN and NO3−–N had a positive effect on these genes in the subsoil (Table S2).

Relative influence of soil chemical properties on the 16S rRNA and nifH gene abundance estimated by thy ABT models (n = 24). SOM: soil organic matter; TN: total nitrogen; C/N: the ratio of total carbon to nitrogen; DOC and DON: dissolved organic carbon and nitrogen; NH4+–N: ammonium nitrogen; NO3−–N: nitrate nitrogen; AP and AK: available phosphorus and potassium
3.3 The N2 fixation rate
As showed in Fig. 3a, RNfix ranged from 0.34 to 9.73 nmol C2H4 kg−1 dws h−1. Both the BC and NPK had a significant effect on RNfix (P < 0.05, Table 2). The BC and BNPK had the higher RNfix in the topsoil (by 52.2% and 38.4%, respectively) compared with the CK (P < 0.05), and the NPK had the lowest RNfix in the subsoil (P < 0.05). There was a significant positive relationship between RNfix and nifH gene copies in the topsoil (R2 = 0.95, P < 0.001), whereas a weak linear relation (R2 = 0.04, P = 0.257) was observed in the subsoil (Fig. 3b). In addition, RNfix was significantly and positively correlated with SOM, C/N and pH in both soil layers, and was positively related to DOC, DON and NH4+–N, and negatively related to AP (P < 0.05, Table S2).

RNfix (nmol C2H2 kg−1 dry weight soil h−1) in topsoil and subsoil under different treatments (a). Bars represent ± standard errors of the means (n = 3). Significant differences (P < 0.05) among the treatments within same soil layer are indicated by different uppercase letters and the two soil layers within the same treatment by different lowercase letters. The asterisks denote the significance level: * P < 0.05; ** P < 0.01; *** P < 0.001. Relationships between the nifH gene abundance and RNfix (b). CK: no fertilizer; NPK: chemical fertilizer; BC: biochar; BNPK: biochar plus chemical fertilizer
3.4 Diazotroph community structure
After quality filtering, a total of 408,241 high-quality sequences were obtained from 24 samples ranging from 13,073 to 22,512 in each sample, and the number of observed OTUs (based on the normalized OTU table, the same below) among the treatments ranged from 235 to 556 (Table 3). According to Multivariate ANOVAs, soil depth significantly affected the OTUs and Shannon index, increasing with depth; in contrast, the BC had no significant effect on the alpha- diversity indices; and the NPK and BNPK interaction had a significant effect on Shannon index (P < 0.05).Spearman’s correlation analysis showed that soil TN, NH4+–N and AP had a positive correlation with the alpha-diversity indices (P < 0.05, Table S3).
Taxonomic assignment of phyla showed that Proteobacteria dominated the diazotroph community composition, accounting for 65.9 to 96.4% of the total nifH sequences, followed by Cyanobacteria (≤ 9%), Verrucomicrobia (≤ 4%) and Firmicutes (≤ 1%) (Table S4). The hierarchical cluster diagram showed that samples from different treatments in the same layer were clustered into adjacent branches at the level of order (Fig. 4a). Rhizobiales (range 22.8–58.6%), Burkholderiales (0.9–32.3%), Rhodospirillales (2.9–23.6%), and Methylococcales (0.7–16.9%) were the four most abundant orders in all the treatments. Rhizobiales and Methylococcales increased in abundance with soil depth, whereas Burkholderiales and Rhodospirillales did the opposite. Rhizobiales, that include the core members of diazotrophs, were significantly higher (32.7–58.6%) in the subsoil than other orders. The BC contained the higher relative abundance of Rhizobiales (30.4% in topsoil and 58.6% in subsoil) compared with the other treatments (P < 0.05). In the topsoil, the BC was conducive to the enrichment of Chroococcales, accounting for 4.7%, but had a negative effect on Burkholderiales (P < 0.05). The NPK treatment enhanced the relative abundance of Methylococcales both in topsoil and subsoil compared with the other treatments (P < 0.05). Subsoil layer was more beneficial to the growth of anaerobic diazotrophs, such as Desulfuromonadales, Rhodocyclales, and Opitutales, that were distributing evenly in this layer (Fig. 4a) compared with the topsoil (P < 0.05).

Hierarchical cluster diagram of community composition of diazotrophs (at the order level) in various treatments (a). Principal coordinate analysis (PCoA) plot of diazotroph community structure in the topsoil and subsoil layers in various treatments (b). CK: no fertilizer; NPK: chemical fertilizer; BC: biochar; BNPK: biochar plus chemical fertilizer
We performed PCoA to visualize the difference in the vertical distribution of the diazotroph community structure (Fig. 4b). Similar to the hierarchical cluster (Fig. 4a), samples from the same layer were grouped together. The ANOSIM also confirmed significant differences in diazotroph communities between the topsoil and the subsoil (ANOSIM statistic R = 0.944, P = 0.001).
3.5 Relationships among soil properties, diazotroph community size and structure, and RNfix
The relationships between the vertical distribution of the diazotroph communities and soil properties were determined by MRT analysis (Fig. 5a). In total, the MRT model explained 67.74% of the variation in diazotroph communities, and NO3−–N was the major factor constraining the vertical pattern of diazotroph communities, splitting the communities into two major groups: one containing samples mostly from the topsoil (NO3−–N ≥ 1.69), and the other group with samples from the subsoil (NO3−–N < 1.69). To determine how soil properties impacted the diazotroph community in each soil layer, we performed Spearman’s correlations using the Mantel test (Table 4) between diazotroph communities and soil properties, and RDA (Fig. 5(b) and (c)).

Multivariate regression tree of diazotroph communities in topsoil and subsoil under different treatments (a). The soil chemical properties units were same as shown in Table 1. Redundancy Discriminant Analyses (RDAs) of diazotroph community structure in topsoil (b) and subsoil (c). The positions and lengths of the arrows indicate the directions and strengths, respectively, of the effects of soil properties on the diazotroph communities. CK: no fertilizer; NPK: chemical fertilizer; BC: biochar; BNPK: biochar plus chemical fertilizer. AP and AK: available phosphorus and potassium; NH4+–N: ammonium nitrogen; NO3−–N: nitrate nitrogen; C/N: the ratio of total carbon and nitrogen
The Mantel test results showed that diazotroph community was significantly correlated with C/N ratio, NH4+–N, SOM, AK, AP, and pH in the 0–20 cm, and with AK, NH4+–N, AP, and DON in the 20–40 cm. The soil properties included in RDA were selected on the basis of their collinearity and strength of their correlations with the diazotroph community structure (by VIF < 20 and significance (≤ 0.05) in Monte Carlo permutation test, Table S8) (Bai et al. 2015). The first two axes of the RDAs explained 57.18% and 7.78% of the diazotroph community variance in the 0–20 cm and 31.23% and 23.21% in the 20–40 cm soils, respectively, which were significant in the ANOVA permutation test (P < 0.05). Soil C/N ratio, NH4+-N and AP were the constraining variables with the highest interpretation ratio in the 0–20 cm soil, as were NH4+–N, AK and AP, NO3−–N in the 20–40 cm soil (Table S8).
The RNfix was significantly correlated with the diazotroph community structure both in the topsoil (Mantel test: R = 0.36, P = 0.008) and the subsoil (Mantel test: R = 0.53, P = 0.004) (Table 4), whereas it was significantly associated with nifH gene copies in the topsoil (Fig. 3b). Likewise, the diazotroph community structure was significantly correlated with nifH gene copies only in the topsoil (Mantel test: R = 0.51, P = 0.001). On the whole, it suggested that the diazotroph community structure and abundance were inconsistent in influencing the soil nitrogenase activity down the soil depth. The relationships among the vertical distribution of RNfix, diazotroph community abundance and structure and the major factors, including 16S rRNA gene copies, TN, SOM, NO3−–N, DOC, and pH, were explored by path analysis (Fig. 6). To simplify the SEM model, covariance coefficients among the soil properties were not noted in Fig. 6, but were detailed in Table S7. The final SEM demonstrated that RNfix was strongly influenced by diazotroph community structure (0.802), but had a weak relationship with the nifH gene copies (−0.028). It also confirmed that TN (0.934), SOM (0.473) and NO3−–N (0.981) were the dominant factors influencing 16S rRNA and nifH gene abundance, diazotroph community structure, respectively. Spearman’s correlations between the diazotroph community composition (at the level of order), such as Rhizobiales and Burkholderiales, and RNfix also confirmed the significant relationships (Table S6). The pH and DOC had significant relationships with RNfix, whereas 16S rRNA gene copies and SOM were the major factors influencing nifH gene copies.

Structural equation modeling (SEM) illustrating paths through which soil chemical properties, bacteria and diazotroph abundance, and diazotroph community structure affect the rate of nitrogen fixation (n = 24; df = 11, χ2 P = 0.413, GFI = 0.912, CFI = 0.999, RMSEA: 0.038, AIC: 79.370). Path coefficients (through 1000 bootstraps) are reflected in the width of the line, and positive and negative effects are showed by black and red, respectively. Solid arrows denote significant paths and dashed arrows denote nonsignificant paths. The asterisks indicate the significant level: * P < 0.05; ** P < 0.01; *** P < 0.001. “Diazotroph community” represents the first axis score derived from PCoA1 ordination. NO3−–N: nitrate nitrogen; DOC: dissolved organic carbon; TN: total nitrogen; SOM: soil organic matter
4 Discussion
Because of a strong link between the BNF and N content in agroecosystems, the responses of soil diazotroph communities to fertilizer regimes, such as N fertilizer additions, are important to the sustainability of soil N supply (Lin et al. 2018; Liu et al. 2020). Over the past decade, biochar has received great attention in agroecosystems as an amendment to cropland (Chen et al. 2019; Lehmann et al. 2011; Gul and Whalen 2016).
4.1 The dominant variables driving soil diazotroph abundance
In the present study, biochar had a significant influence on the abundance of soil bacteria as well as diazotrophs and its activity compared with chemical fertilizer (Table 2), increasing the abundance of nifH and 16S rRNA genes in the topsoil (P < 0.05, Fig. 1a), which was in accordance with previous studies (Azeem et al. 2021; Ibrahim et al. 2020; Jaiswal et al. 2019; Khadem et al. 2021; Liu et al. 2019b). This may be due to a number of reasons. First, C and N are the crucial components of microbial proteins, nucleic acids and phospholipids (Liu et al. 2020). The response of soil nifH gene copies to SOM revealed in ABT and SEM (Figs. 2 and 6), and the correlations with SOM, C/N and DOC in topsoil (0–20 cm) (P = 0.06) (Table S2) suggested that C source was the major factor driving soil diazotroph growth, especially in the topsoil. The significant correlation between the diazotroph community structure and SOM or C/N ratio in the topsoil (Table 4 and Fig. 5b) further confirmed this result. As an exogenous C, biochar plays an important role in soil energy maintenance and supply of microbial growth requirements in bulk soils (Che et al. 2018; Chen et al. 2019). Similarly, C sources had a significant effect on 16S rRNA gene abundance in the topsoil, but N sources were also the main constraints to bacteria in the subsoil (Table S2). The demand for N and C sources made TN and SOM the most important factors regulating bacterial growth (Figs. 2 and 6). Second, the high water-holding (Fig. S1) and temperature-maintaining capacity (data not showed) could protect microorganisms against drought, and were conducive to microorganism growth (Gul and Whalen 2016). The high porosity and large surface area of biochar make it favorable for microbial community adhesion via flocculation, surface adsorption, and cross-linking of cells (Lehmann et al. 2011).
The multivariate ANOVAs showed that, compared with the fertilizer regimes, soil depth had a stronger effect on 16S rRNA and nifH gene abundance and RNfix (P < 0.01, Table 2). In the present study, the abundance of nifH and 16S rRNA genes and RNfix were decreased in the subsoil, which could be explained by most of the soil available nutrients becoming deficient with soil depth (Table 1) (Liu et al. 2020; Wang et al. 2016). Some studies also indicated that the capacity of microorganism to use C source decreased in the subsoil due to diminished oxygen partial pressure and enhanced moisture content (Li et al. 2019b; Liu et al. 2020). Availability of nutrients to microorganisms down the soil depth is influenced greatly by the agricultural practices and biogeochemical processes (Li et al. 2019b; Qualls et al. 1991). In fact, the soil treated with biochar alone showed a greater reduction in the genes abundance (Fig. 1a), which might be attributed to the biochar adsorption and immobilization of nutrients in the topsoil (Gul and Whalen 2016), thus aggravating nutrient deficiency in the deeper soil. Hagemann et al. (2017) described an organo-mineral layer formation on the soil surface after biochar application, allowing mineral exchange between biochar and soil, which would result in nutrient retention through a variety of mechanisms; for instance, N adsorption can be increased via electrostatic or capillary forces, functional group complexation or a combination of these forces on the surface of the biochar particles as well as pores (Ibrahim et al. 2020), which was enabled by the application of biochar with N fertilizer, increasing the TN content in topsoil (Table 1). BNF is an energy intensive enzymatic process; although diazotrophs can fix N under well-aerated conditions, the high porosity of biochar makes N fixers in the subsoil may have to develop particular mechanisms and expend some energy to protect nitrogenase from inactivation by oxygen (Lin et al. 2018). Thus, it was not surprising that bacterial biomass, including diazotrophs, decreased in the BC treatment in the subsoil due to limited available carbon sources, but the decrease in diazotrophs was not significant (Fig. 1a). Diazotrophs would prefer to utilize inorganic N directly rather than engage in the energy expensive BNF processes in a larger microbial population (Norman and Friesen 2017).
Depending on the ecosystems, such as the topsoil and subsoil properties, N availability could have contrasting effects on the microbial community (Che et al. 2018; Reardon et al. 2014; Yang et al. 2019). An important interest in examining the dynamics of microbial community in subsoil is to understand the effect of fertilization on the whole root zone. In some situations, for example, when intense N mineralization in the topsoil (Zhang et al. 2020) masks the effects of fertilization, or when the sampling time is uncoupled from the duration of nutrient release from fertilizers, the knowledge of the effects of fertilizer application on soil ecosystems is poor. Therefore, soil microbial communities in the subsoil are more suitable as indicators of long-term changes in soil fertility and ecosystem functions because of their sensitivity and independence in response to the environment (Liu et al. 2020). Anaerobic environments are usually considered suitable for most diazotroph species (Kumar et al. 2020; Limmer and Drake 1996) and Shannon diversity of diazotroph community significantly increased with soil depth (Table 3), but the close relationship between the N sources, such as TN and NO3−–N (Table S2), and the genes abundance in the subsoil suggested an elevated importance of N source in the subsoil. Our study supported the view that C availability would not govern the abundance of diazotroph community when N availability was the limiting factor. This was consistent with several previous studies, implying that C and N sources played important roles in diazotroph abundance (Reardon et al. 2014; Wang et al. 2016). Additionally, the nifH abundance was significantly and positively correlated with the 16S rRNA in both soil layers (Fig. 1b), which was in accordance with the variations of diazotroph community in different agroecosystems reported by Reardon et al. (2014) and Wang et al. (2016). Taken together, it suggested that the growth of soil diazotrophs was influenced mainly by soil C and N dynamics (Figs. 2 and 6), and was related to the general microbial metabolism, rather than specific functional traits.
4.2 Biochar application was associated with an increase in BNF activity
Although the close links of diazotroph community size and structure with the soil variables, including pH, SOM, C/N and DOC, were revealed in the topsoil, RNfix showed strong positive correlations with these soil properties both in the topsoil and subsoil (Table S2). On the one hand, optimum pH for growth of most bacteria, including diazotrophs, is near neutral (Lin et al. 2018). The enhanced pH in the treatments with biochar addition would facilitate bacterial and diazotroph growth in the topsoil and enhance molybdenum (Mo) availability (Liu et al. 2019c), which is important because Mo is the key component of nitrogenase enzyme (Che et al. 2018; Tang et al. 2018; Zhang et al. 2020). On the other hand, these findings reflected dependence of nitrogenase activity on C source, especially for the free-living heterotrophic diazotrophs (Zhang et al. 2020), such as Burkholderiales and Rhodospirillales (Fig. 4a), whose abundance decreased with increasing soil depth and decreasing C source content. The Chroococcales order was increased in the topsoil of the BC compared with the BNPK and NPK (Fig. 4a), which might have been due to the sensitivity of the autotrophic diazotrophs to plentiful N, P and K nutrients (Zhang et al. 2020). The SEM result also verified that pH and DOC were the important factors influencing BNF activity (Fig. 6). Consequently, biochar application created an environment conducive to nitrogen fixation by the diazotroph community. The Burkholderiales order (represented mainly by genus Azohydromonas in Table S5) was another core lineage of diazotrophs, with the capacity of nitrogen-fixing symbiosis and plant growth promotion (Weber and King 2012);but it was suppressed in the biochar plots (Fig. 4a), which might be due to the sensitivity to soil salinity (Chen et al. 2018; Yang et al. 2016). The abundance of Azohydromonas decreased with an increase in salt concentration (Yang et al. 2016), and this was also confirmed in an incubation experiment (Chen et al. 2018). Our previous study (Guo et al. 2018) also showed that the total base cations, such as Ca2+, Mg2+ and Na+, were higher in the BC and BNPK than in the other treatments (P < 0.05), and it could be an ash component of the biochar pyrolysis process.
In agroecosystems, agricultural practices will often create and govern individual microecosystems, whereby added fertilizers or organic substrates may induce the succession of different types of microbial communities (Yang et al. 2019) varying spatially and temporally and due to climatic changes (Wang et al. 2016; Liu et al. 2020). Depending on the fertilizer regime, we speculated that diazotrophs in the BNPK would utilize inorganic N and organic C sources for growth. Due to the scarcity of inorganic nitrogen sources at the harvest stage (sampling time), bacteria (including diazotrophs) would be expected to show little, if any, biomass increase. Hence, the BNPK effect was lower than the BC effect in the topsoil, e.g. in the case of nifH abundance (Fig. 1a), which was not contradictory to the views that N fertilization usually negatively affects diazotrophs and the BNF (Wang et al. 2017; Tang et al. 2017). Regarding the mechanism, we speculated that biochar application would promote root growth (Gul and Whalen 2016) and increase root biomass (Abiven et al. 2015), length, and area (Jaiswal et al. 2019); however, nutrient supply due to chemical fertilization might have a negative feedback on root length extension (Chew et al. 2020). The BNPK might cause some inhibition of peanut root growth in topsoil, and a significant proportion of the microorganisms in the collected bulk soil samples were driven by the root metabolic activities during the peanut growing season, such as residual roots and their exudates (Zhao et al. 2021), which might have resulted in no significant increase in 16S rRNA gene abundance in topsoil compared with the BC. In spite of this, the BNPK failed to prevent a highly significant increase in BNF rate in the topsoil (P < 0.001), while mitigating the inhibitory effect of N fertilization on such rate in the subsoil (Fig. 3a).
4.3 Fertilization-induced succession of diazotroph community composition governed RNfix
Biochar and chemical fertilizer both had a significant effect on RNfix (Table 2), but biochar played a positive one, while chemical fertilizer did the opposite (Fig. 3a). Although the NPK did not result in lower nifH gene abundance in the subsoil (Fig. 1a), the extremely low RNfix indicated that BNF was negatively affected by chemical fertilization and, certainly, the RNfix had the relative moderate values in the BNPK. In addition, the RNfix showed a negative correlation with AP and AK (Table S2), which could be the consequence of the application of chemical fertilizers (i.e., calcium superphosphate and potassium chloride). Clearly, changes in RNfix were governed by fertilizer regimes, which drove vertical variations in diazotroph community structure through the vertical distribution of soil NO3−–N (Fig. 5a). The findings of this present study suggested that the RNfix was significantly related to the abundance of diazotrophs in the topsoil, but not in the subsoil (Fig. 3). Actually, the SEM showed that diazotroph community structure had the greatest effect on BNF activity (Fig. 6). The RNfix, closely linked to diazotroph community structure (Table 4), would be significantly affected by changes in diazotroph community composition (Table S6).
Members of the Methylococcales order, capable of nitrogen fixation, were abundant in both soil layers in the NPK treatment (Fig. 4a). Methylococcales are also active in nitrification (Llorens-Marès et al. 2020), which is generally promoted by applying N fertilizer (Liu et al. 2020). When N leaching occurs, nitrification may also occur in the subsoil (Llorens-Marès et al. 2020), potentially leading to higher abundance of Methylococcales in the subsoil than the topsoil (Fig. 4a). This implied that members of Methylococcales order might be involved more in the nitrification process than the BNF, especially in the subsoil, though Methylococcales represent a minor proportion of nitrifiers. One mechanism for the inhibitory effect of N fertilizer on BNF activity may be a shift in the function of the associated microbiota rather than a decrease in the abundance of diazotrophs (Table S6). A large decrease in the abundance of Methylococcales and an increase in RNfix in the BNPK treatment (Figs. 3a and 4a) indicated that chemical fertilizer combined with biochar could partly offset and even reverse the negative effects of N fertilization on RNfix (Fig. 3a), which might be due to the fact that N sequestration in the topsoil in the BNPK reduced N leaching to the subsoil. Rhizobia are a paraphyletic group of soil-borne bacteria that induce nodule organogenesis and fix N2 from atmosphere in symbiosis with plants (Garrido-Oter et al. 2018) demonstrating a consistent distribution with root biomass (Liu et al. 2020). More than 60% of peanut roots were in the 0 to 30 cm soil layer (Jongrungklang et al. 2012). In the present study, a considerable number of diazotroph species belonged to the Rhizobiales order (Fig. 4a), as one of the core microbiota involved in BNF (Lin et al. 2018; Zhang et al. 2020). The significant correlation between relative abundance of Rhizobiales and RNfix confirmed the close link between Rhizobiales and BNF activity (Table S6). The high relative abundance of Rhizobiales in the BC treatment (30% and 59% in the topsoil and subsoil, respectively) compared with the BNPK and NPK (P < 0.05) might have been the reason for the increased BNF activity induced by biochar application alone, especially in the subsoil (Fig. 3a). It also supports the preceding view that biochar promoted the growth of the peanut roots and microorganisms associated with them. In summary, although there was a link between the diazotroph abundance and N2 fixation rate in the topsoil, the variation in diazotroph community composition with soil depth was associated significantly with the N2 fixation rate in different soil layers and was therefore a key driver of BNF activity variation. This study also revealed the reasons for the abundance and activity of diazotrophs in the subsoil responding inconsistently to chemical fertilizer or biochar application.
5 Conclusions
Our study demonstrated that the soil carbon source (e.g. SOM, C/N ratio and DOC) and pH were the main environmental drivers of diazotroph abundance and BNF activity in the topsoil, whereas soil nitrogen source (e.g.TN and NO3−–N) was the primary factor governing the abundance of bacteria and diazotrophs in the subsoil. Applying biochar was associated with the increase in soil carbon and pH, which led to an increase in the abundance of bacteria and diazotrophs and enhanced BNF activity in the topsoil.
NO3−–N greatly impacted the vertical pattern of diazotroph community structure, which was associated with the fertilizer regime. Chemical fertilizer negatively affected BNF and caused a significant decrease in RNfix in the subsoil, likely by a mechanism related to changes in diazotroph community structure (Methylococcales) rather than abundance. Biochar addition could induce an increase in the core lineage of diazotroph community (e.g. Rhizobiales) and had a significant positive effect on the BNF, though impeded the growth of bacteria in the subsoil. The combined application of biochar and chemical fertilizer partly offset the negative effects when they were applied separately. Hence, we suggested that diazotroph community structure played an important role in diazotroph activity, regulating BNF.
Data availability
All data generated or analyzed during this study are included in this published article (and its supplementary information files).
References
Abiven S, Hund A, Martinsen V, Cornelissen G (2015) Biochar amendment increases maize root surface areas and branching: a shovelomics study in Zambia. Plant Soil 395:45–55. https://doi.org/10.1007/s11104-015-2533-2
Azeem M et al (2021) Tea leaves biochar as a carrier of Bacillus cereus improves the soil function and crop productivity. Appl Soil Ecol 157:103732. https://doi.org/10.1016/j.apsoil.2020.103732
Bai SH et al (2015) Wood biochar increases nitrogen retention in field settings mainly through abiotic processes. Soil Biol Biochem 90:232–240. https://doi.org/10.1016/j.soilbio.2015.08.007
Bao S (2000) Soil Argri-chemistry Analysis 3rd, Ed. China Agriculture Press, Beijing
Belnap J (2002) Nitrogen fixation in biological soil crusts from southeast Utah USA. Biol Fert Soils 35:128–135. https://doi.org/10.1007/s00374-002-0452-x
Che R et al (2018) Autotrophic and symbiotic diazotrophs dominate nitrogen-fixing communities in Tibetan grassland soils. Sci Total Environ 639:997–1006. https://doi.org/10.1016/j.scitotenv.2018.05.238
Chen G, Zhang Z, Zhang Z, Zhang R (2018) Redox-active reactions in denitrification provided by biochars pyrolyzed at different temperatures. Sci Total Environ 615:1547–1556. https://doi.org/10.1016/j.scitotenv.2017.09.125
Chen W, Meng J, Han X, Lan Y, Zhang W (2019) Past, present, and future of biochar. Biochar 1:75–87. https://doi.org/10.1007/s42773-019-00008-3
Chen K, Peng J, Li J, Yang Q, Zhan X, Liu N, Han X (2020) Stabilization of soil aggregate and organic matter under the application of three organic resources and biochar-based compound fertilizer. J Soils Sediments 20:3633–3643. https://doi.org/10.1007/s11368-020-02693-1
Chew J et al (2020) Biochar-based fertilizer: supercharging root membrane potential and biomass yield of rice. Sci Total Environ 713:136431. https://doi.org/10.1016/j.scitotenv.2019.136431
Furlan AL, Bianucci E, Castro S, Dietz K-J (2017) Metabolic features involved in drought stress tolerance mechanisms in peanut nodules and their contribution to biological nitrogen fixation. Plant Sci 263:12–22. https://doi.org/10.1016/j.plantsci.2017.06.009
Garrido-Oter R, Nakano RT, Dombrowski N, Ma K-W, McHardy AC, Schulze-Lefert P (2018) Modular traits of the rhizobiales root microbiota and their evolutionary relationship with symbiotic rhizobia. Cell Host Microbe 24:155-167.e155. https://doi.org/10.1016/j.chom.2018.06.006
Gul S, Whalen JK (2016) Biochemical cycling of nitrogen and phosphorus in biochar-amended soils. Soil Biol Biochem 103:1–15. https://doi.org/10.1016/j.soilbio.2016.08.001
Guo C, Li N, Peng J, Gao T, Ma L, Han X-R (2018) Direct returning of maize straw or as biochar to the field triggers change in acidity and exchangeable capacity in soil. J Plant Nutr Fertilizers 24:1205–1213
Hagemann N et al (2017) Organic coating on biochar explains its nutrient retention and stimulation of soil fertility. Nat Commun 8:1089. https://doi.org/10.1038/s41467-017-01123-0
Hu J et al (2019) Utilization of the saccharification residue of rice straw in the preparation of biochar is a novel strategy for reducing CO2 emissions. Sci Total Environ 650:1141–1148. https://doi.org/10.1016/j.scitotenv.2018.09.099
Ibrahim MM, Hu K, Tong C, Xing S, Zou S, Mao Y (2020) De-ashed biochar enhances nitrogen retention in manured soil and changes soil microbial dynamics. Geoderma 378:114589. https://doi.org/10.1016/j.geoderma.2020.114589
Jaiswal AK, Graber ER, Elad Y, Frenkel O (2019) Biochar as a management tool for soilborne diseases affecting early stage nursery seedling production. Crop Prot 120:34–42. https://doi.org/10.1016/j.cropro.2019.02.014
Jongrungklang N, Toomsan B, Vorasoot N, Jogloy S, Boote KJ, Hoogenboom G, Patanothai A (2012) Classification of root distribution patterns and their contributions to yield in peanut genotypes under mid-season drought stress. Field Crop Res 127:181–190. https://doi.org/10.1016/j.fcr.2011.11.023
Khadem A, Raiesi F, Besharati H, Khalaj MA (2021) The effects of biochar on soil nutrients status, microbial activity and carbon sequestration potential in two calcareous soils. Biochar 3:105–116. https://doi.org/10.1007/s42773-020-00076-w
Kumar U, Nayak AK, Sahoo S, Kumar A, Kaviraj M, Shahid M (2020) Combined effects of elevated CO2, N fertilizer and water deficit stress on diazotrophic community in sub-humid tropical paddy soil. Appl Soil Ecol 155:103682. https://doi.org/10.1016/j.apsoil.2020.103682
Lehmann J, Rillig MC, Thies J, Masiello CA, Hockaday WC, Crowley D (2011) Biochar effects on soil biota—a review. Soil Biol Biochem 43:1812–1836. https://doi.org/10.1016/j.soilbio.2011.04.022
Li S, Wang S, Shangguan Z (2019a) Combined biochar and nitrogen fertilization at appropriate rates could balance the leaching and availability of soil inorganic nitrogen. Agric Ecosyst Environ 276:21–30. https://doi.org/10.1016/j.agee.2019.02.013
Li X, Wang H, Li X, Li X, Zhang H (2019b) Shifts in bacterial community composition increase with depth in three soil types from paddy fields in China. Pedobiologia 77:150589. https://doi.org/10.1016/j.pedobi.2019.150589
Limmer C, Drake HL (1996) Non-symbiotic N2-fixation in acidic and pH-neutral forest soils: aerobic and anaerobic differentials. Soil Biol Biochem 28:177–183. https://doi.org/10.1016/0038-0717(95)00118-2
Lin Y et al (2018) Long-term application of lime or pig manure rather than plant residues suppressed diazotroph abundance and diversity and altered community structure in an acidic Ultisol. Soil Biol Biochem 123:218–228. https://doi.org/10.1016/j.soilbio.2018.05.018
Liu XJ et al (2013) Enhanced nitrogen deposition over China. Nature 494:459–462. https://doi.org/10.1038/nature11917
Liu C, Wang H, Li P, Xian Q, Tang X (2019) Biochar’s impact on dissolved organic matter (DOM) export from a cropland soil during natural rainfalls. Sci Total Environ. 650:1988–1995. https://doi.org/10.1016/j.scitotenv.2018.09.356
Liu Q et al (2019) Biochar application as a tool to decrease soil nitrogen losses (NH3 volatilization, N2 O emissions, and N leaching) from croplands: options and mitigation strength in a global perspective. Glob Chang Biol 25:2077–2093. https://doi.org/10.1111/gcb.14613
Liu X et al (2019b) Impact of biochar amendment on the abundance and structure of diazotrophic community in an alkaline soil. Sci Total Environ 688:944–951. https://doi.org/10.1016/j.scitotenv.2019.06.293
Liu M et al (2020) Nitrogen leaching greatly impacts bacterial community and denitrifiers abundance in subsoil under long-term fertilization. Agric Ecosyst Environ 294:106885. https://doi.org/10.1016/j.agee.2020.106885
Llorens-Marès T, Catalan J, Casamayor EO (2020) Taxonomy and functional interactions in upper and bottom waters of an oligotrophic high-mountain deep lake (Redon, Pyrenees) unveiled by microbial metagenomics. Sci Total Environ. 707:135929. https://doi.org/10.1016/j.scitotenv.2019.135929
Mia S, van Groenigen JW, van de Voorde TFJ, Oram NJ, Bezemer TM, Mommer L, Jeffery S (2014) Biochar application rate affects biological nitrogen fixation in red clover conditional on potassium availability. Agric Ecosyst Environ 191:83–91. https://doi.org/10.1016/j.agee.2014.03.011
Norman JS, Friesen ML (2017) Complex N acquisition by soil diazotrophs: how the ability to release exoenzymes affects N fixation by terrestrial free-living diazotrophs. ISME J 11:315–326. https://doi.org/10.1038/ismej.2016.127
Oram NJ, van de Voorde TFJ, Ouwehand G-J, Bezemer TM, Mommer L, Jeffery S, Groenigen JWV (2014) Soil amendment with biochar increases the competitive ability of legumes via increased potassium availability. Agri Ecosyst Environ 191:92–98. https://doi.org/10.1016/j.agee.2014.03.031
Qualls RG, Haines BL, Swank WT (1991) Fluxes of dissolved organic nutrients and humic substances in a deciduous forest. Ecology 72:254–266. https://doi.org/10.2307/1938919
Reardon CL, Gollany HT, Wuest SB (2014) Diazotroph community structure and abundance in wheat–fallow and wheat–pea crop rotations. Soil Biol Biochem 69:406–412. https://doi.org/10.1016/j.soilbio.2013.10.038
Rosch C, Mergel A, Bothe H (2002) Biodiversity of denitrifying and dinitrogen-fixing bacteria in an acid forest soil. Appl Environ Microbiol 68:3818–3829. https://doi.org/10.1128/aem.68.8.3818-3829.2002
Tang Y, Zhang M, Chen A, Zhang W, Wei W, Sheng R (2017) Impact of fertilization regimes on diazotroph community compositions and N2-fixation activity in paddy soil. Agric Ecosyst Environ 247:1–8. https://doi.org/10.1016/j.agee.2017.06.009
Tang Y, Yu G, Zhang X, Wang Q, Ge J, Liu S (2018) Changes in nitrogen-cycling microbial communities with depth in temperate and subtropical forest soils. Appl Soil Ecol 124:218–228. https://doi.org/10.1016/j.apsoil.2017.10.029
Verhoeven E, Six J (2014) Biochar does not mitigate field-scale N2O emissions in a Northern California vineyard: an assessment across two years. Agric Ecosyst Environ 191:27–38. https://doi.org/10.1016/j.agee.2014.03.008
Wang J, Zhang D, Zhang L, Li J, Raza W, Huang Q, Shen Q (2016) Temporal variation of diazotrophic community abundance and structure in surface and subsoil under four fertilization regimes during a wheat growing season. Agric Ecosyst Environ 216:116–124. https://doi.org/10.1016/j.agee.2015.09.039
Wang C, Zheng M, Song W, Wen S, Wang B, Zhu C, Shen R (2017) Impact of 25 years of inorganic fertilization on diazotrophic abundance and community structure in an acidic soil in southern China. Soil Biol Biochem 113:240–249. https://doi.org/10.1016/j.soilbio.2017.06.019
Weber CF, King GM (2012) The phylogenetic distribution and ecological role of carbon monoxide oxidation in the genus Burkholderia. FEMS Microbiol Ecol 79:167–175. https://doi.org/10.1111/j.1574-6941.2011.01206.x
Xu N, Tan G, Wang H, Gai X (2016) Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur J Soil Biol 74:1–8. https://doi.org/10.1016/j.ejsobi.2016.02.004
Yang H, Hu J, Long X, Liu Z, Rengel Z (2016) Salinity altered root distribution and increased diversity of bacterial communities in the rhizosphere soil of Jerusalem artichoke. Sci Rep 6:20687. https://doi.org/10.1038/srep20687
Yang L et al (2019) Diazotroph abundance and community structure are reshaped by straw return and mineral fertilizer in rice-rice-green manure rotation. Appl Soil Ecol 136:11–20. https://doi.org/10.1016/j.apsoil.2018.12.015
Zhang X et al (2020) Depression of soil nitrogen fixation by drying soil in a degraded alpine peatland. Sci Total Environ 747:141084. https://doi.org/10.1016/j.scitotenv.2020.141084
Zhao M et al (2021) Root exudates drive soil-microbe-nutrient feedbacks in response to plant growth. Plant Cell Environ 44:613–628. https://doi.org/10.1111/pce.13928
Acknowledgements
We are very grateful to Fangfang Cai, Wei Hua, Zhiming Ma, Peng Jin and Peng Geng for their insightful suggestions and comments on this paper.
Funding
This work was funded by the National Natural Science Foundation of China (Grant No. 31972511), the National Key R&D Program of China (Grant No. 2018YFD0201001) and China Agriculture Research System of MOF and MARA (CARS-13).
Author information
Authors and Affiliations
Contributions
XH conceived, designed and financially supported the study; KC analyzed the data and wrote the paper, and conducted the analytical work with SZ and NL; NL and JY supervised the field trial; XZ reviewed and edited the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Conflict of interests
We declare that we have no financial and personal relationships with other people or organizations that can inappropriately influence our work. There is no professional or other personal interest of any nature or kind in any product, service and/or company that could be construed as influencing the position presented in, or the review of, this manuscript.
Supplementary Information
Below is the link to the electronic supplementary material.
Additional file 1.
Table S1. Operating instructions for q-PCR. Tables S2, S3, and S6. Spearman’s correlation analyses. Tables S4 and S5. Comparison of phyla and genera of diazotroph community. Table S7. Covariance coefficients in SEM model. Table S8. Results of the permutation test for RDAs. Figure S1. Soil moisture content.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chen, K., Li, N., Zhang, S. et al. Biochar-induced changes in the soil diazotroph community abundance and structure in a peanut field trial. Biochar 4, 26 (2022). https://doi.org/10.1007/s42773-022-00133-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s42773-022-00133-6