
Abstract
We investigated by snow tracking and GPS telemetry how wolves Canis lupus used a dense (4 km/km2) network of forest roads for travel and scent marking. Forty-six percent of wolf trails but only 4.6% of telemetry locations were on forest roads. Wolves used forest roads to travel fast and far across their home ranges but spent relatively little time on roads, especially on those with high traffic levels and during the time of highest human activity. The probability of scent marking was higher on roads than off-road and increased with traffic intensity on roads and close to junctions. Our findings suggest that wolves take advantage of the forest road infrastructure, while minimising human encounters by spatio-temporal avoidance of all roads, even those with negligible traffic. The ongoing expansion and improvements of the forest road network might lead to elevated costs for wolves associated with avoidance of humans and roads.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Roads are an omnipresent and ever-expanding feature of all human-modified landscapes (Trombulak and Frissell 2000). High-traffic roads profoundly affect ecosystems by increasing wildlife mortality, changing animal abundance and behaviour, and modifying natural habitats (reviewed in Forman and Alexander 1998; Trombulak and Frissell 2000; Coffin 2007; Fahrig and Rytwinski 2009). Roads have been considered an important factor limiting distribution, dispersal and habitat quality for large carnivores, including wolves Canis lupus (Jędrzejewski et al. 2004; Potvin et al. 2005; Cullingham et al. 2016). Frequently used roads constitute hotspots of human-caused wolf mortality due to traffic collisions, hunting, and poaching (Recio et al. 2018). Therefore, wolves tend to avoid high-traffic roads, especially during times of highest human activity (Theuerkauf et al. 2003, 2007; Lesmerises et al. 2013; Benson et al. 2015).
Roads less intensively used by humans can disturb ecosystems as well, but their ecological effects are subtler and more localised (Suárez‐Esteban et al. 2013). Minor roads built for the purpose of forest management (hereafter: forest roads) contribute to forest fragmentation and degradation of forest ecosystems (Reed et al. 1996; Reimoser and Gossow 1996; Heilman et al. 2002; Saunders et al. 2002; Hall et al. 2003; Laurance et al. 2009). Although forest roads usually have too little traffic to pose a risk of wildlife–vehicle collisions, they may increase human disturbance and wildlife mortality by easing access for tourists, local inhabitants, hunters, and poachers (Wabakken et al. 2001; Person and Russell 2008). On the other hand, minor roads rarely used by humans may act as corridors for terrestrial animals and plants and enhance some ecosystem services (Haddad et al. 2003; Suárez‐Esteban et al. 2013).
Studies on wolf response to forest roads showed contrasting results, from selection for movement and scent marking (Zub et al. 2003; Zimmermann et al. 2014; Dickie et al. 2017) to spatial and spatio-temporal avoidance depending on levels of human activity (Theuerkauf et al. 2003; Whittington et al. 2005). When travelling on roads, wolves move faster, cover larger daily distances, and encounter prey more often (James and Stuart-Smith 2000; Hebblewhite et al. 2005; Whittington et al. 2011; Dickie et al. 2017). Wolf response to roads with low traffic may vary according to wolf behavioural and social status, time of day and season (Zimmermann et al. 2014; Mancinelli et al. 2019). Moreover, minor roads can play an important role in scent communication, which serves as a main information medium for establishment and maintenance of wolf territories (Peters and Mech 1975; Rothman and Mech 1979; Zub et al. 2003). Forest roads and their junctions serve as conspicuous sites for scent marks, providing exposition and, therefore, enhancing the strength of scent signals (Rothman and Mech 1979; Asa et al. 1985). Several authors suggested that the response of wolves to roads depends also on road density, but the results varied among study areas (Whittington et al. 2005; Houle et al. 2010; Zimmermann et al. 2014).
In this study, we investigated the use of an exceptionally dense forest road network (4 km/km2), the highest ever reported in wolf home ranges (Theuerkauf et al. 2007; Houle et al. 2010; Gurarie et al. 2011; Zimmermann et al. 2014; Dickie et al. 2017). Specifically, we used GPS telemetry and snow tracking to quantify (1) the daily distances travelled and the speed of wolves when on roads and when off-road, (2) the spatial and daily patterns of forest road selection for wolf travel and (3) scent marking in relation to traffic intensity.
Methods
Study area
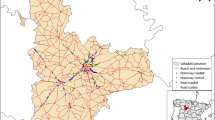
We conducted our study in the western part (ca. 250 km2) of the Lower Silesia Forest, western Poland (N 51° 14.2′–51° 27.1′, E 14° 57.5′–15° 10.9′, Fig. 1). Average annual temperature during the study was 10 °C (daily mean temperature varying between −16 and 29 °C), and snow cover (up to 10 cm deep) persisted for 7–71 days each winter (data provided by the Institute of Meteorology and Water Management). The area is flat (altitude 120–160 m a.s.l.) and is predominantly covered by forests managed by the Polish State Forests, a governmental organisation specialised in timber production. Forests are primarily composed of even-aged Scots pine Pinus sylvestris plantations, which are managed throughout the year by regular clearcutting, planting, and fencing of young stands (Bena 2012). To improve the efficiency of forest management and to enable access to logging sites, the forest is divided into rectangular compartments of 750 m × 375 m, most of which are surrounded by straight forest roads of 3–5 m width. Many such compartments are intersected by an additional road, which further divides tree plantations into squares of 375 × 375 m. As a result, the landscape consists of rectangular patches divided by a network of straight forest roads (Fig. 1). The density of forest roads is almost uniformly 4 km/km2 throughout the study area. Forest roads are used throughout the year, primarily by motor vehicles of foresters and heavy-duty machinery for wood extraction and transportation. Additionally, traffic on forest roads increases seasonally due to hunting, antler collecting, as well as berry and mushroom picking. The public is not allowed to use motor vehicles on forest roads. Due to shallow snow cover, these roads are rarely ploughed. Only few public paved roads run through the study area, their density is 0.2 km/km2.

Study area in the Lower Silesia Forest, Poland, with snow tracking trails (of three wolf packs) and 100% minimum convex polygons of three individuals followed by GPS telemetry (2963 locations)
Wolves began recolonising the study area in the 1980s and the process accelerated in the 2000s (Wolsan et al. 1992; Nowak and Mysłajek 2016). During the first two winters of our study (2012/13 and 2013/14), the area was occupied by one wolf pack, which later split its territory into two. The numbers of wolves in the original and the two resulting packs were dynamic throughout the study, counting from four to eight individuals per pack in winter. Wolves prey predominantly on three species of ungulates: red deer Cervus elaphus, roe deer Capreolus capreolus and wild boar Sus scrofa (Nowak et al. 2011). Wolves are legally protected in Poland; therefore, there has been no legal wolf hunting in the area since 1995. However, wolf poaching is regularly reported in the whole country (Gula 2008; Gula et al. 2009; Nowak and Mysłajek 2016).
Classification of forest roads
We classified roads by visual inspection at crossroads. Based on this inspection and through 100 photo-trapping (Bushnell Trophy Cam HD Aggressor) sessions (50 in winter and 50 in summer) lasting 6–10 days, we functionally classified forest roads into three categories: (1) high-traffic road (density 0.4 km/km2): maintained hard-surfaced gravel road used frequently for wood transportation, forestry activities and hunting (traffic 5–20 motor vehicles per day), (2) medium-traffic road (0.8 km/km2): road with visible signs of frequent use and little vegetation covering the middle of road surface, used regularly (up to five vehicles per day, mean four vehicles per week), (3) low-traffic road (2.8 km/km2): mostly passable by four-wheel drive cars but with most of the surface overgrown by low vegetation, used only sporadically when logging or hunting activities occurred in its proximity. Overall traffic on forest roads peaked during the working hours of forestry personnel (0700–1600 h) and people rarely drove on forest roads during the night. In the study area, sunrise was between 0444 and 0803 h and sunset between 1553 and 2118 h of local time. We did not include logging trails in the analysis, as they appear only temporarily during wood harvesting.
Data collection
To investigate road selection using telemetry data, we captured three wolves using Belisle foot snares (Belisle Enterprises, Canada) and fitted them with GPS collars that also included VHF transmitters (LOTEK Wireless Inc., Canada). The three collared individuals belonged to two packs: a young female (monitored December 2012–January 2013) belonged to the original pack that later split its territory (see above), whereas a breeding female (January 2015–September 2016) and a young male (January–August 2016) belonged to the northern of the two packs created from the original pack (Fig. 1). There was no collared individual in the southern pack created by the split. The GPS collars obtained locations at various intervals: every 2 h for the young female, and at 11-h intervals for the other two individuals with additional 1-h intervals for 1-week periods every month from December to April. The Polish authorities (General Directorate for Environmental Protection: DOP-OZGIZ.6401.08.12.2011.ls, and Regional Directorate for Environmental Protection in Gorzów Wielkopolski: WPN-I.6401.347.2015.JK) approved capture and handling protocols.
To investigate road selection using snow-tracking data, between December and March of 2012–2017, we located fresh wolf tracks when snow cover permitted. We used recent (obtained during the last 24 h) GPS locations and VHF signals of collared individuals to locate tracks. During winters when no individual was collared (2011/12, 2013/14 and 2016/17) or when tracking the non-collared pack, we searched for wolf tracks on forest roads, driving the road network of 1.5 × 1.5 km mesh size intersecting the whole study area. In the latter case, snow tracking always started at places where wolves crossed roads. When we found fresh wolf trails, we followed them on foot while recording the tracks on a hand-held GPS (Garmin GPSmap 62 s) set to record locations at 20-m intervals. We followed the wolf trails in both directions as long as the tracks were visible, or only backwards in case of very fresh tracks.
Finally, to investigate road use for scent marking, during snow-tracking sessions, we recorded all wolf scent marks on the trail (urination, scratching, defaecation). We attributed wolf trails and scent marks to a road if they were located on the road surface or within 3 m from the road edge. To avoid overestimation of road use by wolves, we excluded from the analyses trails that were shorter than 1 km or entirely on roads.
Data analysis
To analyse road selection using GPS-telemetry data, we applied a generalised linear model (GLM) with binomial distribution and logit-link function in R 3.6.1 (R Core Team 2019) using a binary response variable (1: wolf telemetry location, 0: random point). We generated random points within the seasonal (winter: December–March, spring–autumn: April–November) home ranges (100% minimum convex polygons) of each GPS-collared individual. For each wolf and each season, we used the same number of random points as there were telemetry locations (2,963 random points in total). We included as fixed variables the habitat type as a categorical factor with four levels (0—off-road, 1—low-traffic road, 2—medium-traffic road, 3—high-traffic road) and period of day as a nominal factor (working hours of forestry personnel: 0700–1600 h, off-work: the remaining time). We considered a telemetry location or a random location as on a road (road types 1–3) if it was within a 15-m buffer of the midline of the road, which corresponds to the location accuracy of the GPS collars model that we used (Di Orio et al. 2003).
To analyse road selection using snow-tracking data, we applied a GLM the same way as described above, but without the variable “period of day”. We generated wolf trail points every 218 m (to achieve the same number as of telemetry locations) along the wolf trails recorded during snow tracking (2963 wolf trail points) and the same number of random points inside the minimum convex polygon encompassing all the wolf trails.
To assess the daily distances travelled by wolves, on roads and off-road, we used telemetry data obtained during 128 days of intensive sampling of GPS locations (1-h or 2-h intervals) in winter. We corrected distance travelled by dividing the cumulative straight-line distances between consecutive wolf locations by correction coefficients provided by Mills et al. (2006): 0.68 and 0.60 for 1-h and 2-h GPS fix intervals, respectively. To assess wolf travel speed on roads and off-road, we combined telemetry data (proportion of time spent on roads) with snow-tracking data (proportion of distance on roads).
To analyse wolf use of roads for scent marking, we used GLM with binomial distribution and logit-link function in R 3.6.1 to test whether the probability of scent marking was related to its location (on roads or off-road, road category) and distance to junctions. We included a binary response variable (scent mark location/random point along wolf trail) and three fixed variables: habitat type (0—off-road, 1—low-traffic road, 2—medium-traffic road, 3—high-traffic road), distance to the closest junction (m) and their interaction. We performed the analyses separately for each type of scent marking (scats, urination, scratching). We generated the random points along wolf trails in numbers equal to numbers of scent marks of each type.
All spatial analyses were performed in QGIS version 3.2.3 (Quantum GIS Development Team 2018). We used model selection based on the Akaike Information Criterion (AICc) to rank models in the MuMIn package (Bartoń 2019) in R 3.6.1.
Results
Daily patterns of road use
We obtained 2963 GPS locations of wolves, 2009 in winter (488 of young female, 989 of breeding female, 532 of young male) and 954 during the rest of the year (breeding female: 732, young male: 222). The breeding female and her son spent most of the time separately; only 28% of telemetry locations obtained simultaneously for the two individuals (n = 396) were within a 300-m distance from each other. Only 4.6% of all wolf telemetry locations were on roads (Table 1). The probability of a wolf location was best explained by the type of habitat and its interaction with the time of day (Table 2). Thus, based on telemetry data, wolves strongly avoided roads, especially medium- and high-traffic roads (Fig. 2a). The avoidance of medium- and high-traffic roads was stronger during the working hours of forestry personnel; whereas, period of day had no effect on the use of low-traffic roads (Fig. 2a).

Relative probability predicted by GLM (with 95% CI) of a a wolf telemetry location (during the working hours of forestry personnel and during the rest of the day) and b a wolf snow trail point location to be off-road or on a forest road (low, medium and high traffic intensity) in the Lower Silesia Forest, western Poland in 2012–2017 (n = 2963 telemetry locations of three individuals and 646 km of snow tracking)
Road use and selection based on snow tracking
We collected snow tracking data from 646 km of wolf trails in 74 continuous tracking sessions (1.9–36.5 km) over the course of six winters (2011/12–2016/17). Most of the trails (48 tracking sessions, 424 km, winter seasons 2011/12–2013/14) belonged to the original pack, which later split its territory (see above). Twenty-two tracking sessions (147 km, winter seasons 2014/15–2017/18) belonged to the after-division northern pack with the GPS-collared breeding female and young male. Four tracking sessions (75 km, winter seasons 2014/15–2017/18) were located in the after-division southern pack’s territory (without GPS-collared individuals). Out of 74 tracking sessions, 44 originated from locations obtained via telemetry (22 tracking sessions of the original pack and 22 of the northern pack with two collared individuals). Almost half of the wolf trails were located on forest roads (Table 1). Based on snow-tracking data, wolves strongly selected roads (Table 3). Wolves preferred to travel on forest roads of medium and high traffic levels (Fig. 2b).
Daily travel distances and speed in winter
Wolves travelled on average 20.1 ± 1.8 km/day (mean ± 95% CI), which, considering an average of 46% of length travelled on roads in winter, corresponds to 9.3 ± 0.8 km on roads daily. As wolves spent only 5.4% of time on roads in winter (Table 1), this corresponds to an average of 1.3 h per day. Wolf locations appeared on high-traffic roads on only 0.8% of the days when telemetry locations were obtained (on average 4 min per day), on medium-traffic roads in 3.1% of the days (16 min per day), and on low-traffic roads on 12.3% of the days (58 min per day). Their mean speed on roads was 7.1 km/h and was highest on high-traffic roads (21.5 km/h), intermedium on medium-traffic roads (9.4 km/h) and lowest on low-traffic roads (5.4 km/h).
Scent marking
During snow tracking, we found 1,278 wolf scent marks, of which 71% were located on roads. Average scent-marking intensity was 2.0 scent marks per km of wolf trail, and 1.1 and 3.1 scent marks/km off-road and on roads, respectively (Table 1). Probability of marking with scats was best explained by the type of habitat; whereas, the probability of urination or scratching was also affected by the distance to the closest crossroad and the interaction of these variables (Table 4). Thus, wolves selected roads for scent marking and the probability of marking with scats increased with traffic intensity on roads (Fig. 3). Wolves were more likely to scent mark with urine and by scratching close to junctions when marking on roads (Fig. 4).

Relative probability predicted by GLM (with 95% CI) of a wolf scent marking (scats, urine and scratching) in relation to habitat (off-road or forest road of low, medium and high traffic intensity) in the Lower Silesia Forest, western Poland in 2012–2017 (n = 1278 scent marks)

Relative probability predicted by GLM (with 95% CI) of a wolf scent marking (scats, urine and scratching) in relation to distance to the closest crossroad: a scent marks on roads, b scent marks off-road in the Lower Silesia Forest, western Poland in 2012–2017 (n = 1278 scent marks)
Discussion
Wolves in our study area used forest roads to travel fast and far across their home ranges but spent relatively little time on roads. Almost half of the trail length (revealed by snow tracking) was along roads, which is a higher proportion than in other studies, conducted in areas of lower road density in Canada and Poland (Whittington et al. 2005; Pirga 2011). However, our telemetry results indicated that the proportion of time that wolves spent on roads was similar or lower than reported in areas of lower road density (up to 1.5 km/km2) in Canada, Scandinavia and Finland (Gurarie et al. 2011; Zimmermann et al. 2014; Dickie et al. 2017). This seeming contradiction between wolf preference for roads based on telemetry and snow tracking illustrates how different methods may yield opposing results in wildlife studies. Snow tracking does not account for time; whereas, telemetry data require high-frequency acquisition rates to capture and depict behavioural details such as resting and use of roads. Therefore, while wolves used the roads for fast movement, they preferred to rest or feed away from roads, hence the relatively higher numbers of locations off-road, which was also observed by Zimmermann et al. (2014).
Wolf use of forest roads in our study was related to traffic level and period of day. The strong preference of high-traffic roads detected by snow tracking and higher speed on that roads can be explained by the ease of travel on roads (Dickie et al. 2017). The very strong and time-dependent avoidance of high-traffic roads revealed by telemetry was likely an effect of higher risk of encountering humans on roads, especially during working hours of forestry personnel. Wolves preferred low-traffic roads during working time of forestry personnel and selected high-traffic roads at times of low human activity.
The dense road network in our study area had a strong impact on wolf scent-marking behaviour. Wolves increased rates of all three types of scent marking on roads, contrary to previous studies, in which wolves increased only scratching rates (Zub et al. 2003) or scratching and urination rates (Peters and Mech 1975) on roads. Road junctions seemed to play an important role for urine marking and scratching in our study area; however, contrary to other studies, wolves did not select junctions for scat deposition (Peters and Mech 1975; Zub et al. 2003; Barja et al. 2004). Moreover, scent marking intensity also varied in relation to road category, as wolves preferred to scratch and deposit scats on high-traffic roads. Our study is the first to document differences in response to roads relatively rarely used by humans based on variation in traffic (< 20 vehicles/day).
Wolves in our study area took advantage of dense road network for travel and scent marking while avoiding spending time on them to minimise the probability of encountering humans. Similar ambivalent responses towards infrastructure have been suggested as one of the key factors for the recent wolf recovery in human-dominated landscapes (Zimmermann et al. 2014). The adjustment of the daily pattern of road use to human activity is another example of the behavioural plasticity of wolves in avoiding human encounters. It seems that although wolves in our study area have not been legally hunted for over 20 years, they still might perceive forest roads as sites of elevated risk. Since there is no wildlife mortality caused by collisions with vehicles on forest roads in our study area, we believe that illegal wolf shooting, which is regularly reported in Poland (Gula 2008; Gula et al. 2009; Nowak and Mysłajek 2016), contributes to maintaining this behaviour.
The avoidance of roads of negligible traffic suggests that the effect of roads on wolf behaviour is not solely a function of the level of human activity (Hebblewhite and Merrill 2008). Our results suggest that in areas of very high road density, even roads sporadically used by humans, may influence wolf behaviour. Development and improvement of forestry infrastructure, especially of forest roads, in Poland and several other European countries, have accelerated since the transition to the market economy, and have been fostered further by European funds since the admission to the European Union. The ongoing expansion and improvement of the forest road network might lead to increased human-caused mortality and elevated costs of the avoidance of humans and roads. Therefore, in densely populated countries with fragmented forests, like Poland, forest roads should be considered in wolf habitat assessments, and formally existing bans for non-authorised vehicles on forest roads should be reinforced. Finally, our results show that the high-traffic forest roads offer the highest probability to find wolf scats, which may be helpful in the collection of non-invasive material for molecular analyses (Waits and Paetkau 2005).
Limitations of the study
This study has several limitations related to the small sample size (only three individuals and three packs). First, we were not able to address the inter-individual differences in road use, especially the influence of social status on wolf behaviour, which can be an important determinant of wolf response to roads (Zimmermann et al. 2014). Second, the small number of locations during the spring–autumn period likely contributed to the lack of seasonal patterns in road use and selection in our study. Third, the small study area (and uniform road density within) prevented us from addressing the relationship between road density and road use within wolf home ranges and among wolf packs.
References
Asa CS, Mech LD, Seal US (1985) The use of urine, faeces, and anal-gland secretions in scent-marking by a captive wolf (Canis lupus) pack. Anim Behav 33:1034–1036
Barja I, de Miguel FJ, Bárcena F (2004) The importance of crossroads in faecal marking behaviour of the wolves (Canis lupus). Naturwissenschaften 91:489–492
Bartoń K (2019) Mu-MIn: Multi-model inference. R Package Version 0.12.2/r18. https://R-Forge.R-project.org/projects/mumin/
Bena W (2012) Dzieje Puszczy Zgorzelecko-Osiecznickiej. F.H. Agat, Zgorzelec
Benson JF, Mahoney PJ, Patterson BR (2015) Spatiotemporal variation in selection of roads influences mortality risk for canids in an unprotected landscape. Oikos 124:1664–1673
Coffin AW (2007) From roadkill to road ecology: a review of the ecological effects of roads. J Trans Geogr 15:396–406
Cullingham CI, Thiessen CD, Derocher AE, Paquet PC, Miller JM, Hamilton JA, Coltman DW (2016) Population structure and dispersal of wolves in the Canadian Rocky Mountains. J Mammal 97:839–851
Dickie M, Serrouya R, McNay RS, Boutin S (2017) Faster and farther: wolf movement on linear features and implications for hunting behaviour. J Appl Ecol 54:253–263
Di Orio AP, Callas R, Schaefer RJ (2003) Performance of two GPS telemetry collars under different habitat conditions. Wildl Soc B 31:372–379
Fahrig L, Rytwinski T (2009) Effects of roads on animal abundance: an empirical review and synthesis. Ecol Soc 14:21
Forman RT, Alexander LE (1998) Roads and their major ecological effects. Annu Rev Ecol Syst 29:207–231
Gula R (2008) Legal protection of wolves in Poland: implications for the status of the wolf population. Eur J Wildl Res 54:163–170
Gula R, Hausknecht R, Kuehn R (2009) Evidence of wolf dispersal in anthropogenic habitats of the Polish Carpathian Mountains. Biodivers Conserv 18:2173–2184
Gurarie E, Suutarinen J, Kojola I, Ovaskainen O (2011) Summer movements, predation and habitat use of wolves in human modified boreal forests. Oecologia 165:891–903
Haddad NM, Bowne DR, Cunningham A, Danielson BJ, Levey DJ, Sargent S, Spira T (2003) Corridor use by diverse taxa. Ecology 84:609–615
Hall JS, Harris DJ, Medjibe V, Ashton PMS (2003) The effects of selective logging on forest structure and tree species composition in a Central African forest: implications for management of conservation areas. Forest Ecol Manag 183:249–264
Hebblewhite M, Merrill E (2008) Modelling wildlife–human relationships for social species with mixed-effects resource selection models. J Appl Ecol 45:834–844
Hebblewhite M, White CA, Nietvelt CG, McKenzie JA, Hurd TE, Fryxell JM, Bayley S, Paquet PC (2005) Human activity mediates a trophic cascade caused by wolves. Ecology 86:2135–2144
Heilman GE, Strittholt JR, Slosser NC, Dellasala DA (2002) Forest fragmentation of the conterminous United States: assessing forest intactness through road density and spatial characteristics: forest fragmentation can be measured and monitored in a powerful new way by combining remote sensing, geographic information systems, and analytical software. AIBS Bull 52:411–422
Houle M, Fortin D, Dussault C, Courtois R, Ouellet JP (2010) Cumulative effects of forestry on habitat use by gray wolf (Canis lupus) in the boreal forest. Landsc Ecol 25:419–433
James AR, Stuart-Smith AK (2000) Distribution of caribou and wolves in relation to linear corridors. J Wildl Manag 64:154–159
Jędrzejewski W, Niedziałkowska M, Nowak S, Jȩdrzejewska B (2004) Habitat variables associated with wolf (Canis lupus) distribution and abundance in northern Poland. Divers Distrib 10:225–233
Laurance WF, Goosem M, Laurance SG (2009) Impacts of roads and linear clearings on tropical forests. Trends Ecol Evol 24:659–669
Lesmerises F, Dussault C, St-Laurent MH (2013) Major roadwork impacts the space use behaviour of gray wolf. Landsc Urban Plan 112:18–25
Mancinelli S, Falco M, Boitani L, Ciucci P (2019) Social, behavioural and temporal components of wolf (Canis lupus) responses to anthropogenic landscape features in the central Apennines, Italy. J Zool. https://doi.org/10.1111/jzo.12708
Mills KJ, Patterson BR, Murray DL, Bowman J (2006) Effects of variable sampling frequencies on GPS transmitter efficiency and estimated wolf home range size and movement distance. Wildl Soc B 34:1463–1469
Nowak S, Mysłajek RW (2016) Wolf recovery and population dynamics in Western Poland, 2001–2012. Mamm Res 61:83–98
Nowak S, Mysłajek RW, Kłosińska A, Gabryś G (2011) Diet and prey selection of wolves (Canis lupus) recolonising Western and Central Poland. Mamm Biol 76(6):709–715
Person DK, Russell AL (2008) Correlates of mortality in an exploited wolf population. J Wildl Manag 72:1540–1549
Peters RP, Mech LD (1975) Scent-marking in wolves: Radio-tracking of wolf packs has provided definite evidence that olfactory sign is used for territory maintenance and may serve for other forms of communication within the pack as well. Am Sci 63:628–637
Pirga B (2011) Wpływ czynników antropogenicznych na strukturę i użytkowanie areałów przez wilki. Ph.D. Dissertation, Institute of Nature Conservation, Polish Academy of Sciences, Kraków, Poland (in Polish)
Potvin MJ, Drummer TD, Vucetich JA, Beyer DE, Peterson RO, Hammill JH (2005) Monitoring and habitat analysis for wolves in upper Michigan. J Wildl Manag 69:1660–1669
Quantum GIS Development Team (2018) Quantum GIS Geographic Information System. Open Source Geospatial Foundation Project. https://qgis.osgeo.org
R Core Team (2019) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Recio MR, Zimmermann B, Wikenros C, Zetterberg A, Wabakken P, Sand H (2018) Integrated spatially-explicit models predict pervasive risks to recolonizing wolves in Scandinavia from human-driven mortality. Biol Conserv 226:111–119
Reimoser F, Gossow H (1996) Impact of ungulates on forest vegetation and its dependence on the silvicultural system. For Ecol Manag 88:107–119
Reed RA, Johnson-Barnard J, Baker WL (1996) Contribution of roads to forest fragmentation in the Rocky Mountains. Conserv Biol 10:1098–1106
Rothman RJ, Mech LD (1979) Scent-marking in lone wolves and newly formed pairs. Anim Behav 27:750–760
Saunders SC, Mislivets MR, Chen J, Cleland DT (2002) Effects of roads on landscape structure within nested ecological units of the Northern Great Lakes Region. USA Biol Conserv 103:209–225
Suárez-Esteban A, Delibes M, Fedriani JM (2013) Barriers or corridors? The overlooked role of unpaved roads in endozoochorous seed dispersal. J Appl Ecol 50:767–774
Theuerkauf J, Jȩdrzejewski W, Schmidt K, Gula R (2003) Spatiotemporal segregation of wolves from humans in the Białowieża Forest (Poland). J Wildlife Manag 67:706–716
Theuerkauf J, Gula R, Pirga B, Tsunoda H, Eggermann J, Brzezowska B, Rouys S, Radler S (2007) Human impact on wolf activity in the Bieszczady Mountains, SE Poland. Ann Zool Fenn 44:225–231
Trombulak SC, Frissell CA (2000) Review of ecological effects of roads on terrestrial and aquatic communities. Conserv Biol 14:18–30
Wabakken P, Sand H, Liberg O, Bjärvall A (2001) The recovery, distribution, and population dynamics of wolves on the Scandinavian peninsula, 1978–1998. Can J Zool 79:710–725
Waits LP, Paetkau D (2005) Noninvasive genetic sampling tools for wildlife biologists: a review of applications and recommendations for accurate data collection. J Wildl Manag 69:1419–1433
Whittington J, St Clair CC, Mercer G (2005) Spatial responses of wolves to roads and trails in mountain valleys. Ecol Appl 15:543–553
Whittington J, Hebblewhite M, DeCesare NJ, Neufeld L, Bradley M, Wilmshurst J, Musiani M (2011) Caribou encounters with wolves increase near roads and trails: a time-to-event approach. J Appl Ecol 48:1535–1542
Wolsan M, Bieniek M, Buchalczyk T (1992) The history, and distributional and numerical changes of the wolf Canis lupus (L.) in Poland. In: Proceedings of the 18th IUGB Congress, Kraków
Zimmermann B, Nelson L, Wabakken P, Sand H, Liberg O (2014) Behavioural responses of wolves to roads: scale-dependent ambivalence. Behav Ecol 25:1353–1364
Zub K, Theuerkauf J, Jędrzejewski W, Jędrzejewska B, Schmidt K, Kowalczyk R (2003) Wolf pack territory marking in the Białowieża primeval forest (Poland). Behaviour 140:635–648
Acknowledgements
This study was funded by the Polish National Science Centre (Research Grants NCN2011/01/B/NZ8/04233 and NCN2012/05/N/NZ8/00860) and by the SAVE Wildlife Conservation Fund. We thank students, volunteers and members of forestry personnel of Ruszów, Wymiarki and Pieńsk for their support during field work.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
All authors have participated in (a) conception and design, or analysis and interpretation of the data; (b) drafting the article or revising it critically for important intellectual content; and (c) approval of the final version.
Additional information
Handling Editor: Luca Corlatti.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bojarska, K., Sulich, J., Bachmann, S. et al. Opportunity and peril: how wolves use a dense network of forest roads. Mamm Biol 100, 203–211 (2020). https://doi.org/10.1007/s42991-020-00014-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-020-00014-0