
Abstract
Common in Fungal Extracellular Membrane (CFEM) domain proteins are considered to be unique to fungi and closely related to pathogenicity. However, the Puccinia striiformis f. sp. tritici (Pst) effector containing the CFEM domain has not been reported. Here, we obtained an effector, PstCFEM1, containing a functional N-terminal signal peptide sequence and the CFEM domain from Pst race CYR31. qRT-PCR assay indicated that the transcript levels of PstCFEM1 were highly induced during the early stages of infection. Overexpression of PstCFEM1 suppressed Pst322 (an elicitor-like protein of Pst)-trigged cell death, reactive oxygen species (ROS) accumulation and callose deposition. Host-induced gene silencing (HIGS) experiments showed that knockdown of PstCFEM1 decreased the virulence of Pst, while ROS accumulation in silenced plants increased near the infection site. In addition, wheat containing the PstCFEM1-silenced construct increased resistance to multiple races of Pst. Our data suggest that PstCFEM1 suppresses wheat defense by inhibiting ROS accumulation and contributes to increased virulence of Pst.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wheat stripe rust, caused by the basidiomycetous fungus Puccinia striiformis f. sp. tritici (Pst), one of the most common and destructive wheat diseases in most wheat growing areas (Wellings, 2011; Chen et al., 2014), has been one of the most significant biotic threats to global wheat production in the twenty-first century (Schwessinger, 2017). Present strategies to manage this disease depend on resistant cultivars combined with fungicide application (Chen et al., 2014). Hence, new viable methods must be discovered to protect wheat crops from rust fungi (Fisher et al., 2012). Understanding the molecular basis of Pst pathogenesis and the Pst–wheat interaction will contribute new strategies for long-term stripe rust control (Brown, 2015; Chen et al., 2014).
Pst is an obligate biotrophic parasite that cannot be cultured in vitro (Staples, 2000) and can only invade living host tissues, where it forms haustoria that invade the host cells to absorb nutrients (Voegele & Mendgen, 2011). Because Pst lacks a reliable and efficient transformation mechanism, the function of only a few effectors has been studied (Petre et al., 2014). The use of the host-induced gene silencing (HIGS) technique has greatly aided the study of Pst effectors and research related to pathogenicity (Yin & Hulbert, 2015; Qi et al., 2019b). Pst18363, a stripe rust effector, targets and stabilizes TaNUDX23 that encourages stripe rust disease (Yang et al., 2020). To overcome reactive oxygen species (ROS)-induced defense in wheat, Pst effector PstGSRE1 interrupts nuclear localization of ROS-promoting transcription factor TaLOL2 (Qi et al., 2019a). An effector protein Pst_12806 of Pst targets the wheat TaISP protein (a putative component of the cytochrome b6-f complex) and suppresses chloroplast function (Xu et al., 2019). Pst_8713 is an effector that impairs plant immunity and enhances Pst virulence (Zhao et al., 2018). Effector PEC6 overwhelms pattern-triggered immunity (PTI) in a host species-independent manner (Liu et al., 2016). However, there have been no reports indicating that silencing of effector gene confers broad-spectrum resistance of wheat against Pst.
Fungal extracellular membrane proteins commonly include the CFEM domain (Zhang et al., 2015), a protein domain unique to fungi, containing distinctive eight cysteine residues (Kulkarni et al., 2003). Many studies have shown that the CFEM protein is closely linked to fungal pathogenicity. CFEM domain protein MoPTH11 of Magnaporthe oryzae functions as an intracellular signal molecule in appressoria and bud tracheids which is required for proper development of the appressoria, and appressoria-like structures (Kou et al., 2017). CFEM domain protein MoACI1 directly affects the development of appressoria (Kulkarni & Dean, 2004; Salomon et al., 2014). CFEM domain protein MoWISH is essential for surface sensing, asexual and pathogenic differentiation in M. oryzae (Sabnam & Barma, 2017). CFEM domain proteins CSA1 of Candida albicans and AG2 of Coccidioides immitis regulate the growth of hyphae and pathogenicity (Peng et al., 1999; Lamarre et al., 2000). Pathogenicity, conidial development, and stress tolerance are all influenced by BcCFEM1 in Botrytis cinerea (Zhu et al., 2017). However, an effector containing the CFEM domain in the stripe rust fungus has not been documented.
In the current study, we isolated and characterized a candidate effector PstCFEM1 from Pst and used overexpression and HIGS technologies to explore its function. Our results indicated that PstCFEM1 facilitates Pst infection by suppressing ROS accumulation and wheat plants containing the PstCFEM1-silenced construct show broad-spectrum resistance to Pst.
Results
Identification of the candidate effector
A number of Pst secreted proteins have been identified (Zheng et al., 2013). In our previous study using an A. tumefaciens-mediated transient expression assay in N. benthamiana, 30 effectors were selected by Agrobacterium carrying mouse pro-apoptotic protein-bax (Yang et al., 2020). A candidate effector PstCFEM1 was shown to suppress bax-triggered programmed cell death (PCD) (Fig. S1). PstCFEM1 was cloned from Pst race CYR31 and found to have an open reading frame of 579 bp, encoding a protein of 192 amino acids. The sequence of this protein contains a CFEM domain and is consistent with the sequence of hypothetical protein PSTG_04849 through NCBI Blastp (GenBank accession number KNF02028.1). Since there is no report of this type of effector in Pst, we named this protein “PstCFEM1” as this is the first report of a CFEM protein in Pst.
PstCFEM1 is significantly induced by Pst infection
To determine whether PstCFEM1 is involved in Pst infection, a qRT-PCR assay was used to analyze PstCFEM1 transcript levels at different Pst infection stages. The transcript levels were gradually induced as early as 6 h post inoculation (hpi) and attained the maximum level of 11.5-fold at 48 hpi compared with the control. Then, the transcript level returned to the original level at 72 hpi (Fig. 1). This result indicates that transcription of PstCFEM1 is induced by Pst infection.

Transcript profiles of PstCFEM1 in different Pst infection stages. Wheat leaves inoculated with compatible race CYR31 were sampled at 0, 6, 12, 18, 24, 48, 72 and 120 hpi. Error bars represent the variations among three independent replicates. qRT-PCR values were normalized using TaEF-1α. Differences between time-course points were assessed using student’s t-test (*, P < 0.05; **, P < 0.01)
Secretion validation of the N-terminal signal peptide of PstCFEM1
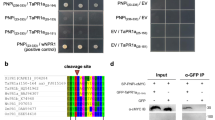
SignalP 5.0 predicted that PstCFEM1 has a signal peptide encoded by the first 75 bp (Fig. S2). To confirm the secretory function of the predicted signal peptide of PstCFEM1, we used a signal sequence trap system (Zhao et al., 2018). Empty vector pSUC2 was used as a negative control, and the signal peptide of effector Avr1b from Phytophthora sojae was used as a positive control (Gu et al., 2011). Similar to signal peptide of Avr1b, the signal peptide of PstCFEM1 enabled the invertase mutant yeast strain to grow on CMD-W medium (yeast growth without invertase secretion) and YPRAA medium (yeast growth only when invertase is secreted) (Fig. 2). In addition, to further confirm the secretory function of the signal peptide, we tested invertase enzymatic activity, in which secreted invertase reduced TTC to insoluble red colored TPF. We found that the TTC-treated signal peptides of PstCFEM1 and positive control culture filtrates turned red, whereas the negative control culture filtrates treated with TTC remained colorless (Fig. 2). This result verified the secretory function of N-terminal signal peptide of PstCFEM1.

Functional validation of the signal peptide of PstCFEM1. A The sequence of the putative PstCFEM1 signal peptide was fused in-frame to the invertase sequence in the pSUC2 vector and transformed into yeast strain YTK12. The untransformed YTK12 strain and empty pSUC2 vector were used as negative controls, and the oomycete effector Avr1b from Phytophthora sojae was used as a positive control. Only yeast strains that are able to secrete invertase can grow on YPRAA media. TTC is reduced to red formazan when invertase is secreted. B Western blot of PstCFEM1 in intercellular fluid. Cytoplasmic effector Pst18363 from Pst as negative control. Three independent biological replicates were conducted for each experiment
To further investigate its localization, we firstly performed bioinformatic analysis. The results indicated that PstCFEM1 has no predicted transmembrane region (Fig. S3) but has a predicted GPI-anchor site (Fig. S4), and it’s a putative apoplastic effector (Fig. S5). To verify the prediction, PstCFEM1 protein in the intercellular fluid of N. benthamiana was detected using the western blot method. As shown in Fig. 2B, PstCFEM1 was present in the intercellular fluid, but the GFP protein was not detected. In addition, Pst18363 which had been identified as a cytoplasmic effector (Yang et al., 2020) was not been detected in the intercellular fluid (Fig. 2B). Thus, the results suggest that PstCFEM1 is an apoplastic effector.
PstCFEM1 suppresses PCD and callose deposition
To explore the mechanisms by which PstCFEM1 contributes to pathogenicity, we investigated whether, as other fungal effectors, PstCFEM1 could suppress PCD in N. benthamiana. In this study, we chose the Pst candidate elicitor-like protein Pst322 as a trigger for cell death (Wang et al., 2012) and Pst18363 was used as a positive control to suppress PCD (Yang et al., 2020). The leaves were infiltrated with A. tumefaciens cells containing PstCFEM1, Pst18363 and GFP, then A. tumefaciens cells carrying Pst322 were infiltrated in the same position after 24 h. After 5 d of the final infiltration, no cell death symptoms were observed on the leaf pre-infiltrated with A. tumefaciens carrying PstCFEM1 and Pst18363, whereas Pst322-induced cell death symptoms were evident in regions pre-infiltrated with A. tumefaciens carrying GFP (Fig. 3A). As expected, A. tumefaciens carrying PstCFEM1 and GFP alone did not elicit cell death (Fig. 3A). The results indicated that overexpression of PstCFEM1 suppressed Pst322-trigged cell death. We further investigated the function of PstCFEM1 in suppressing plant immunity by using the same procedure in N. benthamiana. After 24 h of the final infiltration, we observed a lower level of callose deposition in PstCFEM1 and Pst322 infiltrated leaves compared to the tobacco leaves with GFP and Pst322 infiltrated leaves (Fig. S6A). In addition, RT-PCR and western blot assays confirmed the normal expression of genes (GFP, Pst18363, PstCFEM1 and Pst322) in the infiltrated regions (Fig. S6B, C).

Overexpression of PstCFEM1 suppressed programmed cell death and callose deposition. A PstCFEM1 suppressed Pst322-triggered cell death. A. tumefaciens cells respectively carrying the Pst18363, PstCFEM1 and eGFP vector were infiltrated into the leaf of N. benthamiana within the regions indicated by the dashed lines, followed after 24 h by either no further infiltration (left side) or infiltration with A. tumefaciens cells carrying the Pst322 (right side). The phenotype of cell death was scored and photographed at 5 d after infiltration with Pst322, and the leaves were decolorized with ethanol. B The wheat leaves inoculated with MgCl2 buffer, EtHAn, pEDV6:RFP and pEDV6:PstCFEM1 were examined for callose deposition by epifluorescence microscopy after aniline blue staining. Bar = 100 μm. C Average number of callose deposits in wheat leaves inoculated with MgCl2, EtHAn, pEDV6:RFP and pEDV6:PstCFEM1. Data represent the mean of three biological replicates. Different letters indicate significant differences (one-way ANOVA, Tukey’s Multiple Comparison Test, p < 0.05)
It has been reported that the effector detector vector (EDV) can deliver individual non-bacterial effectors to host plant cells via the bacterial TTSS (Sohn et al., 2007). Therefore, PstCFEM1 was also cloned into the expression vector pEDV6, and then delivered into wheat cv. Suwon11 by the modified P. fluorescens strain EtHAn, which carries a functional TTSS (Thomas et al., 2009; Yin & Hulbert, 2011). Callose deposition was observed on wheat leaves after inoculating with EtHAn (Fig. 3B), and the EtHAn strain-carrying RFP was used as the negative control. We observed a decrease in callose deposition in pEDV6:PstCFEM1-inoculated wheat plants (Fig. 3B). Next, we measured the callose deposition in the infiltrated wheat leaves and found that callose deposition in PstCFEM1-infiltrated leaves significantly decreased compared to that in leaves infiltrated with the EtHAn strain carrying pEDV6:RFP (Fig. 3C). These results indicated that overexpression of PstCFEM1 suppresses PCD and callose deposition, and influences the virulence of PstCFEM1.
Silencing of PstCFEM1 weakens the virulence of Pst
The HIGS technique has facilitated research on Pst effectors (Qi et al., 2019a; Xu et al., 2019). In this study, HIGS mediated by Barley Stripe Mosaic Virus (BSMV) was used to knock down the transcripts of PstCFEM1. Two fragments were selected in the coding region to specifically silence PstCFEM1. The second leaves of Suwon11 seedlings were inoculated with BSMV:TaPDS (wheat phytoene desaturase gene)-as, BSMV:PstCFEM1–1as/2as antisense (PstCFEM1-silenced) and BSMV:γ (control). A similar number of seedlings were also mock-inoculated with a buffer lacking BSMV. The photo-bleaching phenotype was observed when TaPDS was silenced (Fig. 4A). All wheat plants containing the PstCFEM1-silenced construct and control plants expressed mild chlorotic mosaic symptoms, confirming that the BSMV-mediated gene silencing system functioned correctly and could be used in further experiments (Fig. 4A). To clarify whether PstCFEM1 was successfully silenced, transcript levels of PstCFEM1 were examined by qRT-PCR assay, which indicated that its transcript was noticeably decreased at 24, 48, and 120 hpi (Fig. 4B). The fourth leaves of wheat plants were then inoculated with the virulent Pst isolate CYR31. The phenotypes showed that wheat plants containing the PstCFEM1-silenced construct had reduced sporulation (Fig. 4C) and they had significantly reduced biomass (Fig. 4D) compared to control plants.

BSMV-mediated HIGS of PstCFEM1 weaken the pathogenicity of Pst. A Mild chlorotic mosaic symptoms were observed on leaves inoculated with BSMV:γ, BSMV:TaPDS-as and BSMV:PstCFEM1–1as/2as at 10 dpi. Mock, wheat leaves treated with 1 × Fes buffer. B Relative expression of PstCFEM1 during the interaction between wheat containing the PstCFEM1-silenced construct and CYR31. qRT-PCR values were normalized using PstEF1, and were presented as fold changes relative to that in plants with BSMV:γ treatment at time 0. C Photos of fourth leaves of wheat containing the PstCFEM1-silenced construct inoculated with fresh urediospores of the CYR31 race. Typical leaves were photographed at 14 dpi. D Ratio of fungal to wheat nuclear content determined using the contents of fungal PstEF1 and wheat TaEF-1α genes, respectively. Genomic DNA was extracted from three different plants at 7 dpi. Data represent the mean of three biological replicates. Asterisks indicate significant differences between that in silenced plants and control plants at the same time points using student’s t-test (*, P < 0.05)
Based on the phenotypic variation of wheat containing the PstCFEM1-silenced construct inoculated with Pst, we assessed the growth of Pst. At 48 hpi, the colony area per infection site was obviously smaller in wheat containing the PstCFEM1-silenced construct compared with control plants (Fig. 5A, B), indicating that PstCFEM1 impairs Pst growth. To further confirm the suppressed Pst growth in wheat containing the PstCFEM1-silenced construct, the transcripts of pathogenesis-related (PR) marker genes (TaPR1 and TaPR2) (Liu et al., 2019), were examined by qRT-PCR assay. The transcript levels of TaPR1 and TaPR2 were significantly induced in wheat containing the PstCFEM1-silenced construct during Pst infection (Fig. 5C, D).

Histological observation of fungal growth and transcript levels of TaPR genes in wheat containing the PstCFEM1-silenced construct compared to control plant. A The fungal structures were stained with WGA in wheat leaves inoculated with Pst and observed under a fluorescence microscope. SV, sub-stomatal vesicle; HMC, haustorial mother cell; IH, infection hypha; H, haustoria. B The Infection area were calculated using DP-BSW software. All results were calculated from 30 to 40 infection sites. Transcript levels of (C) TaPR1 and (D) TaPR2 in PstCFEM1-silenced compared to control plants. qRT-PCR values were normalized with TaEF-1α. Error bar represents the variation among three independent replicates. All experiments were performed with three biological replicates. Asterisks indicate significant differences between that in silenced plants and control at the same time points using student’s t-test (*, P < 0.05; **, P < 0.01)
Silencing of PstCFEM1 increases ROS accumulation during Pst-wheat interaction
To examine the host responses in wheat containing the PstCFEM1-silenced construct, we measured ROS accumulation by staining the infected wheat tissues at 48 hpi with DAB. H2O2 area per infection site in wheat containing the PstCFEM1-silenced construct was significantly increased compared with control plants (Fig. 6A, B). Meanwhile, the transcripts of two ROS-related marker genes (TaCAT3 and TaNOX) (Hawku et al., 2021; Liu et al., 2019), were examined by qRT-PCR assay. TaCAT3 was significantly decreased at 24 and 48 hpi in wheat containing the PstCFEM1-silenced construct compared with control plants (Fig. 6C), TaNOX exhibited a significantly increased transcript at 48 hpi in wheat containing the PstCFEM1-silenced construct compared with control plants (Fig. 6D). These results suggest that silencing of PstCFEM1 induces host ROS accumulation.

Production of ROS in control and wheat containing the PstCFEM1-silenced construct challenged with CYR31. A H2O2 production was observed by DAB staining. H2O2 accumulation and fungal growth in BSMV-silenced leaves at 48 hpi. Bars = 20 μm. SV, sub-stomatal vesicle; HMC, haustorial mother cell; IH, infection hypha and H, haustorium. B The H2O2 areas were calculated using DP-BSW software. All results were calculated from 30 to 40 infection sites. Transcript levels of (C) TaCAT3 and (D) TaNOX in PstCFEM1-silenced compared to control plants. qRT-PCR values were normalized for TaEF-1α. Error bar represents the variation among three independent replicates. All experiments were performed with three biological replicates. Asterisks indicate significant differences between that in silenced plants and control plants at the same time points using student’s t-test (*, P < 0.05; **, P < 0.01)
PstCFEM1 inhibits ROS accumulation in N. benthamiana
Because ROS is a crucial trigger of cell death and to test if PstCFEM1 suppresses Pst322-triggered cell death by preventing ROS accumulation, DAB staining was used to examine the ROS levels in infiltrated leaves of N. benthamiana. RT-PCR and western blot assays confirmed the normal expression of genes (GFP, PstCFEM1 and Pst322) in the infiltrated regions (Fig. 7A, B). The DAB staining in the leaf regions infiltrated with PstCFEM1/Pst322 was much weaker compared with those infiltrated with buffer/Pst322 and GFP/Pst322 (Fig. 7C). Meanwhile, we used qRT-PCR assay to test the transcript levels of two ROS-scavenging genes (NbSOD and NbCAT) (Guo et al., 2018). NbSOD (Fig. 7D) and NbCAT (Fig. 7E) exhibited a significantly increased transcript level at 1 d in the PstCFEM1/Pst322 region compared with the GFP/Pst322 region. These results indicated that PstCFEM1 suppresses Pst322-triggered cell death by preventing ROS accumulation.

PstCFEM1 decreases ROS accumulation. RT-PCR (A) and western blot (B) was performed to confirm the expression of PstCFEM1, GFP and Pst322 in N. benthamiana. Nbactin was used as a reference gene. C H2O2 production in N. benthamiana leaves was determined by DAB staining. N. benthamiana leaves were infiltrated with A. tumefaciens cells carrying the PstCFEM1 or GFP, followed after 24 h by infiltration with A. tumefaciens cells carrying the Pst322. The measurement was performed 2 d after infiltration with Pst322. Transcript levels of (D) NbSOD and (E) NbCAT in PstCFEM1 + Pst322 region compared to GFP + Pst322 region at 1d after infiltration with Pst322. qRT-PCR values were normalized with Nbactin. Error bar represents the variation among three independent replicates. All experiments were performed with three biological replicates. Differences between PstCFEM1 + Pst322 region and GFP + Pst322 region were assessed using student’s t-test (*, P < 0.05; **, P < 0.01)
Wheat plants containing the PstCFEM1-silenced construct express broad-spectrum resistance to Pst
HIGS employing RNA silencing mechanism and, specifically, silencing the targets of invading pathogens, has been successfully applied in crop disease prevention (Qi et al., 2019b). Because the current prevalent Pst races in China are CYR32, CYR33, and CYR34 (Liu et al., 2017), we cloned the PstCFEM1 gene in those three races and found by sequence analysis that PstCFEM1 is conserved in different races (Fig. S7). We used the same method above to silence PstCFEM1, and found that both wheat plants containing the PstCFEM1-silenced construct and control plants were susceptible to CYR32 (Fig. 8A), CYR33 (Fig. 8B), and CYR34 (Fig. 8C), Compared with control plants, fungal biomass was significantly reduced in wheat containing the PstCFEM1-silenced construct (Fig. 8A-C). Thus, our results indicate that silencing of PstCFEM1 confers broad-spectrum resistance of wheat to Pst.

PstCFEM1-silenced plants expressed broad-spectrum resistance to Pst. Foliar parts of BSMV:PstCFEM1–1as/2as plants inoculated with Pst isolate CYR32 (A), CYR33 (B), and CYR34 (C). Ratio of fungal to wheat nuclear DNA content was determined using the contents of fungal PstEF1 and wheat TaEF-1α genes, respectively. Genomic DNA was extracted from three different plants at 7 days post inoculation. Error bars represent the variation among three independent replicates. Asterisks indicate significant differences between that in silenced plants and control plants using student’s t-test (*, P < 0.05)
Discussion
CFEM domains are unique to fungi and originated from the common ancestor of Ascomycota and Basidiomycota (Zhang et al., 2015). The original function of CFEM domains is cell wall/membrane constitution, but divergence from normal functioning facilitates other various roles, such as pathogenicity (Zhang et al., 2015). MoPTH11 is an important G-protein-coupled receptor (GPCR) containing seven transmembrane domains (Kou et al., 2016; Weis et al., 2018), and the CFEM protein CSA1 of Candida albicans and AG2 of Coccidioides immitis also are membrane proteins (Peng et al., 1999; Lamarre et al., 2000). CFEM proteins without transmembrane structure usually comprise GPI-anchored sites, such as CFEM-domain GPI-anchored proteins (CfmA-C) and BcCFEM1 (Vaknin et al., 2014; Zhu et al., 2017). In plants, GPI-anchored proteins are involved in regulation of cell expansion and cell wall biosynthesis (Brady et al., 2007). Several studies have indicated that GPI-anchored proteins attach to the plasma membrane and play an important role in maintaining the cell wall and stress tolerance (Kitagaki et al., 2002). In this study, we found that PstCFEM1 has a predicted GPI-anchor site (Fig. S4), and it’s an apoplastic effector (Fig. S5; Fig. 2B). Apoplastic effectors play roles in extracellular spaces and cope with physical and chemical barriers to break the first line of plant defenses (Giraldo et al., 2013; Tariqjaveed et al., 2021). Meanwhile, PstCFEM1 was proven to be a virulence effector in this study. Therefore, we propose that PstCFEM1 is a potential GPI-anchored CFEM protein, which is anchored to the outer layer of the cell membrane through a C-terminal GPI anchor, and it’s an apoplastic effector in the host apoplast during Pst-wheat interaction.
The CFEM domain has been reported to regulate mycelial growth, appressorium formation and pathogenicity (Salomon et al., 2014; Kou et al., 2016), but some CFEM proteins are avirulence factors or perform other functions. The three Aspergillus fumigatus CFEM-domain GPI-anchored proteins affect cell-wall stability but do not perform a role in fungal virulence (Vaknin et al., 2014). MoCDIP2 in the rice blast fungus is the only CFEM effector that can induce PCD (Chen et al., 2013), and we speculate that it may act as an elicitor-like protein. In Fusarium graminearum, none of the CFEM-containing GPCRs is essential for infection (Jiang et al., 2019). This indicates that functions of CFEM proteins may be diverse. In this study, we identified a CFEM domain protein PstCFEM1 from Pst that suppresses Pst322-induced cell death (Fig. 3A). Through transient overexpression in wheat, we found that it can inhibit callose accumulation (Fig. 3B, C), suggesting that PstCFEM1 is a virulence effector. In addition, a previous study reported that CFEM protein possesses antioxidant properties in M. oryzae (Kou et al., 2017). A similar result, that PstCFEM1 can affect ROS accumulation in host plants, was also observed in this study (Fig. 6, 7).
Pathogens secrete effectors into host cells and manipulate the host to promote their infection and colonization (Presti et al., 2015). At present, a variety of effectors have been known in different rust fungus. AvrM, AvrL567, Avr123 and AvrP4 have been reported in Uromyces striatus (Kemen et al., 2005), and the transfer protein RTP was identified in Uromyces fabae (Kemen et al., 2013; Pretsch et al., 2013). Pst_8713, Pst18363, PstGSRE1, Pst_12806 and PEC6 were identified in Pst (Liu et al., 2016; Zhao et al., 2018; Xu et al., 2019; Qi et al., 2019a; Yang et al., 2020). These identified effectors are secreted proteins with unknown functions. In this study, we found a CFEM-containing protein in Pst that is a virulence effector suppressing accumulation of ROS in wheat and promoting the growth of Pst. Because the length and architecture of CFEM domains are relatively conserved (Zhang et al., 2015), we speculate that the orthology of PstCFEM1 in other rust fungi also has a similar function. In addition, Pst is a biotrophic, obligate parasite. It invades the host cell through the haustorium, which contributes in the exchange of nutrition and signals (Rafiqi et al., 2012). Moreover, because rust fungi cannot be cultured in vitro and cannot be genetically transformed, their study is relatively lagging (Voegele & Mendgen, 2011). Bacterial TTSS-mediated overexpression and HIGS technology have provided effective methods for the identification and functional research of rust effectors (Zhao et al., 2018; Qi et al., 2019b). In summary, our study provides novel insights into CFEM-containing proteins in host-pathogen interactions.
Conclusion
In summary, our study revealed that PstCFEM1 suppresses wheat defense by inhibiting ROS accumulation and contributes to pathogenicity of Pst. In addition, wheat plants containing the PstCFEM1-silenced construct showed increased resistance to multiple races of Pst. This is the first evidence indicating that silencing a vital CFEM domain-containing protein gene confers broad-spectrum resistance to wheat stripe rust.
Methods
Plant materials and fungal isolates
Wheat cultivar Suwon11 was used for gene transcription analysis and HIGS assays (Liu et al., 2019). This cultivar, carrying the YrSu gene, is highly susceptible to CYR31 (Cao et al., 2003). Wheat seedlings were inoculated with Pst and maintained according to the procedures and conditions previously described (Wang et al., 2007). For RNA extraction, the second leaves inoculated with CYR31 or treated with sterile distilled water (control) were harvested at 0, 6, 12, 18, 24, 48, 72, and 120 hpi. Pst races CYR31, CYR32, CYR33 and CYR34 were used in this study. Suwon11 showed compatible interaction with Pst races CYR31, CYR32, CYR33, and CYR34.
RNA extraction and qRT-PCR assays
The Quick RNA Isolation Kit (Huayueyang Biotechnology, China, Beijing) was used to extract RNA from all samples. The HiScript® QRT SuperMix for qPCR (Vazyme, Nanjing, China) was used for reverse transcription with 2 mg of total RNA. qRT-PCR was performed on a CFX Connect Real-Time System (Bio-Rad, USA). Table S1 in the supporting information contains a list of the primers used. PstEF1 and TaEF-1α were used to normalize values obtained with the qRT-PCR assay (Bai et al., 2021).
Yeast signal sequence trap system
The yeast signal sequence trap method was used as previously described (Yin et al., 2018). Using specific primers, we cloned the predicted signal peptide sequence of PstCFEM1 into vector pSUC2 (Table S1). Then it was transformed into YTK12, an invertase mutant yeast strain (Oh et al., 2009). Invertase enzymatic activity was detected by reduction of TTC to insoluble red formazon TPF according to the procedures and conditions previously described (Jacobs et al., 1997). The specific method is the same as used in our previous study (Yang et al., 2020).
Agrobacterium tumefaciens infiltration assays
The suppression of BAX- or Pst322-triggered PCD by PstCFEM1 was tested with the A. tumefaciens-mediated transient expression system (Zhao et al., 2018). The PVX-PstCFEM1 plasmid was constructed by amplifying the sequence encoding PstCFEM1 minus the signal peptide with primers listed in Table S1. N. benthamiana leaves were infected with resuspended Agrobacterium carrying eGFP or Pst18363 or PstCFEM1 at a final OD600 of 0.2 and 10 mM MgCl2 buffer (Bos et al., 2006). The inoculation sites were infiltrated with A. tumefaciens carrying BAX or Pst322 at a final OD600 of 0.2 in the same position at 24 hpi. The infiltrated N. benthamiana plants were placed in a glasshouse at a temperature of 25 °C in high intensity light. Three days after infiltration, RT-PCR and western blot was used to detect gene expression in all infiltration sites. At 4–5 days after infiltration, symptoms were observed and recorded. For each experiment, three biological replicates were carried out independently.
The apoplastic fluid from N. benthamiana leaves was extracted by the infiltration-centrifugation method as described (Nie et al., 2019). We cloned the PstCFEM1 sequence into vector pCAMBIA1302 using specific primers (Table S1) and N. benthamiana leaves were infected with resuspended Agrobacterium carrying empty vector or PstCFEM1-GFP or Pst18363-GFP at a final OD600 of 0.2 and 10 mM MgCl2 buffer. At 2–3 days after infiltration, the samples were extracted.
Bacterial TTSS-mediated overexpression in wheat plants
Electroporation was used to introduce the pEDV6:PstCFEM1 construct into the P. fluorescens strain EtHAn. Bacterial cells expressing PstCFEM1 were cultured overnight at 28 °C in KB medium (Yin & Hulbert, 2011), then collected and resuspended in an infiltration medium for transient expression in wheat cells (10 mM MgCl2). The P. fluorescens strain EtHAn was diluted to an OD600 of 0.6 and infiltrated using a syringe without a needle into the second leaves of wheat cultivar Suwon11. The infiltrated wheat plants were maintained at a constant temperature of 25 °C in a growth chamber. The wild-type strain EtHAn was used to trigger PTI. As a negative control, wheat leaves were infiltrated with recombinant EtHAn carrying the RFP. Leaf samples were stained with aniline blue to analyze the suppression of callose deposition as previously described (Hood & Shew, 1996). An Olympus BX-51 microscope was used to analyze the specimens (Olympus, Japan). All experiments were repeated three times.
BSMV-mediated PstCFEM1 gene silencing
Two specific cDNA fragments of PstCFEM1 were obtained for HIGS analysis, we confirmed that the fragments showed nonsimilarity with any other genes by Si-Fi software analysis of the wheat and Pst databases. BSMV constructs (BSMV:PstCFEM1–1as / 2as) were used to inoculate wheat seedlings. The control for the BSMV infection test was BSMV:TaPDS, which contained the wheat phytoene desaturase (PDS) gene. Mock inoculations were performed with 1 × FES buffer (Yang et al., 2020). At 9–12 days after virus inoculation, the fourth leaves were inoculated with fresh urediniospores. At 14–16 days after Pst inoculation, the phenotypes of the fourth leaves were examined and photographed, and the leaves were collected for RNA extraction at 24, 48, and 120 hpi (Qi et al., 2018). The silencing efficiencies of PstCFEM1 were assessed by qRT-PCR. Fungal biomass was measured as previously studied (Qi et al., 2018). Pst growth and H2O2 accumulation in wheat plants were analyzed using histological tools. H2O2 was detected using the DAB staining process. An Olympus BX-51 microscope was used to analyze the stained samples (Olympus, Japan). Decolorized leaf segments were stained with wheat germ agglutinin (WGA) conjugated to Alexa-488 (Invitrogen, USA). Three biological replicates were used in this assay.
Availability of data and materials
All data and materials are available in the paper and online supplemental files.
References
Bai XX, Huang XL, Tian SX, Peng H, Zhan GM, Goher F, Guo J, Kang ZS, Guo J (2021) RNAi-mediated stable silencing of TaCSN5 confers broad-spectrum resistance to Puccinia striiformis f. sp. tritici. Mol Plant Pathol 22(4):410–421. https://doi.org/10.1111/mpp.13034
Bos JIB, Kanneganti T, Young C, Cakir C, Huitema E, Win J, Miles R, Armstrong MR, Birch PRJ, Kamoun S (2006) The C-terminal half of Phytophthora infestans RXLR effector AVR3a is sufficient to trigger R3a-mediated hypersensitivity and suppress INF1-induced cell death in Nicotiana benthamiana. Plant J 48(2):165–176. https://doi.org/10.1111/j.1365-313X.2006.02866.x
Brady SM, Song S, Dhugga KS, Rafalski JA, Benfey PN (2007) Combining expression and comparative evolutionary analysis: the COBRA gene family. Plant Physiol 143(1):172–187. https://doi.org/10.1104/pp.106.087262
Brown JKM (2015) Durable resistance of crops to disease: a darwinian perspective. Annu Rev Phytopathol 53(1):513–539. https://doi.org/10.1146/annurev-phyto-102313-045914
Cao ZJ, Jing JX, Wang MN, Shang HS, Li ZQ (2003) Relation analysis of stripe rust resistance gene in wheat important cultivar Suwon11, Suwon92 and hybrid 46. Acta Bot Boreal-Occident Sin 23(1):64–68. https://doi.org/10.3321/j.issn:1000-4025.2003.01.013
Chen SB, Songkumarn P, Venu RC, Gowda M, Bellizzi M, Hu JN, Liu WD, Ebbole D, Meyers B, Mitchell T, Wang GL (2013) Identification and characterization of in planta-expressed secreted effector proteins from Magnaporthe oryzae that induce cell death in rice. Mol Plant-Microbe Interact 26(2):191–202. https://doi.org/10.1094/MPMI-05-12-0117-R
Chen WQ, Wellings C, Chen XM, Kang ZS, Liu TG (2014) Wheat stripe (yellow) rust caused by Puccinia striiformis f. sp. tritici. Mol Plant Pathol 15(5):433–446. https://doi.org/10.1111/mpp.12116
Fisher MC, Henk DA, Briggs CJ, Brownstein JS, Madoff LC, Mccraw SL, Gurr SJ (2012) Emerging fungal threats to animal, plant and ecosystem health. Nature 484(7393):186–194. https://doi.org/10.1038/nature10947
Giraldo MC, Dagdas YF, Gupta YK, Mentlak TA, Yi M, Martinez-Rocha AL, Saitoh H, Terauchi R, Talbot NJ, Valent B (2013) Two distinct secretion systems facilitate tissue invasion by the rice blast fungus Magnaporthe oryzae. Nat Commun 4(1):1996. https://doi.org/10.1038/ncomms2996
Gu B, Kale SD, Wang QH, Wang DH, Pan QN, Cao H, Meng YL, Kang ZS, Tyler BM, Shan WX (2011) Rust secreted protein Ps87 is conserved in diverse fungal pathogens and contains a RXLR-like motif sufficient for translocation into plant cells. PLoS One 6(11):e27217. https://doi.org/10.1371/journal.pone.0027217
Guo XR, Niu JF, Cao XY (2018) Heterologous expression of salvia miltiorrhiza microRNA408 enhances tolerance to salt stress in Nicotiana benthamiana. Int J Mol Sci 19(12):3985. https://doi.org/10.3390/ijms19123985
Hawku MD, Goher F, Islam MA, Guo J, He FX, Bai XX, Yuan P, Kang ZS, Guo J (2021) TaAP2-15, an AP2/ERF transcription factor, is positively involved in wheat resistance to Puccinia striiformis f. sp. tritici. Int J Mol Sci 22(4):2080. https://doi.org/10.3390/ijms22042080
Hood ME, Shew HD (1996) Applications of KOH-aniline blue fluorescence in the study of plant-fungal interactions. Phytopathology 86(7):704–708. https://doi.org/10.1094/Phyto-86-704
Jacobs KA, Collins-Racie LA, Colbert M, Duckett MK, Golden-Fleet M, Kelleher K, Kriz R, LaVallie ER, Merberg D, Spaulding V, Stover J, Williamson MJ, McCoy JM (1997) A genetic selection for isolating cDNAs encoding secreted proteins. Gene 198(1–2):289–296. https://doi.org/10.1016/s0378-1119(97)00330-2
Jiang C, Cao SL, Wang ZY, Xu HJ, Liang J, Liu HQ, Wang GH, Ding MY, Wang QH, Gong C, Feng CJ, Hao CF, Xu JR (2019) An expanded subfamily of G-protein-coupled receptor genes in fusarium graminearum required for wheat infection. Nat Microbiol 4(9):1582–1591. https://doi.org/10.1038/s41564-019-0468-8
Kemen E, Kemen A, Ehlers A, Voegele R, Mendgen K (2013) A novel structural effector from rust fungi is capable of fibril formation. Plant J 75(5):767–780. https://doi.org/10.1111/tpj.12237
Kemen E, Kemen AC, Rafiqi M, Hempel U, Mendgen K, Hahn M, Voegele RT (2005) Identification of a protein from rust fungi transferred from haustoria into infected plant cells. Mol Plant-Microbe Interact 18(11):1130–1139. https://doi.org/10.1094/MPMI-18-1130
Kitagaki H, Wu H, Shimoi H, Ito K (2002) Two homologous genes, DCW1 (YKL046c) and DFG5, are essential for cell growth and encode glycosylphosphatidylinositol (GPI)-anchored membrane proteins required for cell wall biogenesis in Saccharomyces cerevisiae. Mol Microbiol 46(4):1011–1022. https://doi.org/10.1046/j.1365-2958.2002.03244.x
Kou YJ, Naqvi NI (2016) Surface sensing and signaling networks in plant pathogenic fungi. Semin Cell Dev Biol 57:84–92. https://doi.org/10.1016/j.semcdb.2016.04.019
Kou YJ, Tan YH, Ramanujam R, Naqvi NI (2017) Structure-function analyses of the Pth11 receptor reveal an important role for CFEM motif and redox regulation in rice blast. New Phytol 214(1):330–342. https://doi.org/10.1111/nph.14347
Kulkarni RD, Dean RA (2004) Identification of proteins that interact with two regulators of appressorium development, adenylate cyclase and cAMP-dependent protein kinase a, in the rice blast fungus Magnaporthe grisea. Mol Gen Genomics 270(6):497–508. https://doi.org/10.1007/s00438-003-0935-y
Kulkarni RD, Kelkar HS, Dean RA (2003) An eight-cysteine-containing CFEM domain unique to a group of fungal membrane proteins. Trends Biochem Sci 28(3):118–121. https://doi.org/10.1016/S0968-0004(03)00025-2
Lamarre C, Deslauriers N, Bourbonnais Y (2000) Expression cloning of the Candida albicans CSA1 gene encoding a mycelial surface antigen by sorting of Saccharomyces cerevisiae transformants with monoclonal antibody-coated magnetic beads. Mol Microbiol 35(2):444–453. https://doi.org/10.1046/j.1365-2958.2000.01715.x
Liu B, Liu TG, Zhang ZY, Jia QZ, Wang BT, Gao L, Peng YL, Jin SL, Chen WQ (2017) Discovery and pathogenicity of CYR34, a new race of Puccinia striiformis f. sp. tritici in China. Acta Phytopathologica Sinica 47(5):681–687. https://doi.org/10.13926/j.cnki.apps.000071
Liu CH, Pedersen C, Schultz-Larsen T, Aguilar GB, Madriz-Ordenana K, Hovmøller MS, Thordal-Christensen H (2016) The stripe rust fungal effector PEC6 suppresses pattern-triggered immunity in a host species-independent manner and interacts with adenosine kinases. New Phytol. https://doi.org/10.1111/nph.14034
Liu P, Guo J, Zhang RM, Zhao JX, Liu C, Qi T, Duan YH, Kang ZS, Guo J (2019) TaCIPK10 interacts with and phosphorylates TaNH2 to activate wheat defense responses to stripe rust. Plant Biotechnol J 17(5):956–968. https://doi.org/10.1111/pbi.13031
Nie J, Yin Z, Li Z, Wu Y, Huang LL (2019) A small cysteine-rich protein from two kingdoms of microbes is recognized as a novel pathogen-associated molecular pattern. New Phytol 222(2):995–1011. https://doi.org/10.1111/nph.15631
Oh SK, Young C, Lee M, Oliva R, Bozkurt T, Cano L, Win J, Bos JIB, Liu HY, Damme MV, Morgan W, Choi D, Vossen EAGVD, Vleeshouwers VGAA, Kamoun S (2009) In planta expression screens of phytophthora infestans RXLR effectors reveal diverse phenotypes, including activation of the Solanum bulbocastanum disease resistance protein Rpi-blb2. Plant Cell 21(9):2928–2947. https://doi.org/10.1105/tpc.109.068247
Peng T, Orsborn KI, Orbach MJ, Galgiani JN (1999) Proline-rich vaccine candidate antigen of Coccidioides immitis: conservation among isolates and differential expression with spherule maturation. J Infect Dis 179(2):518–521. https://doi.org/10.1086/314604
Petre B, Joly DL, Duplessis S (2014) Effector proteins of rust fungi. Front Plant Sci 5:416. https://doi.org/10.3389/fpls.2014.00416
Presti LL, Lanver D, Schweizer G, Tanaka S, Liang L, Tollot M et al (2015) Fungal effectors and plant susceptibility. Annu Rev Plant Biol 66(1):513–545. https://doi.org/10.1146/annurev-arplant-043014-114623
Pretsch K, Kemen A, Kemen E, Geiger M, Mendgen K, Voegele R (2013) The rust transferred proteins-a new family of effector proteins exhibiting protease inhibitor function. Mol Plant Pathol 14(1):96–107. https://doi.org/10.1111/j.1364-3703.2012.00832.x
Qi T, Guo J, Liu P, He FX, Wan CP, Islam MA, Tyler BM, Kang ZS, Guo J (2019a) Stripe rust effector PstGSRE1 disrupts nuclear localization of ROS-promoting transcription factor TaLOL2 to defeat ROS-induced defense in wheat. Mol Plant 12(12):1624–1638. https://doi.org/10.1016/j.molp.2019.09.010
Qi T, Guo J, Peng H, Liu P, Kang ZS, Guo J (2019b) Host-induced gene silencing: a powerful strategy to control diseases of wheat and barley. Int J Mol Sci 20(1):206. https://doi.org/10.3390/ijms20010206
Qi T, Zhu XG, Tan CL, Liu P, Guo J, Kang ZS, Guo J (2018) Host-induced gene silencing of an important pathogenicity factor PsCPK1 in Puccinia striiformis f. sp. tritici enhances resistance of wheat to stripe rust. Plant Biotechnol J 16(3):797–807. https://doi.org/10.1111/pbi.12829
Rafiqi M, Ellis JG, Ludowici VA, Hardham AR, Dodds PN (2012) Challenges and progress towards understanding the role of effectors in plant-fungal interactions. Curr Opin Plant Biol 15(4):477–482. https://doi.org/10.1016/j.pbi.2012.05.003
Sabnam N, Barma SR (2017) WISH, a novel CFEM GPCR is indispensable for surface sensing, asexual and pathogenic differentiation in rice blast fungus. Fungal Genet Biol 105:37–51. https://doi.org/10.1016/j.fgb.2017.05.006
Salomon D, Kinch LN, Trudgian DC, Guo XF, Klimko JA, Grishin NV, Mirzaei H, Orth K (2014) Marker for type VI secretion system effectors. Proc Natl Acad Sci U S A 111(25):9271–9276. https://doi.org/10.1073/pnas.1406110111
Schwessinger B (2017) Fundamental wheat stripe rust research in the 21st century. New Phytol 213(4):1625–1631. https://doi.org/10.1111/nph.14159
Sohn KH, Lei R, Nemri A, Jones JD (2007) The downy mildew effector proteins ATR1 and ATR13 promote disease susceptibility in Arabidopsis thaliana. Plant Cell 19(12):4077–4090. https://doi.org/10.1105/tpc.107.054262
Staples RC (2000) Research on the rust fungi during the twentieth century. Annu Rev Phytopathol 38(1):49–69. https://doi.org/10.1146/annurev.phyto.38.1.49
Tariqjaveed M, Mateen A, Wang SZ, Qiu SS, Zheng XH, Zhang J, Bhadauria V, Sun WX (2021) Versatile effectors of phytopathogenic fungi target host immunity. J Integr Plant Biol 63(11):1856–1873. https://doi.org/10.1111/jipb.13162
Thomas WJ, Thireault CA, Kimbrel JA, Chang JH (2009) Recombineering and stable integration of the pseudomonas syringae pv. Syringae 61 hrp/hrcclusterin to the genomeofthesoil bacterium Pseudomonas fluorescens Pfo-1. Plant J 60(5):919–928. https://doi.org/10.1111/j.1365-313X.2009.03998.x
Vaknin Y, Shadkchan Y, Levdansky E, Morozov M, Romano J, Osherov N (2014) The three aspergillus fumigatus CFEM-domain GPI-anchored proteins (CfmA-C) affect cell-wall stability but do not play a role in fungal virulence. Fungal Genet Biol 63:55–64. https://doi.org/10.1016/j.fgb.2013.12.005
Voegele RT, Mendgen KW (2011) Nutrient uptake in rust fungi: how sweet is parasitic life? Euphytica 179(1):41–55. https://doi.org/10.1007/s10681-011-0358-5
Wang CF, Huang LL, Buchenauer H, Han QM, Zhang HC, Kang ZS (2007) Histochemical studies on the accumulation of reactive oxygen species (O2− and H2O2) in the incompatible and compatible interaction of wheat-Puccinia striiformis f. sp tritici. Physiol Mol Plant Pathol 71(4–6):230–239. https://doi.org/10.1016/j.pmpp.2008.02.006
Wang XD, Wang XJ, Feng H, Tang CL, Bai PF, Wei GR, Huang L. L, Kang ZS (2012) TaMCA4, a novel wheat metacaspase gene functions in programmed cell death induced by the fungal pathogen Puccinia striiformis f. sp. tritici. Mol Plant-Microbe Interact 25(6):755–764. https://doi.org/10.1094/MPMI-11-11-0283-R
Weis WI, Kobilka BK (2018) The molecular basis of G protein-coupled receptor activation. Annu Rev Biochem 87(1):897–919. https://doi.org/10.1146/annurev-biochem-060614-033910
Wellings CR (2011) Global status of stripe rust: a review of historical and current threats. Euphytica 179(1):129–141. https://doi.org/10.1007/s10681-011-0360-y
Xu Q, Tang CL, Wang XD, Sun ST, Zhao JR, Kang ZS, Wang XJ (2019) An effector protein of the wheat stripe rust fungus targets chloroplasts and suppresses chloroplast function. Nat Commun 10(1):5571. https://doi.org/10.1038/s41467-019-13487-6
Yang Q, Huai BY, Lu YX, Cai KY, Guo J, Zhu XG, Kang ZS, Guo J (2020) A stripe rust effector Pst18363 targets and stabilises TaNUDX23 that promotes stripe rust disease. New Phytol 225(2):880–895. https://doi.org/10.1111/nph.16199
Yin C, Hulbert S (2015) Host induced gene silencing (HIGS), a promising strategy for developing disease resistant crops. Gene Technol 4(03):130. https://doi.org/10.4172/2329-6682.1000130
Yin CT, Hulbert S (2011) Prospects for functional analysis of effectors from cereal rust fungi. Euphytica 179(1):57–67. https://doi.org/10.1007/s10681-010-0285-x
Yin WX, Wang YF, Chen T, Lin Y, Luo CX (2018) Functional evaluation of the signal peptides of secreted proteins. Bio-Protocol 8(9):2839. https://doi.org/10.21769/BioProtoc.2839
Zhang ZN, Wu QY, Zhang GZ, Zhu YY, Murphy RW, Liu Z, Zou CG (2015) Systematic analyses reveal uniqueness and origin of the CFEM domain in fungi. Sci Rep 5(1):13032. https://doi.org/10.1038/srep13032
Zhao MX, Wang JF, Ji S, Chen ZJ, Xu JH, Tang CL, Chen ST, Kang ZS, Wang XJ (2018) Candidate effector Pst_8713 impairs the plant immunity and contributes to virulence of Puccinia striiformis f. sp. tritici. Front Plant Sci 9:1294. https://doi.org/10.3389/fpls.2018.01294
Zheng WM, Huang LL, Huang JQ, Wang XJ, Chen XM, Zhao J, Guo J, Zhuang H, Qiu CZ, Liu J, Liu HQ, Huang XL, Pei GL, Zhan GM, Tang CL, Cheng YL, Liu MJ, Zhang JS, Zhao ZT, Zhang SJ, Han QM, Han DJ, Zhang HC, Zhao J, Gao XN, Wang JF, Ni PX, Dong W, Yang LF, Yang HM, Xu JR, Zhang GY, Kang ZS (2013) High genome heterozygosity and endemic genetic recombination in the wheat stripe rust fungus. Nat Commun 4(1):2673. https://doi.org/10.1038/ncomms3673
Zhu WJ, Wei W, Wu YY, Zhou Y, Peng F, Zhang SP, Chen P, Xu XW (2017) BcCFEM1, a CFEM domain-containing protein with putative GPI-anchored site, is involved in pathogenicity, conidial production, and stress tolerance in Botrytis cinerea. Front Microbiol 8:1807. https://doi.org/10.3389/fmicb.2017.01807
Acknowledgments
We thank Professor Larry Dunkle from Purdue University, USA for critical reading of the manuscript.
Funding
This study was financially supported by the National Key Research and Development Program of China (2021YFD1401003), the National Natural Science Foundation of China (32172381 and 31972224), Key Research and Development Program of Shaanxi (2021ZDLNY01–01), Natural Science Basic Research Program of Shaanxi (2020JZ-13) and the 111 Project from the Ministry of Education of China (B07049).
Author information
Authors and Affiliations
Contributions
XB conducted the experiments, analyzed the data, and wrote the manuscript. HP, GF, MI, SX, JG (Jia Guo) assisted in some experiments. ZK revised the manuscript. JG (Jun Guo) conceived and directed the project and revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not relevant.
Consent for publication
All authors agree to publish.
Competing interests
Author ZK is a member of the Editorial Board, but was not involved in the journal’s review of, or decisions related to, this manuscript.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Supplementary Fig. 1. PstCFEM1 in Pst suppresses plant cell death triggered by BAX in N. benthamiana.
Additional file 2:
Supplementary Fig. 2. SignalP 5.0 predicts that PstCFEM1 has a signal peptide.
Additional file 3:
Supplementary Fig. 3. TMHMM predicts that PstCFEM1 lack a transmembrane domain.
Additional file 4:
Supplementary Fig. 4. GPI-anchor Predictor predicts that PstCFEM1 has a GPI-anchor site.
Additional file 5:
Supplementary Fig. 5. EffectorP 3.0 predicts that PstCFEM1 is an apoplastic effector.
Additional file 6:
Supplementary Fig. 6. A Overexpression of PstCFEM1 suppressed Pst322-triggered callose deposition in N. benthamiana. B RT-PCR and (C) western blot were performed to confirm the expression of Pst18363, PstCFEM1, GFP and Pst322 in each infiltration site with specific primers. Nbactin was used as a reference gene.
Additional file 7:
Supplementary Fig. 7. Alignment of the PstCFEM1 coding regions in CYR32, CYR33 and CYR34 races.
Additional file 8:
Supplementary Table 1. Primers used in this study.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bai, X., Peng, H., Goher, F. et al. A candidate effector protein PstCFEM1 contributes to virulence of stripe rust fungus and impairs wheat immunity. Stress Biology 2, 21 (2022). https://doi.org/10.1007/s44154-022-00042-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s44154-022-00042-5