
Abstract
Although the newly developed second-generation anti-androgen drug enzalutamide can repress prostate cancer progression significantly, it only extends the survival of prostate cancer patients by 4–6 months mainly due to the occurrence of enzalutamide resistance. Most of the previous studies on AR antagonist resistance have been focused on AR signaling. Therefore, the non-AR pathways on enzalutamide resistance remain largely unknown. By using C4-2, CWR22Rv1 and LNCaP cell lines, as well as mice bearing CWR22Rv1 xenografts treated with either enzalutamide or metformin alone or in combination, we demonstrated that metformin is capable of reversing enzalutamide resistance and restores sensitivity of CWR22Rv1 xenografts to enzalutamide. We showed that metformin alleviated resistance to enzalutamide by inhibiting EMT. Furthermore, based on the effect of metformin on the activation of STAT3 and expression of TGF-β1, we propose that metformin exerts its effects by targeting the TGF-β1/STAT3 axis. These findings suggest that combination of metformin with enzalutamide could be a more efficacious therapeutic strategy for the treatment of castration-resistant prostate cancer.
Similar content being viewed by others

Main
Prostate cancer is the most commonly diagnosed cancer in men in the western world which causes over 80 deaths per day in the United States of American.1 Androgen-deprivation therapy (ADT) has been used as a standard treatment for advanced prostate cancer for more than six decades.2 Although the recently developed ADT drugs abiraterone and enzalutamide slow down tumor growth effectively,3, 4, 5 recurrence with metastases after ADT are still the major concern in the treatment of castration-resistant prostate cancer (CRPC).6 Lin et al.7 found that bicalutamide and enzalutamide can even induce macrophage migration and tumor cell invasion in orthotopic tumor models of CRPC. Mechanistically, enzalutamide exerts its anti-prostate cancer effects by interrupting the interaction between AR and DHT, blocking AR nuclear translocation, and preventing recruitment of AR to androgen responsive elements.8 Nevertheless, enzalutamide is only capable of extending survival of CRPC patients by 4–6 months partially due to the development of enzalutamide resistance9 but the underlying mechanisms have not been well defined.
Epithelial mesenchymal transition (EMT) was initially identified as a developmental process from an epithelial phenotype to an invasive mesenchymal phenotype. Many transcriptional factors including Twist, Snail, Slug and Zeb1/2 are involved in this process.10 EMT also plays essential roles in cancer cell invasion and metastasis.10, 11, 12, 13, 14 In prostate cancer, elevated levels of mesenchymal biomarkers and reduced epithelial differentiation markers are highly correlated with invasion, metastasis and resistance to ADT.15, 16, 17, 18 It has also been proposed that ADT itself may exert a causal effect in EMT19 and subsequently lead to resistance to enzalutamide treatment.20, 21
Metformin possesses anti-tumor effects to many cancers, including melanoma,22 colon cancer,23 ovarian cancer,24 bladder cancer,25 prostate cancer.26 Different mechanisms have been proposed including inhibiting proliferation, enhancing apoptosis, repressing EMT, targeting cancer stem cells and inhibiting autophagy. Results from a multicenter phase 2 clinical trial showed that metformin could yield objective PSA responses and induce disease stabilization for chemotherapy-naive CRPC.27 We found that combinatorial treatment of metformin and bicalutamide can additively repress the growth of prostate cancer.26 In this research, we demonstrated that metformin is capable of inhibiting enzalutamide-induced EMT in prostate cancer cells via repressing TGF-β1/STAT3 axis.
Results
Metformin enhances enzalutamide’s inhibitory effect on prostate cancer cell growth
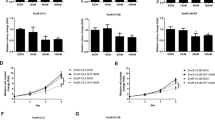
To investigate whether metformin can enhance the anti-tumor effect of enzalutamide on prostate cancer, three prostate cancer cells, C4-2, CWR22Rv1 and LNCaP were treated with either enzalutamide (20 μM) or metformin (5 mM) alone or in combination of both. As shown in Figures 1a–c, comparing to the non-treatment control, the growth rates of all three cancer cell lines were inhibited when they were treated with either agent alone. However, the inhibitive effect is more dramatic when they were combined. In addition, we tested the additive/synergistic effects between enzalutamide and metformin using colony formation assays. Again, colony formation was significantly inhibited for both C4-2 and LNCaP cells when they were treated with either reagent alone, and no colony was detectable when they were treated with the combination of both reagents (Figure 1d). It is well established that CWR22Rv1 cells are resistant to enzalutamide treatment mainly due to the increased expression of the AR-V7, an AR variant constitutively active even in the absence of androgens. To test whether metformin can reverse the resistance of CW22Rv1 cells to enzalutamide, mice bearing xenografts derived from CWR22Rv1 cells were treated with either vehicle, enzalutamide and metformin alone or combination of enzalutamide and metformin for 3 weeks as described in methods. As shown in Figure 1e, the sizes and weights of the tumors were comparable between the enzalutamide-treated and those in the vehicle group and the mouse body weights were comparable in all the four groups. However, metformin alone decreased the sizes and weights of the tumors derived from the CW22Rv1 cells. Surprisingly, the combination of enzalutamide and metformin showed more dramatic inhibitive effect than metformin alone although enzalutamide itself had no effect on the tumors formed from these cells. An alternative explanation is that metformin may be able to reverse the resistance of CWR22Rv1 xenografts to enzalutamide.

Combinatorial effects of enzalutamide and metformin on prostate cancer growth. (a–c) C4-2, CWR22Rv1 and LNCaP cells were seeded in 96-well plates with 0.5 × 105 cells per well in growth media with or without enzalutamide (20 μM) and metformin (5 mM) and cultured for 48 h. Cell viabilities were estimated by CCK-8. (d) Those three cell lines were seeded in six-well plates with 1000 cells per well and treated as indicated for 14 days. Cells were fixed with methanol, stained with crystal violet, and the numbers of colonies were counted. (e) Mice bearing CWR22Rv1 xenografts were treated with vehicle control, enzalutamide, metformin or their combination for 3 weeks; the tumors were collected and weighed. The data represent means±S.D. *P<0.05
Effect of metformin on enzalutamide-mediated cancer cell invasion and migration
Multiple lines of evidence suggest that enzalutamide can promote cell invasion28 and metastasis.29 To determine whether metformin can antagonize this effect, we measured the invasion and migration of different prostate cancer cells using transwell assays with or without a Matrigel-coated membrane. Enzalutamide treatment alone indeed enhanced both invasion and migration in all three cell lines tested (Figures 2a and b). Similar to that reported previously30 that metformin is capable of inhibiting invasion and migration of PC-3 and CWR22Rv1 cells, we found that metformin inhibited the invasion and migration of both LNCaP and C4-2 cells (Figures 2a and b). More importantly, metformin is capable of inhibiting enzalutamide-induced invasion and migration of all three prostate cancer cell lines (Figures 2a and b).

Metformin inhibits enzalutamide-induced cell invasion and migration. (a and b) C4-2, CWR22Rv1 and LNCaP cells were seeded in 24-well transwell chambers with or without Matrigel in growth media with or without enzalutamide (20 μM) and metformin (5 mM) and cultured for 48 h. Cell invasion (a) and migration (b) were estimated. Quantifications were shown on right. The data represent means±S.D. *P<0.05
Effect of metformin on enzalutamide-induced EMT
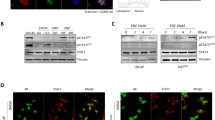
It has been reported that enzalutamide can promote prostate cancer metastasis via the TGF-β1/smad3/MMP9 axis.28 Since this axis plays important role in EMT, we decided to test whether enzalutamide can induce EMT and whether metformin can counteract enzalutamide-induced EMT. Three prostate cancer cells C4-2, CWR22Rv1, and LNCaP were treated with enzalutamide or metformin alone, or combination of both for 48 h followed by western blot assays to estimate the levels of factors involved in EMT. Figures 3a–c showed that enzalutamide treatment alone upregulated the levels of the mesenchymal biomarkers (N-cadherin, Vimentin and TWIST) and downregulated the epithelial markers (E-cadherin), suggesting enzalutamide is capable of promoting EMT. On the other hand, metformin is capable of counteracting enzalutamide-mediated up- and downregulating of the factors involved in EMT (Figures 3a–c). To quantify the expression of EMT-associated proteins, we used ‘Image J’ to analyze bands in western blots, the results of statistical analysis were showed in Supplementary Figure S1. Furthermore, these effects have been further substantiated by the IHC results in the CWR22Rv1-derived xenografts (Figure 3d).

Metformin reverses enzalutamide-induced EMT. (a–c) C4-2, CWR22Rv1 and LNCaP cells was seeded in six-well plates and exposed to enzalutamide (20 μM) or/and metformin (5 mM) for 48 h. Cell lysates were assayed by western blot with antibodies against E-cadherin, N-cadherin, Vimentin and Twist. (d) Tumors collected from 22RV1 cell xenograft model of nude mice treated as indicated were conducted IHC staining with antibodies against E-cadherin, N-cadherin, vimentin and twist. Magnifications: × 400
Metformin represses enzalutamide-induced STAT3 activation and TGF-β1 expression
Since TGF-β1 plays an important role in the enzalutamide-induced invasion and metastasis28 and metformin is capable of inhibiting TGF-β1 by activating AMPK in both breast cancer patients and mouse model,31 we decided to determine if metformin counteracts enzalutamide-induced EMT in prostate cancer by downregulating TGF-β1. Western blot assays showed that enzalutamide upregulated the levels TGF-β1 in all three tested prostate cancer cell lines (Figures 4a–c) and metformin is able to counteract enzalutamide upregulated TGF-β1. These findings are consistent with the levels of TGF-β1 in culture media estimated by ELISA assays (Figures 4d–f). Finally, results from IHC staining of the xenografts derived from CWR22v1 cells (Figure 5a) further substantiated above-mentioned findings. Since STAT3 not only regulates a broad range of genes32 involved in EMT in different cancers33 but activated STAT3 also has been shown to confer enzalutamide resistance,7, 34 we then examined the effect of metformin on STAT3 activation. First, enzalutamide is capable of activating STAT3 in all three prostate cancer cell lines evidenced by the elevated levels of phosphorylated STAT3 (pSTAT3) without affecting the total levels of STAT3 (Figures 4a–c). More importantly, metformin is able to counteract enzalutamide-mediated STAT3 activation (Figures 4a–c). These findings were further substantiated by IHC staining of the CW22RV1 xenografts (Figure 5b). To further substantiate the inhibitory effect of metformin on enzalutamide-induced TGF-β1/STAT3 axis-regulated EMT, we used different concentrations of metformin (1, 5, 10 and 20 mM) combined with enzalutamide (20 μM) to treat the three cell lines. We found that metformin was indeed capable of inhibiting enzalutamide-induced EMT via TGF-β1/STAT3 axis in a dose dependent manner (Supplementary Figure S2). Altogether, these results indicate that metformin can reverse enzalutamide resistance by counteracting enzalutamide-mediated STAT3 activation, subsequent upregulation of TGF-β1 and ultimate prostate cancer invasion and metastasis (Figure 6), suggesting that combination of metformin and enzalutamide could be more efficient in treating castration-resistant prostate cancer.

Metformin represses enzalutamide-induced TGF-β1 expression and STAT3 activation. (a–c) C4-2, CWR22Rv1 and LNCaP cells was seeded in 6-well plates and treated as indicated for 48 h. The levels of TGF-β1, p-STAT3 and STAT3 were estimated by western blot assays. (d–f) and the culture media were collect for ELISA assays of TGF-β1. The data represent means±S.D. *P<0.05

Tumor tissues from 22RV1 cell xenograft model of nude mice were stained for TGF-β1 (a) and p-STAT3 (b). Magnifications: × 400; higher magnification images, bottom panels

Schematic model of the hypothesized mechanism by which metformin reverses enzalutamide resistance
Discussion
As a second-generation AR antagonist, enzalutamide has been widely used in treatment of advanced prostate cancer although the development of resistance is inevitable.35 In this study, we showed that EMT plays an important role in the development of enzalutamide resistance and metformin is capable of reversing enzalutamide-induced EMT. Mechanistically, we demonstrated that metformin exerts its effect by inhibiting TGF-β1 expression and STAT3 activation. These findings were highly consistent in our prostate cancer cell models in vitro and CWR22Rv1 xenograft mice models in vivo, suggesting that combination of metformin and enzalutamide would be more efficient in treating patients with advanced prostate cancer, especially the ones with enzalutamide-resistant cancer.
It is well established that alteration of TGF-β1 signaling and aberrant activation of STAT3 are closely correlated with the progression of various types of tumors, and an active STAT3 signaling pathway is essential for TGF-β-induced EMT.36, 37, 38 By directly binding to the promoter region of Snail, STAT3 serves as a regulatory transcriptional factor in regulation of the expression of Snail and subsequent induction of EMT.39 We have reported that activation of STAT3 is associated with EMT30 and CSCs repopulations.40 In this study, we found that metformin is capable of inhibiting enzalutamide-induced TGF-β1 expression and STAT3 activation. Based on that (1) elevated levels of TGF-β1 in enzalutamide-treated prostate cancers occurred concurrently with activation of STAT3 and enhanced EMT; and (2) metformin is capable of downregulating TGF-β1, inhibiting STAT3 activation and more importantly the enzalutamide-induced EMT, we speculate that one of the mechanisms in metformin-mediated reversal of enzalutamide resistance is by targeting EMT via TGF-β1/STAT3 axis. It has also been reported that metformin is capable of inhibiting the interaction between TGF-β1 and its receptor as well as type II TGF-β1 receptor dimerization.41 However, whether metformin’s effect on the interaction between TGF-β1 and its receptor is unclear and worth of further examination.
In addition, both ligand binding domain (LBD) mutation of AR and formation of constitutively active AR variants, especially AR-V742, 43 play crucial roles in the development of enzalutamide resistance. Chengfei et al.44 showed that chronical treatment of prostate cancer patient with enzalutamide upregulated the expression of AR variants. Antonarakis et al.45 found that presence of AR-V7 in CTCs prior treatment conferred the cancer cells’ resistance to both enzalutamide and abiraterone. In addition, one of our previous studies26 has demonstrated that metformin is capable of downregulating ARV7. Therefore, we do not exclude the possibility that metformin counteracts enzalutamide-induced resistance by targeting either the mutant full-length AR or AR variants. Since autophagy has been considered as a novel mechanism for CRPC cells to evade enzalutamide treatment46 and metformin can inhibit autophagy in prostate cancer cells via AMPK/mTOR signaling pathway, it is worthy to explore the effect of metformin along this axis.
In summary, we demonstrated that metformin is capable of improving the efficacy of enzalutamide treatment by inhibiting enzalutamide-induced EMT. This finding provides the rationale for using combination of enzalutamide and metformin as a more efficient therapeutic strategy in the treatment of advanced prostate cancer.
Materials and methods
Reagents and cell culture
Human prostate cancer cell lines (LNCaP, C4-2 and CWR22Rv1) were obtained from Cell Bank of Shanghai Institutes for biological Sciences (Chinese Academy of Sciences), and cultured in RPMI 1640 supplemented with 10% FBS (Gibco) at 37 °C with 5% CO2. Enzalutamide was kindly provided by Dr. Lei and metformin was purchased from Sigma.
Cell viability assay
C4-2, CWR22Rv1 and LNCaP cells were seeded into 96-well plates and after grown to 60–75% confluence, were treated with vehicle control, enzalutamide, metformin, and combination in RPMI 1640 with 10% FBS for 48 h. The viabilities of the cells were evaluated using the Cell Counting Kit-8 (CCK8) assay. Briefly, at termination of exposure, cells were aspirated and rinsed with PBS then treated with 10 μl/well CCK-8 for 2 h at 37 °C. Absorbance was measured at 450nm spectrophotometrically (Bio-Rad, Hercules, CA, USA).
Clonogenic assays
C4-2, CWR22Rv1 or LNCaP cells were seeded in 6-well plates with 1000 cells per well and incubated at 37 °C in culture media containing either enzalutamide (20 μM) and metformin (5 mM) alone or in combination for 14 days with the medium changed every 7 days. At the end of the experiment, cells were fixed with methanol, stained with crystal violet and the numbers of colonies were counted.
Transwell assays
To assess cell migration and invasion in vitro, we used 24-well transwell chambers with or without Matrigel. LNCaP, C4-2 and CWR22Rv1 cells were trypsinized and seeded into the top chamber at a density of 5 × 104 cells per well in 200 μl Dulbecco’s modified Eagle’s medium containing metformin (5 mM), enzalutamide (20 μM) or combination of metformin (5 mM) and enzalutamide (20 μM). The outer chambers contained 800 μl of medium (10% fetal calf serum). After incubation at 37 °C for 48 h, cells attached to the upper surface of the membrane were carefully removed with cotton swabs, whereas cells that reached the underside of the chamber were fixed with 10% formalin and stained with crystal violet for 3 min at room temperature and counted.
Western blotting
LNCaP, C4-2 and CWR22Rv1 cells were seeded in six-well plates, 2 × 105cells/well, treated with corresponding reagents, including metformin (5 mM), enzalutamide (20 μM) or combination of metformin (5 mM) and enzalutamide (20 μM). Cell lysates were separated on SDS-PAGE followed by western blotting assay as described previously26, 47with the following primary antibodies: E-cadherin (1:1000 Proteintech, Rosemont, IL, USA), N-cadherin (1:500 Proteintech), TWIST (1:500 Santa Cruz Biotechnology, Dallas, TX, USA), Vimentin (1:1000 Proteintech), STAT3 (1:2000 Abcam, Cambridge, MA, USA), p-STAT3 (Tyr705) (1:2000 Cell Signaling, Danvers, MA, USA), TGF-β1 (1:200 Proteintech) and β-actin(1:5000 Cell Signaling). Image J (NIH, USA) was used to quantify the expression of proteins.
TGF-β1 ELISA assay
C4-2, CWR22Rv1 and LNCaP Cells were seeded in six-well plates and treated with enzalutamide (20 μM), metformin (5 mM) or combination for 24 h. The supernatant was collected and TGF-β1 released to the culture media was measured using commercially available enzyme-linked immunosorbent assay (ELISA) kits from Proteintech.
In vivo tumorigenesis assay
CWR22Rv1 cells (four million/50 μl) mixed with matrigel (1:1) were injected subcutaneously into the flanks of 6–7 weeks castrated male nude mice. Three weeks after injection, tumor-bearing mice (tumor volumes were around 50–100 mm3) were randomized into four groups (with 4 mice in each group) and treated with vehicle control (NC group), or enzalutamide (25 mg/kg/day) (ENZ group), or metformin (300 mg/kg/day) (MET group), or combination of enzalutamide (25 mg/kg/day) and metformin (300 mg/kg/day) (ENZ+MET group) for 3 weeks via esophageal gavaging. After killing the mice, the transplanted tumors were weighed and then fixed in 10% formalin and paraffin embedded for immunostaining examination.
Immunohistochemical staining
Transplanted tumor specimens were fixed in 10% formaldehyde solution and embedded in paraffin, and sections were mounted onto glass slides. The sections were then deparaffinized in xylene, re-hydrated through ethanol, and heated for 30 min to enhance the heat-induced antigen retrieval. To block non-specific reactions, slides were blocked in respective serum at 4 °C overnight. Primary antibodies against E-cadherin (1:500 Proteintech), N-cadherin (1:50 Proteintech), TWIST (1:50 Santa Cruz Biotechnology), Vimentin (1:100 Proteintech), p-STAT3 (Tyr705) (1:100 Cell Signaling), TGF-β1 (1:50 Proteintech) were used. Tissue sections were incubated with each antibody overnight at 4 °C, and then incubated with horseradish peroxidase-conjugated anti-rabbit IgG secondary antibodies. Slides were subsequently treated with a streptavidin-peroxidase reagent and incubated in phosphate-buffered salinediaminobenzidine and 1% hydrogen peroxide, followed by counterstaining with Mayer’s haematoxylin.
Statistical analysis
GraphPad Prism 5.0 (San Diego, CA, USA) was used for all statistical analyses. Data were presented as means±S.D. Differences between individual groups were analyzed by one-way analysis of variance followed by the LSD procedure for comparison of means, a P-value of <0.05 was considered statistically significant.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
Siegel RL, Miller KD, Jemal A . Cancer statistics, 2015. CA Cancer J Clin 2015; 65: 5–29.
Huggins C, Hodges CV, . Studies on prostatic cancer: I. The effect of castration, of estrogen and of androgen injection on serum phosphatases in metastatic carcinoma of the prostate. 1941. J Urol 2002; 168: 9–12.
Tran C, Ouk S, Clegg NJ, Chen Y, Watson PA, Arora V et al. Development of a second-generation antiandrogen for treatment of advanced prostate cancer. Science 2009; 324: 787–790.
Ryan CJ, Smith MR, de Bono JS, Molina A, Logothetis CJ, de Souza P et al. Abiraterone in metastatic prostate cancer without previous chemotherapy. N Engl J Med 2013; 368: 138–148.
Beer TM, Tombal B . Enzalutamide in metastatic prostate cancer before chemotherapy. N Engl J Med 2014; 371: 1755–1756.
Miyamoto H, Messing EM, Chang C . Androgen deprivation therapy for prostate cancer: current status and future prospects. Prostate 2004; 61: 332–353.
Lin TH, Izumi K, Lee SO, Lin WJ, Yeh S, Chang C . Anti-androgen receptor ASC-J9 versus anti-androgens MDV3100 (Enzalutamide) or Casodex (Bicalutamide) leads to opposite effects on prostate cancer metastasis via differential modulation of macrophage infiltration and STAT3-CCL2 signaling. Cell Death Dis 2013; 4: e764.
Thadani-Mulero M, Nanus DM, Giannakakou P . Androgen receptor on the move: boarding the microtubule expressway to the nucleus. Cancer Res 2012; 72: 4611–4615.
Bianchini D, Lorente D, Rodriguez-Vida A, Omlin A, Pezaro C, Ferraldeschi R et al. Antitumour activity of enzalutamide (MDV3100) in patients with metastatic castration-resistant prostate cancer (CRPC) pre-treated with docetaxel and abiraterone. Eur J Cancer 2014; 50: 78–84.
Kalluri R, Weinberg RA . The basics of epithelial-mesenchymal transition. J Clin Invest 2009; 119: 1420–1428.
Singh A, Settleman J . EMT, cancer stem cells and drug resistance: an emerging axis of evil in the war on cancer. Oncogene 2010; 29: 4741–4751.
Ouyang G, Wang Z, Fang X, Liu J, Yang CJ . Molecular signaling of the epithelial to mesenchymal transition in generating and maintaining cancer stem cells. Cell Mol Life Sci 2010; 67: 2605–2618.
Thiery JP, Acloque H, Huang RY, Nieto MA . Epithelial-mesenchymal transitions in development and disease. Cell 2009; 139: 871–890.
Yang J, Weinberg RA . Epithelial-mesenchymal transition: at the crossroads of development and tumor metastasis. Dev Cell 2008; 14: 818–829.
Beach S, Tang H, Park S, Dhillon AS, Keller ET, Kolch W et al. Snail is a repressor of RKIP transcription in metastatic prostate cancer cells. Oncogene 2008; 27: 2243–2248.
Odero-Marah VA, Wang R, Chu G, Zayzafoon M, Xu J, Shi C et al. Receptor activator of NF-kappaB Ligand (RANKL) expression is associated with epithelial to mesenchymal transition in human prostate cancer cells. Cell Res 2008; 18: 858–870.
Bitting RL, Schaeffer D, Somarelli JA, Garcia-Blanco MA, Armstrong AJ . The role of epithelial plasticity in prostate cancer dissemination and treatment resistance. Cancer Metastasis Rev 2014; 33: 441–468.
Sun Y, Campisi J, Higano C, Beer TM, Porter P, Coleman I et al. Treatment-induced damage to the tumor microenvironment promotes prostate cancer therapy resistance through WNT16B. Nat Med 2012; 18: 1359–1368.
Sun Y, Wang BE, Leong KG, Yue P, Li L, Jhunjhunwala S et al. Androgen deprivation causes epithelial-mesenchymal transition in the prostate: implications for androgen-deprivation therapy. Cancer Res 2012; 72: 527–536.
Martin SK, Pu H, Penticuff JC, Cao Z, Horbinski C, Kyprianou N . Multinucleation and mesenchymal-to-epithelial transition alleviate resistance to combined cabazitaxel and antiandrogen therapy in advanced prostate cancer. Cancer Res 2016; 76: 912–926.
Ware KE, Somarelli JA, Schaeffer D, Li J, Zhang T, Park S et al. Snail promotes resistance to enzalutamide through regulation of androgen receptor activity in prostate cancer. Oncotarget 2016; 7: 50507–50521.
Tomic T, Botton T, Cerezo M, Robert G, Luciano F, Puissant A et al. Metformin inhibits melanoma development through autophagy and apoptosis mechanisms. Cell Death Dis 2011; 2: e199.
Buzzai M, Jones RG, Amaravadi RK, Lum JJ, DeBerardinis RJ, Zhao F et al. Systemic treatment with the antidiabetic drug metformin selectively impairs p53-deficient tumor cell growth. Cancer Res 2007; 67: 6745–6752.
Patel S, Singh N, Kumar L . Evaluation of effects of metformin in primary ovarian cancer cells. Asian Pac J Cancer prevention: APJCP 2015; 16: 6973–6979.
Nayan M, Bhindi B, Yu JL, Hermanns T, Mohammed A, Hamilton RJ et al. The effect of metformin on cancer-specific survival outcomes in diabetic patients undergoing radical cystectomy for urothelial carcinoma of the bladder. Urol Oncol 2015; 33: 386 e7–13.
Wang Y, Liu G, Tong D, Parmar H, Hasenmayer D, Yuan W et al. Metformin represses androgen-dependent and androgen-independent prostate cancers by targeting androgen receptor. Prostate 2015; 75: 1187–1196.
Rothermundt C, Hayoz S, Templeton AJ, Winterhalder R, Strebel RT, Bartschi D et al. Metformin in chemotherapy-naive castration-resistant prostate cancer: a multicenter phase 2 trial (SAKK 08/09). Eur Urol 2014; 66: 468–474.
Lin TH, Lee SO, Niu Y, Xu D, Liang L, Li L et al. Differential androgen deprivation therapies with anti-androgens casodex/bicalutamide or MDV3100/Enzalutamide versus anti-androgen receptor ASC-J9(R) Lead to promotion versus suppression of prostate cancer metastasis. J Biolo Chem 2013; 288: 19359–19369.
Sha K, Yeh S, Chang C, Nastiuk KL, Krolewski JJ . TNF signaling mediates an enzalutamide-induced metastatic phenotype of prostate cancer and microenvironment cell co-cultures. Oncotarget 2015; 6: 25726–25740.
Tong D, Liu Q, Liu G, Xu J, Lan W, Jiang Y et al. Metformin inhibits castration-induced EMT in prostate cancer by repressing COX2/PGE2/STAT3 axis. Cancer Lett 2016; 389: 23–32.
Li NS, Zou JR, Lin H, Ke R, He XL, Xiao L et al. LKB1/AMPK inhibits TGF-beta1 production and the TGF-beta signaling pathway in breast cancer cells. Tumour Biol 2016; 37: 8249–8258.
Yu H, Lee H, Herrmann A, Buettner R, Jove R . Revisiting STAT3 signalling in cancer: new and unexpected biological functions. Nat Rev Cancer 2014; 14: 736–746.
Marcucci F, Rumio C, Lefoulon F . Anti-Cancer Stem-like Cell Compounds in Clinical Development - An Overview and Critical Appraisal. Front Oncol 2016; 6: 115.
Liu C, Zhu Y, Lou W, Cui Y, Evans CP, Gao AC . Inhibition of constitutively active Stat3 reverses enzalutamide resistance in LNCaP derivative prostate cancer cells. Prostate 2014; 74: 201–209.
Kim W, Ryan CJ . Androgen receptor directed therapies in castration-resistant metastatic prostate cancer. Curr Treat Options Oncol 2012; 13: 189–200.
Liu RY, Zeng Y, Lei Z, Wang L, Yang H, Liu Z et al. JAK/STAT3 signaling is required for TGF-beta-induced epithelial-mesenchymal transition in lung cancer cells. Int J Oncol 2014; 44: 1643–1651.
Cho KH, Jeong KJ, Shin SC, Kang J, Park CG, Lee HY . STAT3 mediates TGF-beta1-induced TWIST1 expression and prostate cancer invasion. Cancer Lett 2013; 336: 167–173.
Zhao S, Venkatasubbarao K, Lazor JW, Sperry J, Jin C, Cao L et al. Inhibition of STAT3 Tyr705 phosphorylation by Smad4 suppresses transforming growth factor beta-mediated invasion and metastasis in pancreatic cancer cells. Cancer Res 2008; 68: 4221–4228.
Balanis N, Wendt MK, Schiemann BJ, Wang Z, Schiemann WP, Carlin CR . Epithelial to mesenchymal transition promotes breast cancer progression via a fibronectin-dependent STAT3 signaling pathway. J Biol Chem 2013; 288: 17954–17967.
Liu Q, Yuan W, Tong D, Liu G, Lan W, Zhang D et al. Metformin represses bladder cancer progression by inhibiting stem cell repopulation via COX2/PGE2/STAT3 axis. Oncotarget 2016; 7: 28235–28246.
Xiao H, Zhang J, Xu Z, Feng Y, Zhang M, Liu J et al. Metformin is a novel suppressor for transforming growth factor (TGF)-beta1. Sci Rep 2016; 6: 28597.
Welti J, Rodrigues DN, Sharp A, Sun S, Lorente D, Riisnaes R et al. Analytical validation and clinical qualification of a new immunohistochemical assay for androgen receptor splice variant-7 protein expression in metastatic castration-resistant prostate cancer. Eur Urol 2016; 70: 599–608.
Karantanos T, Evans CP, Tombal B, Thompson TC, Montironi R, Isaacs WB . Understanding the mechanisms of androgen deprivation resistance in prostate cancer at the molecular level. Eur Urol 2015; 67: 470–479.
Liu C, Lou W, Zhu Y, Nadiminty N, Schwartz CT, Evans CP et al. Niclosamide inhibits androgen receptor variants expression and overcomes enzalutamide resistance in castration-resistant prostate cancer. Clin Cancer Res 2014; 20: 3198–3210.
Antonarakis ES, Lu C, Wang H, Luber B, Nakazawa M, Roeser JC et al. AR-V7 and resistance to enzalutamide and abiraterone in prostate cancer. N Engl J Med 2014; 371: 1028–1038.
Nguyen HG, Yang JC, Kung HJ, Shi XB, Tilki D, Lara PN Jr. et al. Targeting autophagy overcomes Enzalutamide resistance in castration-resistant prostate cancer cells and improves therapeutic response in a xenograft model. Oncogene 2014; 33: 4521–4530.
Tong D, Liu Q, Liu G, Yuan W, Wang L, Guo Y et al. The HIF/PHF8/AR axis promotes prostate cancer progression. Oncogenesis 2016; 5: e283.
Acknowledgements
We thank Dr. Alan Gao’s (University of California, Davis, USA) kindness of providing the cell lines expressing IL6. We also appreciate the help on western blot assays provided by Shawn Kennemuth and Tina Joseph (Philadelphia College of Osteopathic Medicine, PA, USA). This work was supported for Jun Jiang and Dali Tong by the National Natural Science Foundation of China (NSFC) (Grant No: 81172442 and 81402120) and Chongqing Science and Technology Commission (Grant No: 2011GZ0047).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Edited by A Oberst
Supplementary Information accompanies this paper on Cell Death and Disease website
Rights and permissions
Cell Death and Disease is an open-access journal published by Nature Publishing Group. This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

{kind=link}
{kind=link}
Cite this article
Liu, Q., Tong, D., Liu, G. et al. Metformin reverses prostate cancer resistance to enzalutamide by targeting TGF-β1/STAT3 axis-regulated EMT. Cell Death Dis 8, e3007 (2017). https://doi.org/10.1038/cddis.2017.417
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cddis.2017.417
- Springer Nature Limited
This article is cited by
-
Molecular panorama of therapy resistance in prostate cancer: a pre-clinical and bioinformatics analysis for clinical translation
Cancer and Metastasis Reviews (2024)
-
Analysis of differential membrane proteins related to matrix stiffness-mediated metformin resistance in hepatocellular carcinoma cells
Proteome Science (2023)
-
Genetic profiling of hormone-sensitive and castration-resistant prostate cancers and identification of genetic mutations prone to castration-resistant prostate cancer
Prostate Cancer and Prostatic Diseases (2023)
-
Expression profiling of extramedullary acute myeloid leukemia suggests involvement of epithelial–mesenchymal transition pathways
Leukemia (2023)
-
Anti-cancer Efficacy of Metformin: Recent Updates on Breast and Other Cancers
Current Pharmacology Reports (2023)