

Abstract
Climate change threatens the role of European forests as a long-term carbon sink. Assisted migration aims to increase the resilience of forest tree populations to climate change, using species-specific climatic limits and local adaptations through transferring seed provenances. We modelled assisted migration scenarios for seven main European tree species and analysed the effects of species and seed provenance selection, accounting for environmental and genetic variations, on the annual above-ground carbon sink of regrowing juvenile forests. To increase forest resilience, coniferous trees need to be replaced by deciduous species over large parts of their distribution. If local seed provenances are used, this would result in a decrease of the current carbon sink (40 TgC yr−1) by 34–41% by 2061–2080. However, if seed provenances adapted to future climates are used, current sinks could be maintained or even increased to 48–60 TgC yr−1.
Similar content being viewed by others

Main
By removing carbon dioxide from the atmosphere, forests can play a crucial role in limiting the rise in global annual temperatures to 1.5–2 °C (refs. 1,2). European forests cover ~35% of the total land surface and store ~9.8 Tg of carbon, with an average annual sequestration of 155 TgC between 2010 and 20203. However, climate change-induced disturbances pose a serious threat to this carbon pool4,5,6 and other forest ecosystem services. Therefore, adaptation actions are required to maintain ecosystem services and improve the resilience of forests towards further climate warming7,8,9,10. To use the carbon sequestration capacity of forests, tree planting and forest restoration have been suggested as nature-based solutions to mitigate global warming11,12,13,14. The selection of tree species, suitable forest reproductive material and the promotion of mixed forests14,15 are expected to play important roles in forest adaptation and climate change (CC) mitigation16,17.
Owing to the strong impact of CC on tree species distributions18,19,20 and the limited migration capacity of trees21,22, forest restoration and adaptation will need to take the site suitability of species and populations into account. Moreover, restricted gene flow, as well as the limited capacity of trees to adapt genetically to the fast pace of CC, will result in serious maladaptations (but see ref. 23), local species extinctions and a reduction of ecosystem services within the remaining forests23,24,25,26. Therefore, assisted migration (AM) of tree species and seed provenances have been proposed as measures to use species-specific climatic limits and local adaptations to improve the adaptive capacity and resilience of forests while maintaining the provision of ecosystem services27,28,29,30,31. While AM can be grouped into ‘assisted gene flow’ referring to the transfer of seed provenances to new locations within the present distribution range and ‘assisted species migration’ referring to moving seed provenances beyond the present species range, we refer to AM as a more generic term including both types28,32. Despite growing evidence for increasing mismatches between local adaptations and the changing climate for forest trees around the world33,34, no comprehensive multispecies, continent-wide analysis of the needs and effects of AM exists to date, limiting our understanding of how local adaptations affect major ecosystem services such as the forest carbon sink (CS). On the European scale, such a study could motivate AM schemes for forest trees across the ranges of species and support policies on nature-based CC mitigation strategies.
Here we demonstrate the implementation of AM in reforestation of seven major European trees by analysing a dataset of 587 range-wide provenance trials, evaluating 2,964 provenances from their entire distribution (Supplementary Table 1 and Supplementary Figs. 1–3) and combining shifts of species distributions with population transfer models. We quantify the combined effects of adapting the selection of species and seed provenances on annual above-ground CS of European forests. Although the positive effects of species mixtures on productivity are well-documented14, our analysis is limited to single-species forests at a level of a 1 × 1 km grid cell to focus on the direct effects of seed provenance selection.
First, we selected the climatically best-suited species (species with the highest probability of occurrence) at a spatial resolution of a 1 × 1 km grid cell with multimodel ensemble species distribution models (SDMs)35 (Supplementary Table 6) and used the underlying single-model SDMs for uncertainty analysis. Second, for each grid cell with the best-suited species, we identified the best-fitting seed provenances by modelling their annual CS. For a realistic estimation of CS, we restricted our predictions to those grid cells of the land surface, where at least 50% of the area is covered by forests36. Thus, our models address reforestation on previous forest sites (forest regrowth), which allows us to assume constant carbon stocks in forest soils given their high variation and uncertain response to CC and forest development37. To identify the best-fitting seed provenances, we developed universal response functions (URFs) for the seven tree species Abies alba, Larix decidua, Picea abies, Pinus sylvestris, Fagus sylvatica, Quercus robur and Quercus petraea. These models account for environmental and genetic trait variation in above-ground CS38,39 and allow estimates of CS to forest regrowth until stand ages of 40 years because of (1) the mitigation potential of tree planting and reforestation needs to enfold high sequestration capacity as soon as possible to limit further global warming and (2) the limitations of the provenance dataset, which covers predominantly trials with ages up to 40 years. To use the URF for meaningful AM applications, tree species and seed source occurrences across Europe were grouped into species-specific seed provenance clusters (SPC) of climatically and geographically similar groups to correspond to range-wide local adaptation patterns (Supplementary Table 8). To quantify the effects of AM, two seed provenance sourcing scenarios were used: (1) ‘local seeds’ where the climate of the planting location and the historical climate of seed origin were identical and geographically proximate and (2) ‘adapted seeds’, where SPC producing highest CS for the climate of the planting location were selected irrespective of their climatic and geographic proximity to the planting location (Supplementary Fig. 4). These scenarios were tested for contemporary climate (1991–2010) and the period 2061–2080 under the two CC scenarios representative concentration pathways RCP 4.5 and RCP 8.5 (ref. 40). For each CC scenario, five bias-corrected regional climate models (RCMs) were applied (Supplementary Table 2) contributing to a full uncertainty analysis (Supplementary Table 9), where the variation of RCMs, the variation of SDMs and the variation of the provenance selection were used to quantify the uncertainty of future CS. The relative contributions of the sources of uncertainty41 related to the modelling frameworks and the AM scenario were mapped to quantify the geographical distribution of the sources of uncertainties across Europe (Supplementary Table 13 and Supplementary Figs. 16–22).
Large changes in forest composition required under climate change
All SDMs and climate model combinations indicate that CC is projected to cause drastic shifts in the climatic suitability of tree species, calculated as the species with the highest probability of occurrence in each forested grid cell of Europe (Table 1 and Supplementary Table 10). The suitability of conifers such as P. abies, A. alba and L. decidua, which have dominant shares in Central and Northern European forests as a result of their wide cultivation in past centuries42, declines drastically resulting in major shifts in species distributions (Table 1 and Supplementary Table 10), as reported previously16,19. Under future climate, the relative share of broadleaved species such as F. sylvatica and Q. robur is expected to exceed the share of conifers (Table 1 and Supplementary Fig. 8) but all species will experience declines in suitable habitats, especially in south and southeastern Europe35. Although the uncertainty in suitable cultivation areas due to different SDMs, RCMs and their combination varies between species and increases from an average of 4% under contemporary climate to 25% under RCP 8.5 (Table 1, Supplementary Table 10 and Supplementary Figs. 5 and 6), the rankings of species in suitable cultivation areas remain largely constant (Supplementary Fig. 8).
The scenarios for AM (Supplementary Fig. 4) of the seven main tree species focused on optimizing the CS potential of the regenerated forests. On the basis of extensive range-wide provenance trials, our models indicate that the CS for most of the tree species can be optimized if non-local SPC suitable for projected CC scenarios are being planted (Fig. 1 and Supplementary Figs. 9 and 11).

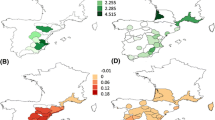
a–f, Geographic locations (a,d) of the SPC and distribution maps of P. abies (b,c) and F. sylvatica (e,f), where the SPC that are predicted to yield the highest annual CS for each 1 × 1 km grid cell are marked in the respective SPC colour. Green colours mark areas where local seed sources are the most productive seed provenances. SPC selections are shown for contemporary climate (1991–2010) and future climate under the RCP 8.5 scenario in the period 2061–2080. Best-suited SPC in the RCP 8.5 scenario was identified by URFs predicted for the mean climate of the five RCMs (Supplementary Table 2). See Supplementary Fig. 11 for other species. For better visualization, the maps present every 1 × 1 km grid cell of the contemporary distribution of the species, while the CSs reported in the study are restricted only to forested grid cells of Europe.
Local seed provenances offer optimal CS in regenerated forests only on restricted parts of their range (Supplementary Table 11). In contemporary climate, local seed provenances provide higher CS compared to adapted seed provenances only in 3–4% of the total suitable area of P. abies and P. sylvestris, 15–18% in L. decidua and A. alba, 8–16% in the two oak species (8–17%) but 50% in case of F. sylvatica (Supplementary Table 11). Under the strong CC scenario RCP 8.5, the suitable area for deployment of local seed provenances decreased to 2–19% of the respective species distribution. The strongest decline in suitable areas for local material was observed in F. sylvatica (5% of the area) and Q. petraea (8%) (Supplementary Table 11). The suitable area for deploying local seeds remained largely constant for L. decidua, P. sylvestris and A. alba and slightly increased from 8% to 15% for Q. robur (Supplementary Table 11). Under contemporary climate, the uncertainty in the share of suitable area for local and adapted seed sources is small ranging from 0.1% to 1.2% of the total area for local material and from 0.6% to 2.1% for adapted material (Supplementary Table 11). For future climate scenarios, uncertainty increased to between 0.5% and 4.0% for local seed sources and 3.4% and 30.9% of the total area for adapted seed sources. The poor performance of many local seed sources regarding CS even under contemporary climate might be explained by the incomplete expansion of many European tree species after the last glaciation21 and by adaptation lags of many tree populations observed particularly within marginal populations23,33,43,44,45,46. Previously, such adaptation lags were mainly found for growth and productivity45 but more recently also for phenological or physiological traits23,44,46.
Following AM, the best seed provenances of A. alba originate from the cold and dry regions of the Carpathians, whereas for L. decidua, the best seed provenances originate from lower elevations of mountain regions of eastern central Europe and the Polish lowlands, where the native range consists of small scattered populations47. For P. abies (Fig. 1) and P. sylvestris (Supplementary Fig. 11), conifers with a large geographic range, a variety of SPC from the Carpathians, central and atlantic Europe are suitable each for a specific region. For the deciduous species F. sylvatica (Fig. 1) and Q. petraea, SPC from the alpine and atlantic regions performed best over a large part of the future distribution, while for Q. robur, SPC from central and partly southeastern Europe outperformed others (Supplementary Fig. 11). Irrespective of variations in their recommended cultivation area due to underlying SDM–RCM–URF combinations, the relative ranking of productive SPC remains largely constant with a few SPC being most suitable across large parts of the future distribution areas (Supplementary Figs. 9 and 11).
Assisted migration can maintain European forest carbon sequestration
Tree species change and seed provenance selections have strong effects on the annual CS of forest regrowth (Table 3). If local seed provenances are selected, the change from predominantly coniferous species to deciduous trees is expected to decrease the overall CS of the reforestations48 from 39.76 ± 1.14 TgC yr−1 to 23.46 ± 0.98 TgC yr−1 under RCP 4.5 and 21.67 ± 2.54 TgC yr−1 under RCP 8.5 scenarios in the period 2061–2080 (Fig. 2 and Table 2).

Total annual above-ground CS (TgC yr−1) for the seven tree species under the two seed source scenarios ‘adapted’ and ‘local’ seed provenances. Local are those seed provenances that are geographically proximate to the planting locations and where the climate of the planting location and the historical climate of seed origin are identical. Adapted seeds are those SPC producing the highest annual CS for the climate of the planting location irrespective of their climatic similarity and geographic proximity to the planting location. CS from local and adapted seed sources in RCP 4.5 and 8.5 scenarios were identified by URFs predicted for the mean climate of five RCMs (Supplementary Table 2). The error bars represent the standard deviation of the annual CS (TgC yr−1) of the sum of the seven species and are calculated from the different combinations of SDMs, URFs and RCMs.
If instead, the most productive SPC according to AM for the respective climate is planted, reforestations could contribute to an increase of annual CS to 66.78 ± 1.72 TgC yr−1 under contemporary climate and 59.61 ± 2.36 TgC yr−1 under the RCP 4.5 scenario during 2061–2080. Moreover, even under the pessimistic CC scenario RCP 8.5, AM would allow increasing the annual CS slightly to 48.18 ± 3.52 TgC yr−1 during 2061–2080 (Fig. 2 and Table 3). Despite variations in CS due to various combinations of SDMs, URFs and RCMs (Supplementary Tables 12 and 13) all models indicate a higher annual CS of adapted seed sources compared to local seed sources and the relative contribution of the seven species on annual CS remained constant (Fig. 2, Table 2 and Supplementary Table 12).
Under contemporary climate, the CS of the modelled forest stands would account for about 26% of the observed annual CS (155 TgC yr−1) of European forests3 if planted with local seed materials and for about 43% if adapted seed sources are used (Table 2). Given that the seven species encompass around two-thirds of the European forest area3 and that our models encompass forest regrowth until the age of 40 years, these model predictions cover the approximate magnitude of observed CSs realistically.
In contemporary climate, a substantial portion of this CS in regrowing forests is attributable to conifers, irrespective of the seed provenances being planted (Fig. 2 and Table 2). Under CC (scenario RCP 8.5), the contribution of conifers to the CS, especially of P. abies and P. sylvestris, declines from 75 to 50% if adapted seed sources are planted and from 40 to 15% if local seed sources are used. This is due to their decreasing suitable area, whereas the contribution of broadleaved species to CS increases (Fig. 2 and Table 2). Although the suitable areas for conifers decline in CC, AM contributes more to higher CS of conifers as compared to broadleaved species (Table 3).
In general, all species gained in annual CS when adapted seed provenances were planted but the benefits of AM were more evident under CC (Table 3) despite increasing uncertainties (Table 3). For regrowing conifers, AM results in a gain of 150–200% (RCP 4.5) annual CS as compared to local seed provenances, while for broadleaved species a gain of 130–172% can be expected (Table 3). The higher gains in conifers are in contrast to the lower genetic differentiation among populations as compared to broadleaved trees49. This might be explained either by the lesser coverage of the broadleaved species’ niches by the provenances and trial sites within our dataset (Supplementary Figs. 2 and 3) or by the broader climatic niche of the conifer species resulting in stronger selection and more local adaptations to manifold regional climates or by higher phenotypic plasticity46,50,51,52. The observed CS differences between local and adapted SPC are not restricted to a certain geographic region and adapted SPC outperforms local provenances within all bioclimatic regions of Europe (Supplementary Fig. 12). However, the gains of adapted SPC are more pronounced in central and northern Europe such as in alpine, boreal and atlantic than in the Pannonian and Mediterranean regions mainly because the seven selected species will have only low suitability in southern Europe in CC. Also, the spatial analysis of components of uncertainty did not reveal notable geographical variation. The components contributed by the AM scenario, explained in total 91% of the variance observed in CS (Supplementary Table 13 and Supplementary Figs. 16–18). However, uncertainties due to modelling framework, that is the choice of RCMs and the SDM algorithms were more pronounced in northern and northeastern Europe than elsewhere (Supplementary Table 13 and Supplementary Figs. 19–22).
Our empirical data are based on provenance trials mostly established with seed lots from natural unimproved forests. Tree breeding programmes, where productivity gains of 10–25% are typically achieved within a breeding generation53 allow for a further increase of annual CS if AM is considered54,55. Scientific studies across disciplines such as genecological experiments56, provenance trials57,58,59, dendroclimatic analysis60 and genome studies43,61, although limited in their geographic scale, have indicated that adapted seed provenances are required to avoid maladaptation and mitigate consequences of CC. Our recommendation for AM based on range-wide empirical data confirms this overarching scientific agreement across disciplines regarding CS.
Limitations and conflicts
Our analysis may be subject to potential limitations, which encompass deficiencies in the available data and constraints associated with the models. Although our transfer models are built upon one of the largest continental tree provenance datasets, we were only able to model the provenance transfer of seven main tree species common to central and northern Europe, which dominate, however, within two-thirds of Europe’s forests62. This is because past provenance experiments with a valid representation of full distribution areas are limited to few stand-forming and commercially important species63. Data for rare and scattered distributed trees are rarely available and this limits the extension of our predictions to mixed forest types. Another uncertainty could be the inclusion of data from non-autochthonous seed sources because forests in Europe have been managed for centuries and seed provenances mainly of conifers have been transferred historically64,65. Thus, our data may include a certain amount of non-autochthonous seed sources, where the known seed origin might be different from the original local adaptation. However, a high number of non-autochthonous seed sources in the dataset is expected to weaken the relationship between climate and provenance origin and would result in a possible underestimation of AM effects. Also, we limit the uncertainty of imperfect empirical data by grouping provenances into large-scale SPC. Our analysis also leaves out Mediterranean forests, where similar adaptation lags due to growth decline were reported33,66,67.
The applied transfer models focus on productivity and carbon sequestration for expected future long-term mean climate; therefore, they may not account for extreme climate events or biotic/abiotic disturbances, which pose a serious risk for the future provision of ecosystem services4,5,9,68,69. Moreover, productivity and CS might not be the only traits for which tree populations developed local adaptations. It can be expected that trade-offs exist between productivity and other traits, such as tolerance to frost69, drought or insect outbreaks, which are not explicitly considered here and may reduce the resilience of forests with transferred populations. Therefore, provenance trials need to be further exploited for developing several trait models for AM as a basis for potential implementation. This would complement increasing evidence for local adaptations and future maladaptation in traits related to phenology and drought tolerance23,25,70, which suggests that seed sources from more southern and drier sites are more drought tolerant and may be used also in core areas of species distributions25,71,72. Also, drought recovery and resilience were recently found to be positively correlated to growth and competitive ability in provenance trials established at the warm and dry distribution edges of the species73,74, suggesting that selecting trees for higher carbon stocks under future conditions might also improve drought performance.
Since the main focus of our analysis was to demonstrate the role of AM on CS, simplified assumptions of selecting the single species with the highest climatic suitability in each 1 × 1 km forested grid cell were made. Although it is widely considered that mixed forests may increase productivity and CS through higher yields and can reduce drought sensitivity and mortality15,16, this simplification was required because mixed forests across Europe are composed of different species combinations with species-specific mixture–productivity relationships75,76 and range-wide models to quantify mixture effects are not available yet77.
To quantify the uncertainty of model choice and the source of climate data, best-suited species and provenances were identified for five RCMs, different SDMs and the subsequent URFs. Although the uncertainty of the various model predictions increased from the current climate to the most extreme CC scenario, the cultivation areas for the best-suited tree species and SPC as well as their relative contribution to annual CS remained stable irrespective of the model combination. Our transfer models are applied only within the changing climatic niche of the target species as modelled by state-of-the-art SDMs35. While this allows us to model the forest CS within the climatic space of the past species distribution and avoids predictions beyond the climate conditions of our trial observations (Supplementary Fig. 2 and 3), it does predict changes in tree species and provenances across large geographic distances. Such drastic changes in the forest might have genetic78, ecological79,80, social81,82 and economic16 consequences, which need to be weighed against the outcomes of inaction83. Given the overwhelming evidence from distribution models19,35,84 and increasing forest disturbances6,68, changing forest species compositions and species distributions are the most widely accepted measure to adapt forests to CC7,8,10. However, the limited migration capacities of many species to reach suitable areas22,85 and the increasing population maladaptation23,86 will ultimately limit natural species reshuffling and endanger forest ecosystem services85 if increasingly mismatching local adaptations are ignored. Therefore, inaction in species and provenance selection may increase the vulnerability of current tree populations to CC10,83,87,88 and endanger valuable genetic resources89,90.
Conclusion
Reforestation and restoration needs in European forests6 require decisions on the selection of tree species and seed provenances. Despite evidence of large-scale species shifts and increasing genetic maladaptation in forest trees, there has been little effort to incorporate this knowledge into ongoing tree plantation initiatives, forest restoration programmes and national and transnational regulations. Preventing maladaptation of current forests can provide a major boost to nature-based solutions for CC mitigation such as forest restoration while ensuring resilience and preserving the forest CS12,60,91. Our results provide evidence for strong effects of seed provenance selection on the CS under future climate and demonstrate that improving forest resilience by planting species for future climate needs to be combined with adequate provenance selection to preserve the annual CS of European forests. Changing species composition alone will result in decreasing CS and the risk of failing the European Union target for climate neutrality92. This evidence calls for further research and transnational cooperation to develop and implement AM and to quantify possible trade-offs between productivity and other traits to avoid unknown risks of AM implementation.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The provenance trials dataset is available via Figshare at https://figshare.com/s/98e405d56bb789b08cb0 (ref. 93). The ECLIPS 2.0 dataset is available via Zenodo at https://doi.org/10.5281/zenodo.3952159 (ref. 94).
Code availability
R codes developed for and used in this study are available upon request.
References
IPCC. Summary for Policymakers. In Global Warming of 1.5 °C (eds Masson-Delmotte, V. et al.) 3–24 (Cambridge Univ. Press, 2018).
Bastin, J. F. et al. The global tree restoration potential. Science 365, 76–79 (2019).
State of Forests 2020 (Forest Europe, 2020); https://foresteurope.org/wp-content/uploads/2016/08/SoEF_2020.pdf
Seidl, R., Schelhaas, M. J., Rammer, W. & Verkerk, P. J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Change 4, 806–810 (2014).
Thom, D. & Seidl, R. Natural disturbance impacts on ecosystem services and biodiversity in temperate and boreal forests. Biol. Rev. Camb. Philos. Soc. 91, 760–781 (2016).
Forzieri, G., Dakos, V., McDowell, N. G., Ramdane, A. & Cescatti, A. Emerging signals of declining forest resilience under climate change. Nature 608, 534–539 (2022).
Bolte, A. et al. Adaptive forest management in central Europe: climate change impacts, strategies and integrative concept. Scand. J. Res. 24, 473–482 (2009).
Spathelf, P. et al. Adaptive measures: integrating adaptive forest management and forest landscape restoration. Ann. For. Sci. 75, 55 (2018).
Millar, C. I. & Stephenson, N. L. Temperate forest health in an era of emerging megadisturbance. Science 349, 823–826 (2015).
Jandl, R., Spathelf, P., Bolte, A. & Prescott, C. E. Forest adaptation to climate change—is non-management an option? Ann. For. Sci. 76, 48 (2019).
Bastin, J. F. et al. Tree Restoration Potential in the European Union https://doi.org/10.13140/RG.2.2.24811.67368/1 (FAO and European Commission Directorate General for Environment (DG ENV), 2020).
Matthews, H. D. et al. Temporary nature-based carbon removal can lower peak warming in a well-below 2 °C scenario. Commun. Earth Environ. 3, 65 (2022).
Nabuurs, G. J. et al. First signs of carbon sink saturation in European forest biomass. Nat. Clim. Change 3, 792–796 (2013).
Liang, J. et al. Positive biodiversity–productivity relationship predominant in global forests. Science https://doi.org/10.1126/science.aaf8957 (2016).
Ammer, C. Diversity and forest productivity in a changing climate. New Phytol. 221, 50–66 (2019).
Hanewinkel, M., Cullmann, D. A., Schelhaas, M. J., Nabuurs, G. J. & Zimmermann, N. E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Change 3, 203–207 (2013).
Duveiller, G. et al. Revealing the widespread potential of forests to increase low level cloud cover. Nat. Commun. 12, 4337 (2021).
Thuiller, W., Lafourcade, B., Engler, R. & Araújo, M. B. BIOMOD—a platform for ensemble forecasting of species distributions. Ecography 32, 369–373 (2009).
Dyderski, M. K., Paź, S., Frelich, L. E. & Jagodziński, A. M. How much does climate change threaten European forest tree species distributions? Glob. Change Biol. 24, 1150–1163 (2018).
Thurm, E. A. et al. Alternative tree species under climate warming in managed European forests. For. Ecol. Manag. 430, 485–497 (2018).
Svenning, J. C. & Skov, F. Limited filling of the potential range in European tree species. Ecol. Lett. 7, 565–573 (2004).
Nathan, R. et al. Spread of North American wind-dispersed trees in future environments. Ecol. Lett. 14, 211–219 (2011).
Frank, A. et al. Risk of genetic maladaptation due to climate change in three major European tree species. Glob. Change Biol. 23, 5358–5371 (2017).
Aitken, S. N., Yeaman, S., Holliday, J. A., Wang, T. & Curtis-McLane, S. Adaptation, migration or extirpation: climate change outcomes for tree populations. Evol. Appl. 1, 95–111 (2008).
Isaac-Renton, M. et al. Northern forest tree populations are physiologically maladapted to drought. Nat. Commun. 9, 5254 (2018).
Kremer, A. et al. Long-distance gene flow and adaptation of forest trees to rapid climate change. Ecol. Lett. https://doi.org/10.1111/j.1461-0248.2012.01746.x (2012).
Alberto, F. J. et al. Potential for evolutionary responses to climate change—evidence from tree populations. Glob. Change Biol. https://doi.org/10.1111/gcb.12181 (2013).
Aitken, S. N. & Bemmels, J. B. Time to get moving: assisted gene flow of forest trees. Evol. Appl. 9, 271–290 (2016).
Pedlar, J. H. et al. Placing forestry in the assisted migration debate. Bioscience 62, 835–842 (2012).
Williams, M. I. & Dumroese, R. K. Preparing for climate change: forestry and assisted migration. J. For. https://doi.org/10.5849/jof.13-016 (2013).
McLachlan, J. S., Hellmann, J. J. & Schwartz, M. W. A framework for debate of assisted migration in an era of climate change. Conserv. Biol. 21, 297–302 (2007).
Hällfors, M. H. et al. Coming to terms with the concept of moving species threatened by climate change—a systematic review of the terminology and definitions. PLoS ONE https://doi.org/10.1371/journal.pone.0102979 (2014).
Fréjaville, T., Vizcaíno-Palomar, N., Fady, B., Kremer, A. & Benito Garzón, M. Range margin populations show high climate adaptation lags in European trees. Glob. Change Biol. 26, 484–495 (2020).
Sáenz-Romero, C. et al. Assisted migration of forest populations for adapting trees to climate change. Rev. Chapingo Ser. Cienc. 22, 303–323 (2016).
Chakraborty, D., Móricz, N., Rasztovits, E., Dobor, L. & Schueler, S. Provisioning forest and conservation science with high-resolution maps of potential distribution of major European tree species under climate change. Ann. For. Sci. 78, 26 (2021).
Gunia, K., Van Brusselen, J., Päivinen, R., Zudin, S. & Zudina, E. Forest Map of Europe (European Forest Institute, 2012).
Cook-Patton, S. C. et al. Mapping carbon accumulation potential from global natural forest regrowth. Nature 585, 545–550 (2020).
Chakraborty, D. et al. Selecting populations for non-analogous climate conditions using universal response functions: the case of Douglas-fir in Central Europe. PLoS ONE 10, e0136357 (2015).
Wang, T. et al. Integrating environmental and genetic effects to predict responses of tree populations to climate. Ecol. Appl. 20, 153–163 (2010).
Riahi, K. et al. The Shared Socioeconomic Pathways and their energy, land use and greenhouse gas emissions implications: an overview. Glob. Environ. Change 42, 153–168 (2017).
Diniz-Filho, J. A. F. et al. Partitioning and mapping uncertainties in ensembles of forecasts of species turnover under climate change. Ecography 32, 897–906 (2009).
McGrath, M. J. et al. Reconstructing European forest management from 1600 to 2010. Biogeosciences 12, 4291–4316 (2015).
Browne, L., Wright, J. W., Fitz-Gibbon, S., Gugger, P. F. & Sork, V. L. Adaptational lag to temperature in valley oak (Quercus lobata) can be mitigated by genome-informed assisted gene flow. Proc. Natl Acad. Sci. USA 116, 25179–25185 (2019).
Etterson, J. R., Cornett, M. W., White, M. A. & Kavajecz, L. C. Assisted migration across fixed seed zones detects adaptation lags in two major North American tree species. Ecol. Appl. 30, e02092 (2020).
Mátyás, C. Adaptation lag: a general feature of natural populations (invited lecture). Paper no. 2.226. In Joint Meeting of Western Forest Genetics Association and IUFRO Working Parties, Douglas-fir, Contorta Pine, Sitka Spruce, and Abies Breeding and Genetic Resources 20–24 (Weyerhaeuser Company, 1990).
Leites, L. & Benito Garzón, M. Forest tree species adaptation to climate across biomes: building on the legacy of ecological genetics to anticipate responses to climate change. Glob. Change Biol. https://doi.org/10.1111/gcb.16711 (2023).
Pâques, M. J. Technical Guidelines for Genetic Conservation and use for European Larch (Larix decidua) (EUFORGEN, 2008).
Luyssaert, S. et al. Trade-offs in using European forests to meet climate objectives. Nature 562, 259–262 (2018).
Petit, R. J. et al. Comparative organization of chloroplast, mitochondrial and nuclear diversity in plant populations. Mol. Ecol. https://doi.org/10.1111/j.1365-294X.2004.02410.x (2005).
Valladares, F. et al. The effects of phenotypic plasticity and local adaptation on forecasts of species range shifts under climate change. Ecol. Lett. 17, 1351–1364 (2014).
Kapeller, S., Dieckmann, U. & Schueler, S. Varying selection differential throughout the climatic range of Norway spruce in Central Europe. Evol. Appl. 10, 25–38 (2017).
Müller, M., Kempen, T., Finkeldey, R. & Gailing, O. Low population differentiation but high phenotypic plasticity of European beech in Germany. Forests 11, 1354 (2020).
Jansson, G., Hansen, J. K., Haapanen, M., Kvaalen, H. & Steffenrem, A. The genetic and economic gains from forest tree breeding programmes in Scandinavia and Finland. Scand. J. For. Res. https://doi.org/10.1080/02827581.2016.1242770 (2017).
Milesi, P. et al. Assessing the potential for assisted gene flow using past introduction of Norway spruce in southern Sweden: local adaptation and genetic basis of quantitative traits in trees. Evol. Appl. 12, 1946–1959 (2019).
Poupon, V. et al. Accelerating adaptation of forest trees to climate change using individual tree response functions. Front. Plant Sci. 12, 758221 (2021).
Frank, A. et al. Distinct genecological patterns in seedlings of Norway spruce and silver fir from a mountainous landscape. Ecology 98, 211–227 (2017).
Kapeller, S., Lexer, M. J., Geburek, T., Hiebl, J. & Schueler, S. Intraspecific variation in climate response of Norway spruce in the eastern Alpine range: selecting appropriate provenances for future climate. For. Ecol. Manag. 271, 46–57 (2012).
Berlin, M. et al. Scots pine transfer effect models for growth and survival in Sweden and Finland. Silva Fenn. 50, 1562 (2016).
Pedlar, J. H., McKenney, D. W. & Lu, P. Critical seed transfer distances for selected tree species in eastern North America. J. Ecol. 109, 2271–2283 (2021).
Girardin, M. P. et al. Annual aboveground carbon uptake enhancements from assisted gene flow in boreal black spruce forests are not long-lasting. Nat. Commun. 12, 1169 (2021).
Gougherty, A. V., Keller, S. R. & Fitzpatrick, M. C. Maladaptation, migration and extirpation fuel climate change risk in a forest tree species. Nat. Clim. Change 11, 166–171 (2021).
Brus, D. J. et al. Statistical mapping of tree species over Europe. Eur. J. Res. 131, 145–157 (2012).
Pâques, L. E. (ed.) Forest Tree Breeding in Europe: Current State-of-the-Art and Perspectives (Springer, 2013).
Jansen, S. & Geburek, T. Historic translocations of European larch (Larix decidua Mill.) genetic resources across Europe—a review from the 17th until the mid-20th century. For. Ecol. Manag. https://doi.org/10.1016/j.foreco.2016.08.007 (2016).
Jansen, S., Konrad, H. & Geburek, T. The extent of historic translocation of Norway spruce forest reproductive material in Europe. Ann. For. Sci. 74, 56 (2017).
Benito Garzón, M. & Vizcaíno-Palomar, N. in Pines and Their Mixed Forest Ecosystems in the Mediterranean Basin (eds Ne'eman, G. & Yagil Osem, Y.) 71–82 (Springer, 2021).
Benito-Garzón, M. & Fernández-Manjarrés, J. F. Testing scenarios for assisted migration of forest trees in Europe. New For. https://doi.org/10.1007/s11056-015-9481-9 (2015).
Hlásny, T. et al. Devastating outbreak of bark beetles in the Czech Republic: drivers, impacts and management implications. For. Ecol. Manag. 490, 119075 (2021).
Montwé, D., Isaac-Renton, M., Hamann, A. & Spiecker, H. Cold adaptation recorded in tree rings highlights risks associated with climate change and assisted migration. Nat. Commun. 9, 1574 (2018).
George, J. P. et al. Inter- and intra-specific variation in drought sensitivity in Abies spec. and its relation to wood density and growth traits. Agric. For. Meteorol. 214–215, 430–443 (2015).
Stojnić, S. et al. Variation in xylem vulnerability to embolism in European beech from geographically marginal populations. Tree Physiol. 38, 173–185 (2018).
Bansal, S., Harrington, C. A., Gould, P. J. & St.Clair, J. B. Climate-related genetic variation in drought-resistance of Douglas-fir (Pseudotsuga menziesii). Glob. Change Biol. 21, 947–958 (2015).
George, J. P. et al. Genetic variation, phenotypic stability and repeatability of drought response in European larch throughout 50 years in a common garden experiment. Tree Physiol. 37, 33–46 (2017).
Trujillo-Moya, C. et al. Drought sensitivity of Norway Spruce at the species’ warmest fringe: quantitative and molecular analysis reveals high genetic variation among and within provenances. G3 8, 1225–1245 (2018).
Baeten, L. et al. Identifying the tree species compositions that maximize ecosystem functioning in European forests. J. Appl. Ecol. 56, 733–744 (2019).
Vospernik, S. Basal area increment models accounting for climate and mixture for Austrian tree species. For. Ecol. Manag. 480, 118725 (2021).
Pretzsch, H., Forrester, D. I. & Rötzer, T. Representation of species mixing in forest growth models: a review and perspective. Ecol. Model. https://doi.org/10.1016/j.ecolmodel.2015.06.044 (2015).
Grummer, J. A. et al. The immediate costs and long-term benefits of assisted gene flow in large populations. Conserv. Biol. 36, e13911 (2022).
Kranabetter, J. M., Stoehr, M., & O’Neill, G. A. Ectomycorrhizal fungal maladaptation and growth reductions associated with assisted migration of Douglas-fir. New Phytol. 206, 1135–1144 (2015).
Winder, R. S., Kranabetter, J. M. & Pedlar, J. H. in Soils and Landscape Restoration (eds Stanturf, J. A. & Callaham, Mac A.) 275–297 (Academic Press, 2021).
Klenk, N. L. The development of assisted migration policy in Canada: an analysis of the politics of composing future forests. Land Use Policy 44, 101–109 (2015).
Pelai, R., Hagerman, S. M. & Kozak, R. Whose expertise counts? Assisted migration and the politics of knowledge in British Columbia’s public forests. Land Use Policy 103, 105296 (2021).
Rodríguez-Labajos, B. Climate change, ecosystem services and costs of action and inaction: scoping the interface. WIRES Clim. Change https://doi.org/10.1002/wcc.247 (2013).
Sykes, M. T., Prentice, I. C. & Cramer, W. A bioclimatic model for the potential distributions of north European tree species under present and future climates. J. Biogeogr. 23, 203–233 (1996).
Mauri, A. et al. Assisted tree migration can reduce but not avert the decline of forest ecosystem services in Europe. Glob. Environ. Change 80, 102676 (2023).
St Clair, J. B. & Howe, G. T. Genetic maladaptation of coastal Douglas-fir seedlings to future climates. Glob. Change Biol. 13, 1441–1454 (2007).
Hornsey, M. J. & Fielding, K. S. Understanding (and reducing) inaction on climate change. Soc. Issues Policy Rev. 14, 3–35 (2020).
Kracke, I., Essl, F., Zulka, K. P. & Schindler, S. Risks and opportunities of assisted colonization:the perspectives of experts. Nat. Conserv. 45, 63–84 (2021).
Schueler, S. et al. Vulnerability of dynamic genetic conservation units of forest trees in Europe to climate change. Glob. Change Biol. 20, 1498–1511 (2014).
Petit-Cailleux, C. et al. Tree mortality risks under climate change in Europe: assessment of silviculture practices and genetic conservation networks. Front. Ecol. Evol. 9, 706414 (2021).
Sha, Z. et al. The global carbon sink potential of terrestrial vegetation can be increased substantially by optimal land management. Commun. Earth Environ. 3, 8 (2022).
Fit for 55: Parliament Agrees to Higher EU Carbon Sink Ambitions by 2030 (European Parliament, 2022); https://www.europarl.europa.eu/news/en/press-room/20220603IPR32133/fit-for-55-parliament-agrees-to-higher-eu-carbon-sink-ambitions-by-2030
Assisted translocation of tree populations preserves the European forest carbon sink in climate change. Figshare https://figshare.com/s/98e405d56bb789b08cb0 (2022).
Chakraborty, D., Dobor, L., Zolles, A., Hlásny, T. & Schueler, S. High-resolution gridded climate data for Europe based on bias-corrected EURO-CORDEX: the ECLIPS-2.0 dataset. Zenodo 10.5281/zenodo.3952158 (2020).
Acknowledgements
This research was supported by the INTERREG Central Europe Programme, Project SUSTREE: Conservation and sustainable utilization of forest tree diversity in climate change (grant no. CE614) and the SUPERB project funded by the European Union Horizon 2020 research and innovation programme under grant agreement no. 101036849. We are thankful to G. Schnabel (BFW) for help in graphical layout. We also acknowledge the support of COST Action CA19128 (PEN-CAFoRR) ‘Pan-European Network for Climate Adaptive Forest Restoration and Reforestation’ for co-authors G.B., M.I. and M.K. Funding for M.L. was received from the EEA/Norway grants 2014–2021 and the Technology Agency of the Czech Republic. We recognize the IUFRO Task Forces on Forest Adaptation and Restoration under Climate Change and Transforming Forest Landscapes for Future Climates and Human Well-Being represented by co-author A.B. We also acknowledge the Slovenian Research and Innovation Agency through research programme P4-0107 and research project V4-2222 co-financed by the Ministry of Agriculture, Forestry and Food received by co-author G.B. A.K. and A.D. received funding support from the EC-Grants FAIROAK (no. PL95-0297), MOTIVE (no. 226544) and FORGER (no. 289119) We are also thankful to all the scientists and institutions that established and maintained provenance trials throughout the past century and supported the data collection within the SUSTREE project.
Author information
Authors and Affiliations
Contributions
D.C. conceived the idea of the paper, formulated research questions, developed and carried out analysis and wrote the paper. A.C. supported analysis in the revised paper. D.B., G.B., E.C., A.H., AD., M.I., R.L., G.M., K.P., E.P., R.S., T.M.S., T.S., A.S., S. Stojnic and K.V. provided provenance trial data. M.B.G., A.K. and L.N. provided provenance trial data and edited the paper. A.B. provided computing resources and edited the paper. R.B., J.C., J.G., J.P.G., M.K., J.K. and J.S. contributed to running URF models. M.L. contributed to running URF models and edited the paper. A.S. carried out paper revisions. S. Schueler conceived the idea of the paper and analysis, obtained research funding, contributed trial data and wrote the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Climate Change thanks Sally Aitken, Gert-Jan Nabuurs and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary Information
Supplementary data, methods, results and references, including Table 1–13 and Figs. 1–22.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chakraborty, D., Ciceu, A., Ballian, D. et al. Assisted tree migration can preserve the European forest carbon sink under climate change. Nat. Clim. Chang. 14, 845–852 (2024). https://doi.org/10.1038/s41558-024-02080-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41558-024-02080-5
- Springer Nature Limited
This article is cited by
-
Tree movements promote carbon sink
Nature Climate Change (2024)