
Abstract
Cold physical plasma has been suggested as a powerful new tool in oncology. However, some cancer cells such as THP-1 leukaemia cells have been shown to be resistant towards plasma-induced cell death, thereby serving as a good model for optimizing plasmas in order to foster pro-apoptotic anticancer effects. A helium/oxygen radio frequency driven atmospheric plasma profoundly induced apoptosis in THP-1 cells whereas helium, humidified helium, and humidified helium/oxygen plasmas were inefficient. Hydrogen peroxide – previously shown as central plasma-derived agent – did not participate in the killing reaction but our results suggest hypochlorous acid to be responsible for the effect observed. Proteomic analysis of THP-1 cells exposed to He/O2 plasma emphasized a prominent growth retardation, cell stress, apoptosis, and a pro-immunogenic profile. Altogether, a plasma setting that inactivates previously unresponsive leukaemia cells is presented. Crucial reactive species in the plasma and liquid environment were identified and discussed, deciphering the complexity of plasma from the gas phase into the liquid down to the cellular response mechanism. These results may help tailoring plasmas for clinical applications such as oxidation-insensitive types of cancer.
Similar content being viewed by others


Introduction
Firmly connecting with redox biology, therapeutical effects of cold plasma-generated reactive molecules are investigated in the field of plasma medicine1. The strong advantage of plasma is the parallel deposition of different biologically active reactive species in a localized manner2. In the plasma gas phase, this includes for example hydroxyl radical, nitric oxide, and atomic oxygen3. In plasma-treated liquids, chemistry is further complexed, and typical molecules detected include superoxide anion, hydrogen peroxide, and peroxynitrite4. With adequate concentrations, these species can overwhelm the cells’ antioxidative response, effectively mediating pro-apoptotic redox signalling responses5. Many tumours display an inequity in their redox balance, rendering them more receptive towards oxidation-induced cell death compared to non-malignant cells6. Accordingly, a number of drugs have been tested in clinical trials aiming at further disturbing the redox-balance of cancers7, ultimately inducing apoptosis8. Thus, plasmas applications have been proposed to be a possible asset in oncology as well, as killing has been achieved for various types of cancers in vitro 9,10,11 and in vivo 12,13,14. Although primary monocytes are susceptible to plasma-induced apoptosis15, some malignant cells such as THP-1 leukaemia cells effectively withstand plasma-induced apoptosis16, and rather respond via growth deceleration17. They, therefore, serve as an ideal target to identify which parameters may render a plasma source more effective for oncological applications in oxidation-resistant cancer cells.
Different parameters can be adjusted to alter the plasma generation and thus the reactive species composition. For example, humidity in the feed argon gas was identified to play a highly critical role18. The humidification of a noble gas dramatically increase hydrogen peroxide (H2O2) production in the gas and liquid phase19 whereas addition of oxygen (O2) or nitrogen (N2) enhances the generation of reactive oxygen and nitrogen species (ROS, RNS)4, respectively. A helium (He)-driven atmospheric pressure plasma jet (µAPPJ) in its COST-jet version was used which has been characterized towards its physical and antimicrobial properties20,21,22. To investigate the biological effects of feed gas alterations, it was humidified and/or spiked with molecular oxygen (O2) with the idea of altering liquid chemistry, redox reactions, and subsequently eradication of the tumour target cells. In this study, we analysed cell morphology and counts, metabolic activity, apoptosis, and the THP-1 proteome on the one hand, and investigate the plasma-treated liquids on the other hand to determine the mechanism of action as function of the feed gas compositions.
Results
The feed gas composition-dependent THP-1 cell inactivation was mediated via apoptosis
THP-1 cells are known to strongly persist oxidative stress-induced killing. The aim of this work was to identify a feed gas composition that may foster cold plasma-induced eradication of these cells to substantiate its potential use in tumour therapy. THP-1 cells were exposed to a radiofrequency driven plasma jet utilizing He, He/0.6% O2, He/0.15% H2O, or He/0.6% O2/0.15% H2O as feed gases (Fig. 1), and were assayed 24 h later. The conversion of the resazurin to its fluorescent product resorufin via nicotinamide adenine dinucleotide phosphate reduction equivalents identifies the overall metabolic activity of given number of cells. A significantly decreased metabolism could be observed in cells treated with He/O2 plasma but not with any of the other feed gas conditions used (Fig. 2a). Flow cytometry was performed to determine the mitochondrial activity on a single cell level using mitotracker orange dye (Fig. 2b). Highly fluorescent once incorporated into mitochondria, its fluorescence decreases if the mitochondrial membrane potential is lost23, which indicates cell death. Only cells exposed to the He/O2 plasma showed a significant impairment (Fig. 2c) which may have contributed to the overall decreased metabolic activity seen with the resazurin dye. It was asked next whether this finding may also be attributed to a decrease in total cell numbers in the appropriate forward scatter (FSC)/side scatter (SSC) gate (Fig. 2d). Again, only the He/O2 plasma condition led to a significant reduction of cells (Fig. 2e). The data of viable cell counts (Fig. 2e) and metabolic activity (Fig. 2a) are not congruent. This may be a consequence of apoptosis being an active and energy consuming process24 together with the observation that plasma treatment may increase activity in viable THP-1 cells16. Altogether, He/O2 but no other plasma conditions efficiently reduced THP-1 cell metabolic activity by inducing a down-modulation of mitochondrial activity and via reduction of total cell counts. In none but the He/O2 condition, cytotoxicity could be observed (Fig. 2). We, therefore, assessed morphological features of control (Fig. 3a) and He/O2 plasma-treated (Fig. 3b) THP-1 cells after 24 h using image cytometry. Treated but not control cells showed features of apoptosis such as membrane blebbing, nuclear defragmentation with similar side-scatter profiles of dead cells as seen with traditional cytometry (Fig. 2d). To further validate the apoptotic process, THP-1 cells were stained for activated executioner caspases 3/7 (Fig. 3c), and He/O2 but not He plasma-treated cells were shown to be significantly positive (Fig. 3d). Linking to these apoptotic events, concentrations of interleukin (IL)-8 were significantly decreased in He/O2 plasma-treated samples as well (Fig. 3e).

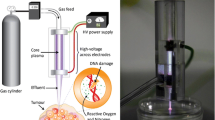
Image of the plasma source and its schematic overview. (a) Schematic representation of the plasma source with the bubbler and gas connections. Additionally, one well of a 24 well plate with the relative position of the jet are shown as well. (b) Photograph of the atmospheric pressure helium plasma jet (µAPPJ).

He/O2 but not He, He/H2O, or He/O2/H2O plasma inactivated THP-1 cells. THP-1 cells were exposed to cold physical plasma under different feed gas conditions, and assayed after 24 h. (a) Overall metabolic activity was significantly decreased in the He/O2 condition, as measured by resazurin to resorufin transformation. (b) Representative histograms of mitotracker fluorescence for each gas condition. (c) Mitotracker fluorescence quantification revealed significant reduction of mitochondrial activity in the He/O2 condition. (d) Representative FSC/SSC dot plots with the appropriate live and cell count gate. (e) Live cell count quantification demonstrated a significant decrease with the He/O2 condition. Data are one representative (b,d) or presented as mean + S.D. (a,c,e) of three experiments. Statistical differences (***p < 0.001) were determined using t-test.

The mode of cytotoxicity of He/O2 plasma in THP-1 cells was apoptosis. (a) Control THP-1 cells showed a round morphology (BF, bright field, left array) with intact nuclei (DRAQ5, middle panel) and characteristic side scatter patterns (SSC, right panel) as measured using image cytometry. (b) Plasma-treated THP-1 cells displayed altered morphological features with membrane blebbing and vacuolization (BF) and segmented nuclei (DRAQ5). (c) Representative histograms of activated caspases 3/7 in plasma-treated cells. (d) Quantification of percent of cells being positive for active caspases 3/7 with a significant increase in He/O2 but not the He plasma condition. (e) THP-1 cell culture supernatant was investigated for IL-8 24 h after exposure to plasma generated using different feed gas compositions. Final concentrations where normalized to each gas-treated control, respectively. He/O2 but not any other gas plasma significantly decreased IL-8 concentrations in the supernatant of THP-1 cells. Data are one representative (a,b,c) or mean (d,e) + S.D. of three experiments. Statistical analysis was determined using the t-test (***p < 0.001).
THP-1 cell killing in the He/O2 condition was mediated via a non-H2O2 mechanism
THP-1 cells showed a high susceptibility towards He/O2 but not He, He/H2O, and He/H2O/O2 plasma. H2O2 was previously shown to be an important mediator in plasma cytotoxicity25. Yet, its concentration was decreased in the He/O2 plasma toxic condition compared to the helium plasma (Fig. 4a). Additionally, the enzymatic antioxidant catalase, a potent scavenger of H2O2 26, did not inhibit the cytotoxic effect of He/O2 plasma (Fig. 4b). H2O2 contributed, therefore, unlikely to the killing mechanism. To test whether the responsible reactive molecule would be rather short or long lived, THP-1 cells were exposed to plasma-treated medium in presence of absence of the potent thiol-containing non-enzymatic antioxidants N-acetyl-cysteine (NAC) or glutathione (GSH). GSH is among the most abundant proteins in cells, and its oxidized dimer GSSG in readily replenished in cells by glutathione reductase27. Intracellularly, NAC serves as a GSH precursor, and its extracellular antioxidant activity is superior to that of GSH28. Similar to direct treatment, helium plasma-treated liquid did not affect cell viability (Fig. 4b). On the contrary, He/O2 plasma-treated liquid exhibited significant cytotoxic activity, although to a significantly lesser extent compared to direct plasma exposure (Fig. 4b). The toxic effects were abrogated in the presence of NAC but not glutathione GSH. To identify the species possibly entangled in this reaction, two dyes (3′-(p-aminophenyl) fluorescein, APF; or 3′-(p-hydroxyphenyl) fluorescein, HPF) were plasma-treated. In contrast to HPF, APF has been reported to distinguish for hypochlorous acid (HOCl)29 and has been used and validate throughout several studies in cells and liquids30,31,32. With APF but not HPF, fluorescence was markedly increased in the He/O2 compared to the He plasma condition (Fig. 5a). NAC and GSH but not catalase partially protected APF from oxidation. These experiments suggested HOCl to be generated in the liquid by He/O2 plasma treatment. HOCl is known to scavenge H2O2. A known concentration of H2O2 was experimentally added to PBS and chloride-free phosphate buffer and both solutions were exposed to plasma (Fig. 5b). With He plasma, H2O2 concentrations increased above that added concentration in both solutions as expected. This was also obtained for He/O2 plasma treating chloride free-buffer, as HOCl cannot be generated without chloride ions. However, He/O2 treatment of chloride-containing buffer fully consumed the experimentally added H2O2, further suggesting the production of HOCl under this condition. It was hypothesized that HOCl in chloride containing liquids can derive especially from atomic oxygen but also singlet oxygen coming from the plasma gas phase or generated in the liquid. Using a fluorescent singlet oxygen sensor, a moderate but significant increase in singlet oxygen in the liquid was found only in the pro-apoptotic He/O2 plasma but no other gas condition (Fig. 5c), underlining that changes of reactive components in that particular plasma composition were crucial for the observed effects.

Not H2O2 but another long-lived oxidant was responsible for THP-1 cell killing. (a) H2O2 was assessed in plasma-treated cell culture medium using different feed gas conditions, and He plasma showed the highest production with catalase scavenging H2O2 in any feed gas condition. (b) THP-1 cells were either exposed to He or He/O2 plasma or to He or He/O2 plasma-treated medium which was immediately added afterwards using different feed gas conditions and in presence or without of either catalase, NAC, or GSH. Cells were assayed for metabolic activity 24 h later, and showed a significant reduction (***) in presence of catalase or GSH but not NAC. This effect was less pronounced and differed significantly (###) for plasma-treated medium compared to direct treatment in the He/O2 condition. Data are presented as mean + S.D. of two experiments. Statistical differences (***/### p < 0.001) were determined using 1-way ANOVA within each gas condition and plasma treatment regimen, and t-test to compare individual values for a single treatment regimen within each gas condition.

He/O2 plasma generated hypochlorous acid in the liquid via gaseous phase oxygen species. (a) Cell culture medium as loaded with either APF or HPF. Scavengers were added or not, solutions were plasma-treated using either He or He/O2 as feed gas, and fluorescence of the redox dyes was acquired which was normalized to each respective gas control. APF in general gave a stronger fluorescence signal which was increased in the He/O2 condition but not using the He plasma. NAC partially scavenged signals. (b) Fifty micromolar H2O2 was added to PBS and chloride-free phosphate buffer (PB). Solutions were plasma-treated using either He or He/O2 plasma. The latter scavenged experimentally added H2O2 in PBS but not PB. (c) Singlet oxygen sensor green was added to cell culture medium which was subsequently exposed to either He, He/O2, He/H2O, or He/H2O/O2 plasma. Fluorescence was acquired and normalized to untreated gas control, and only He/O2 plasma differed significantly. Data are presented as mean + S.D. of two experiments. Statistical difference (***p < 0.001) was determined using t-test.
Pro-apoptotic and growth deceleration proteins regulated in He/O2 plasma-treated cells
THP-1 cells showed a high susceptibility towards He/O2 plasma. Using proteomic technology, up or down-regulation of proteins in the cellular fraction of THP-1 cells 4 h after plasma treatment with either He or He/O2 with or without the presence of NAC was assessed against gas-treated control cells. Gene ontology analysis identified binding (37%), catalytic activity (33%), structural molecule activity (15%), transporter activity (5%), and receptor activity (4%) as main molecular function categories of proteins that were significantly regulated in He/O2 plasma-treated compared to gas-treated control cells. Six representative significantly up or down-regulated proteins for the He/O2 treated cells are given (Table 1) together with the regulation in the other three samples types. Addition of NAC abrogated the regulation of any protein in plasma-treated samples to the level of untreated gas controls. This corroborates the findings with metabolic activity where NAC fully protected against toxic He/O2 plasma activity on THP-1 cells. Consistent with flow cytometric active caspase 3 detection, an upregulation (+11.1-fold) of this protein was also determined in the cellular fraction using mass spectrometry. Other proteins involved in apoptosis, transport or cell growth were also strongly regulated, for example, A-kinase anchor protein 8-like, Exportin-T, Telomeric repeat-binding factor 2-interacting protein, Cell and growth-regulating nucleolar protein (LYAR), and U4/U6.U5 tri-SNRNP -associated protein 1. A pro-immunogenic protein (Squamous cell carcinoma antigen recognized by T-cells 3) was increased as well.
Discussion
Cold physical plasma-derived reactive species are a promising new tool in oncology33. However, some tumours are highly resistant to oxidative stress-induced cell death34. By modulating the reactive species output, it is, therefore, important to identify plasma-settings that also sensitize these cancer cells to apoptosis. Humidification of the carrier gas was previously described to potentiate HO. and H2O2 generation of an atmospheric pressure argon plasma jet manifold18. These plasma-derived molecules are strong anticancer agents35. Yet, THP-1 cells resist high H2O2 concentrations and long plasma treatment times16, 17, 36. By admixing low amounts of oxygen to the feed gas of a helium plasma jet, a redox route was identified that is particularly effective in THP-1 cell killing. In line with classical hallmarks of apoptosis, treated cells showed membrane blebbing and executioner caspase 3 activation37. Interestingly, mitotracker orange fluorescence was also decreased in the viable cell portion. This argues for a destabilized mitochondrial membrane potential23 i.e. at least partial damage in He/O2 plasma-treated cells. The reduction in viability was also reflected by a decrease in constitutively expressed38 IL-8. This is interesting as oxidant exposure with only moderate apoptotic effects strongly increased IL-8 release in THP-1 cells16, 39 and other myeloid cells undergoing plasma-induced cell death40. IL-8 release is subject to redox control41 and a key molecule in regulating inflammation in wound healing42 and cancer43. Its release by e.g. monocytes/macrophages attract neutrophils to clear infection but also spur inflammation44.
Using proteomics, we identified proteins involved in growth retardation and cell death in He/O2 plasma-treated THP-1 cells. Protein phosphatase 1 regulatory subunit 27 negatively regulates phosphatases, and its significantly decreased recovery (−3.4-fold) reasons for sensation of cell stress resulting in target protein phosphorylation45. Cell and growth-regulating nucleolar protein was significantly increased (+4.1-fold). It is a crucial regulator of ATR-Chk146 which is central in sensing redox stress as well47. This hampers cell cycling that is reflected by significant down-regulation of A-kinase anchor protein 8-like (−8.3-fold) which initiates initial phases of DNA replication48. Vice versa, U4/U6.U5 tri-SNRNP-associated protein 1, which is known to induced cell cycle arrest49, was upregulated (+2.6-fold). Paralleling growth deceleration, Exportin-1, important in exporting tRNAs from the nucleus to support protein translation and overexpressed in many tumors due to high proliferation rates50, was significantly down-regulated (−3.4-fold). Similarly, the down-regulation of Telomeric repeat-binding factor 2-interacting protein 1 (−3.4-fold) suggest growth retardation as it phosphorylates the inhibitor of NfκB thereby fostering cellular activation51. The protein LYAR is central for cellular growth52 and was significantly reduced (−2.4-fold) in plasma-treated cells. Vice versa, the proliferation marker Ki-67 was found to be decreased (−2.3-fold) whereas levels of cell death executioner caspase 3 drastically increased (+11.4-fold), in line with the flow cytometry experiments. This is underscored by elevated levels of Mitogen-activated protein kinase kinase kinase MLT (+2.1-fold), an upstream regulator Jnk and p38 known to be pro-apoptotic also in other cancer cells53. Interestingly, Histone acetyltransferase p300, a current target for anticancer therapy54, was increased in He/O2 plasma-treated THP-1 cells. Also of potential therapeutic value is the massive elevation of Squamous cell carcinoma antigen recognized by T-cells 3 (SART3) which has been used to potentiate T cell responses in vaccination trials in cancer patients (+11.1-fold)55. It is overexpressed in the majority of adenocarcinomas and squamous cell carcinomas in different tissues as well as in leukemia but not non-malignant cells56. Several reports conclude that SART3 is among a few targets found in many cancers with value in immunotherapy57,58,59. Altogether, the toxic plasma condition in THP-1 leukemia not only induced pro-apoptotic proteins but also suggested an enhanced immunogenicity of treated cells.
Significant cell responses were found to be present only in the He/O2 but not in any other feed gas condition. Although not necessarily the single responsible agent60,61,62, H2O2 has been determined to be central in plasma-mediated cytotoxicity in a number of studies63,64,65,66. Here, its direct effects were negligible as results with catalase suggest. Instead, we suspect HOCl to be the active cytotoxic agent for a number of reasons. NAC, an efficient scavenger of HOCl67, abrogated the He/O2 plasma’s cytotoxicity. HOCl is also a known scavenger of H2O2 68. A full decrease of experimentally added H2O2 was only apparent in the toxic He/O2 but not the non-toxic He plasma conditions. Moreover, scavenging was abolished in the absence of chloride which is apparently essential for the generation of HOCl in liquids69. Both APF and HPF are reported to sense HO. and ONOO− 70. APF but not HPF is sensitive for HOCl29 and used to detect halogenated acids such as HOCl in phagocytes32 and leukemic cells71. He/O2 but not He plasma increased APF but not HPF fluorescence. These results suggest the presence and activity of HOCl, although a previous report using another plasma jet suggested RNS to be important in THP-1 cell inactivation72. Another possibility is that HOCl acted on cell culture proteins that then have mediated cytotoxic effects. In mammals, HOCl is physiologically generated by myeloperoxidase primarily expressed by neutrophils73. HOCl especially serves to destruct phagocytosed bacteria74. It is also present extracellularly, for example as a by-product of neutrophil extracellular traps generated during infection75. HOCl is highly reactive and capable of inactivating protease inhibitors and lysozyme76. Thus, it has an important role in fighting bacteria and spurring inflammation. Nonetheless, HOCl is readily detoxified by cellular thiols77, albumin78, and cross linking of biological molecules via non-disulfide bonds79, effectively limiting its damage to host cells.
The addition of oxygen to dry helium feed gas was found to be central for plasma to inactivate THP-1 cells in a HOCl-dependent manner. HOCl generation is thought to be mediated via atomic oxygen (1):
Underlining this notion80, O densities in the gas phase of this plasma source have been reported to be markedly elevated in the effective He/O2 plasma (1015 cm−3)81 compared to He and He/H2O plasma (1013 cm−3)82. Hypochlorous acid can be formed via the reaction of O with Cl−, and our experiments using chloride-free buffer support the significance of this route in our study. In humidified feed gas, the generation of OH· and not O is dominant83 which is reflected in the relatively H2O2 resistant THP-1 cells17 not being strongly affected with these plasma conditions in the present study. Importantly, O shows a good solubility in liquids as previous work using phenol as target suggest83 which makes its important role in HOCl generation plausible.
A further source of reactivity in the He/O2 gas mixture could be other excited molecular oxygen species, especially the singlet delta oxygen O2(1Δg) with a gas phase density of around 4 × 1014 cm−3 with similar power and gas conditions as used in this work84. Yet, its density was previously found to decrease with increasing O2 concentration (due to fast quenching in collisions with O atoms in the gas phase), limiting its possible impact. Nonetheless, in He/O2 plasma but no other feed gas condition, singlet delta oxygen was modestly increased in treated medium containing L-histidine, which was technically suboptimal as it can quench singlet oxygen85. According to the following net reaction (2):
Hypochlorous acid oxidizes hydrogen peroxide yielding chloride ions86, water, and singlet delta oxygen (O2(1Δg)). Together with the absence of H2O2 this notion further emphasizes the role of HOCl as an active agent under the discussed circumstances. Consecutively, O2(1Δg) created in this secondary reaction may contribute to the observed effect, but considering its very short lifetime in aqueous, neutral liquids (few µs) compared to HOCl (several minutes)87, to a limited extent only. Notably, ozone production also increases 10-fold in the toxic oxygen setup81. Yet, and in contrast to treating dry surfaces88, modelling studies grant only a minor role to ozone in the liquid89 as it dissolves badly and no reactions with phenols could be detected83.
Conclusion
Cold physical plasma has been suggested to be an option for cancer therapy, but some cancers are highly resistant against plasma-induced peroxide stress. By modulating the feed gas of a helium-driven plasma jet by the addition of oxygen, it was demonstrated that the chemistry can be tuned towards other species such as hypochlorous acid. With similar treatment times, this condition severely hampered cell activity and growth by inducing apoptosis whereas plasma treatments dominated by hydrogen peroxide did not. Additionally, immune stimulatory proteins were found to be elevated. Understanding the biological impact of gas and liquid phase plasma chemistry will help improving the plasmas’ efficacy in order to tailor them to the therapeutic applications’ needs.
Materials and Methods
Plasma Source
The COST jet, a proposed reference jet in the European Cost action MP1101 to compare effects of plasma treatments among different laboratories, was used as plasma source (Fig. 1a)20. Helium (99.9999%; Air liquide, France) at a flow of 1.4 slm was used as feed gas. In some experiments, it was admixed with either 0.6% oxygen or humidified helium with approximately 1450 ppm of H2O, or both. Humidification was achieved by bubbling 0.3 slm of helium flow through 400 ml of double distilled water and mixing it with 1.1 slm of dry Helium to obtain the total flow of 1.4 slm. (Fig. 1b). Calibrated, USB-controlled flow controllers (MKS instruments, Germany) were used to set and monitor all feed gas conditions.
Cell culture and plasma treatment
The leukaemia cell line THP-1 (ATCC TIB-202, Germany) were cultured in fully supplemented RPMI1640 cell culture medium containing 10% fetal bovine serum, 2% glutamine, and 1% penicillin-streptomycin (all Sigma, Germany) in an incubator (Binder, Germany) at 37 °C, 95% relative humidity, and 5% CO2. For plasma-treatment, 1 × 105 cells in 250 µl of medium were seeded in each well of a 24-well plate (Nunc, Denmark). To some wells, the H2O2 scavenging enzyme catalase (20 µg/ml), the antioxidant GSH (1 mM) or the antioxidant NAC (2 mM) was added (all Sigma). Cells were directly exposed to either each plasma condition or their respective gas controls (plasma off). Alternatively, 1 × 105 cells in 125 µl of cell culture medium received 125 µl of 250 µl medium (indirect approach) plasma-treated for the double amount of time for comparison. Following exposure to plasma or plasma-treated liquid, cells were returned to the incubator for 20 h or 24 h depending on the subsequent analysis.
Cellular analysis and viability
THP-1 cells were investigated on a number of properties, such as cell morphology, total metabolic activity, total cell counts, individual mitochondrial activity, and caspase 3/7 activity. For assessment of morphology 24 h after treatment, THP-1 cells were collected and stained with DRAQ5 (BioLegend, UK) prior to image acquisition using an ImageStream Mark X (Merck-Millipore, USA). Cells representative for viable and apoptotic events were analysed using Ideas 6.2 software (Merck-Millipore). For assessment of total metabolic activity 20 h after treatment, resazurin (Alfa Aesar, USA) was added to the wells, and cells were incubated for another 4 h. Supernatants were transferred to 96-well plates and fluorescence was acquired using a microplate reader with λex 530 nm and λem 590 nm (Tecan, Switzerland). To assess mitochondrial activity 24 h after treatment, cells were stained with 500 nM mitotracker orange (CMXRos; life technologies, USA) for 20 min at 37 °C, and mitotracker fluorescence of cells in the respective gate was acquired using an CytoFlex cytometer (Beckman-Coulter, USA). Mitotracker orange is retained in mitochondria due to their chloromethyl group forming a covalent bond with thiols90. Moreover, the dye fluoresces to a lesser extend upon mitochondrial damage and membrane depolarization23. For total cell counts, cells were aliquoted from the 24-well plates into 96-well plates 24 h after treatment and counted with an attune flow cytometer (Applied Biosystems, USA) capable of measuring absolute particle concentrations. To assess apoptosis 24 h after treatment, THP 1 cells were collected, washed, and stained for 30 min at 37 °C with caspase 3/7 indicator (life technologies). The percentage of cells staining caspase positive was quantified using the CytoFlex cytometer.
Redox-sensitive probes
Fully supplemented RPMI1640 medium was loaded with 1 µM singlet oxygen sensor, or PBS was loaded with 1 µM of either the fluorescent redox indicators APF or HPF (life technologies). Both APF and HPF can be oxidized by hydroxyl radicals and peroxynitrite but not hydrogen peroxide whereas only APF is sensitive towards hypochlorous acid29. Two-hundred and fifty microliter was added to each well of a 24-well plate, and wells either received plasma treatment or were left untreated. Subsequently, the liquid was aliquoted into 96-well plates, and fluorescence was acquired using a microplate reader with λex 485 nm and λem 535 nm (Tecan). In order to quantify hydrogen peroxide (H2O2), plasma-treated cell culture medium or plasma-treated chloride-free phosphate buffer was incubated with amplex ultra red (life technologies) according to the vendor’s instructions, and subsequently quantified against an H2O2 standard using a Tecan microplate reader with λex 530 nm and λem 590 nm. Alternatively, cell culture medium was spiked with a known concentration of H2O2 and the scavenging activity of plasma-introduced reactive components on H2O2 was assessed in a similar manner.
Interleukin 8 and global protein expression
Twenty-four hours after plasma treatment, IL-8 in supernatants of THP-1 cells was quantified using ELISA (BioLegend), and concentrations were normalized to each respective gas control. Four hours after treatment, either eight replicates of untreated or plasma-treated THP-1 cells were pooled into micro-centrifuge tubes, washed, and stored at −80 °C. Global protein expression was carried out as previously described91. Briefly, peptides were separated by nano-liquid chromatography (Dionex Ultimate 3000; PepMap RSLC column, 75 µm ID/15 cm), and eluates were ionized by electrospray ionization and analysed by high-resolution mass spectrometry (QExactive, Thermo, USA) mass spectrometer. Data processing was done using Proteome Discoverer 1.4 software (Thermo). Protein candidates were selected upon their involvement in pathways of metabolisms and proliferation as well as on statistical criteria (≥ ± 2.0-fold expression). Data were also analysed with Ingenuity Pathway Analysis software (IPA, Qiagen) and free web based applications (PANTHER and Universal Protein Resource).
Statistics
Statistical analysis was carried out using prism 7.02 (graph pad software, USA). A significance level of α = 0.05 was set for all statistical analysis, and significantly different data were marked with asterisks (*p < 0.05; **p < 0.01; ***p < 0.001).
References
Graves, D. B. The emerging role of reactive oxygen and nitrogen species in redox biology and some implications for plasma applications to medicine and biology. J. Phys. D: Appl. Phys. 45, 263001, doi:10.1088/0022-3727/45/26/263001 (2012).
Dunnbier, M. et al. Ambient air particle transport into the effluent of a cold atmospheric-pressure argon plasma jet investigated by molecular beam mass spectrometry. J. Phys. D: Appl. Phys. 46, 435203, doi:10.1088/0022-3727/46/43/435203 (2013).
Schmidt-Bleker, A., Reuter, S. & Weltmann, K. D. Quantitative schlieren diagnostics for the determination of ambient species density, gas temperature and calorimetric power of cold atmospheric plasma jets. J. Phys. D: Appl. Phys. 48, 175202, doi:10.1088/0022-3727/48/17/175202 (2015).
Jablonowski, H. & von Woedtke, T. Research on plasma medicine-relevant plasma–liquid interaction: What happened in the past five years? Clin. Plas. Med. 3, 42–52, doi:10.1016/j.cpme.2015.11.003 (2015).
Limon-Pacheco, J. & Gonsebatt, M. E. The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress. Mutat. Res. 674, 137–147, doi:10.1016/j.mrgentox.2008.09.015 (2009).
Acharya, A., Das, I., Chandhok, D. & Saha, T. Redox regulation in cancer: a double-edged sword with therapeutic potential. Oxid. Med. Cell. Longev. 3, 23–34, doi:10.4161/oxim.3.1.10095 (2010).
Sainz, R. M., Lombo, F. & Mayo, J. C. Radical decisions in cancer: redox control of cell growth and death. Cancers (Basel) 4, (442–474 (2012).
Trachootham, D., Alexandre, J. & Huang, P. Targeting cancer cells by ROS-mediated mechanisms: a radical therapeutic approach? Nat. Rev. Drug Discov. 8, 579–591, doi:10.1038/nrd2803 (2009).
Ishaq, M., Evans, M. M. & Ostrikov, K. K. Effect of atmospheric gas plasmas on cancer cell signaling. Int. J. Cancer 134, 1517–1528, doi:10.1002/ijc.v134.7 (2014).
Hirst, A. M. et al. Low-temperature plasma treatment induces DNA damage leading to necrotic cell death in primary prostate epithelial cells. Br. J. Cancer 112, 1536–1545, doi:10.1038/bjc.2015.113 (2015).
Ikeda, J. et al. Anti-Cancer Effects of Nonequilibrium Atmospheric Pressure Plasma on Cancer-Initiating Cells in Human Endometrioid Adenocarcinoma Cells. Plasma Process. Polym. 12, 1370–1376, doi:10.1002/ppap.201500097 (2015).
Brulle, L. et al. Effects of a non thermal plasma treatment alone or in combination with gemcitabine in a MIA PaCa2-luc orthotopic pancreatic carcinoma model. PLoS One 7, e52653, doi:10.1371/journal.pone.0052653 (2012).
Hattori, N. et al. Effectiveness of plasma treatment on pancreatic cancer cells. Int. J. Oncol. 47, 1655–1662, doi:10.3892/ijo.2015.3149 (2015).
Utsumi, F. et al. Effect of indirect nonequilibrium atmospheric pressure plasma on anti-proliferative activity against chronic chemo-resistant ovarian cancer cells in vitro and in vivo. PLoS One 8, e81576, doi:10.1371/journal.pone.0081576 (2013).
Bundscherer, L. et al. Viability of human blood leucocytes compared with their respective cell lines after plasma treatment. Plasma Med. 3, 71–80, doi:10.1615/PlasmaMed.v3.i1-2 (2013).
Bekeschus, S. et al. Redox Stimulation of Human THP-1 Monocytes in Response to Cold Physical Plasma. Oxid. Med. Cell. Longev. 2016, 5910695–11, doi:10.1155/2016/5910695 (2016).
Wende, K., Reuter, S., von Woedtke, T., Weltmann, K. D. & Masur, K. Redox-Based Assay for Assessment of Biological Impact of Plasma Treatment. Plasma Process. Polym. 11, 655–663, doi:10.1002/ppap.201300172 (2014).
Winter, J. et al. Feed gas humidity: a vital parameter affecting a cold atmospheric-pressure plasma jet and plasma-treated human skin cells. J. Phys. D: Appl. Phys. 46, 295401, doi:10.1088/0022-3727/46/29/295401 (2013).
Winter, J. et al. Tracking plasma generated H2O2 from gas into liquid phase and revealing its dominant impact on human skin cells. J. Phys. D: Appl. Phys. 47, 285401, doi:10.1088/0022-3727/47/28/285401 (2014).
Golda, J. et al. Concepts and characteristics of the ‘COST Reference Microplasma Jet’. J. Phys. D: Appl. Phys. 49, 084003, doi:10.1088/0022-3727/49/8/084003 (2016).
Lackmann, J. W. et al. Photons and particles emitted from cold atmospheric-pressure plasma inactivate bacteria and biomolecules independently and synergistically. J R Soc Interface 10, 20130591–20130591, doi:10.1098/rsif.2013.0591 (2013).
Lackmann, J.-W. et al. Effects of the Effluent of a Microscale Atmospheric Pressure Plasma-jet Operated with He/O 2 Gas on Bovine Serum Albumin. Plasma Med. 3, 115–124, doi:10.1615/PlasmaMed.v3.i1-2 (2013).
Pendergrass, W., Wolf, N. & Poot, M. Efficacy of MitoTracker Green and CMXrosamine to measure changes in mitochondrial membrane potentials in living cells and tissues. Cytometry A 61, 162–169, doi:10.1002/cyto.a.20033 (2004).
Chiarugi, A. “Simple but not simpler”: toward a unified picture of energy requirements in cell death. The FASEB Journal 19, 1783–1788, doi:10.1096/fj.05-4200rev (2005).
Bekeschus, S. et al. Hydrogen peroxide: A central player in physical plasma-induced oxidative stress in human blood cells. Free Radic. Res. 48, 542–549, doi:10.3109/10715762.2014.892937 (2014).
Kirkman, H. N. & Gaetani, G. F. Catalase: a tetrameric enzyme with four tightly bound molecules of NADPH. Proc. Natl. Acad. Sci. USA 81, 4343–4347, doi:10.1073/pnas.81.14.4343 (1984).
Ida, T. et al. Reactive cysteine persulfides and S-polythiolation regulate oxidative stress and redox signaling. Proc. Natl. Acad. Sci. USA 111, 7606–7611, doi:10.1073/pnas.1321232111 (2014).
Sagristá, M. L., GarcÍa, A. F., De Madariaga, M. A. & Mora, M. Antioxidant and Pro-oxidant Effect of the Thiolic Compounds N -acetyl- l -cysteine and Glutathione against Free Radical-induced Lipid Peroxidation. Free Radic. Res. 36, 329–340, doi:10.1080/10715760290019354 (2009).
Setsukinai, K., Urano, Y., Kakinuma, K., Majima, H. J. & Nagano, T. Development of novel fluorescence probes that can reliably detect reactive oxygen species and distinguish specific species. J. Biol. Chem. 278, 3170–3175, doi:10.1074/jbc.M209264200 (2003).
Flemmig, J. et al. Fast and Specific Assessment of the Halogenating Peroxidase Activity in Leukocyte-enriched Blood Samples. J Vis Exp e54484–e54484, doi:10.3791/54484 (2016).
Pulli, B. et al. Measuring myeloperoxidase activity in biological samples. PLoS One 8, e67976, doi:10.1371/journal.pone.0067976 (2013).
Flemmig, J., Zschaler, J., Remmler, J. & Arnhold, J. The fluorescein-derived dye aminophenyl fluorescein is a suitable tool to detect hypobromous acid (HOBr)-producing activity in eosinophils. J. Biol. Chem. 287, 27913–27923, doi:10.1074/jbc.M112.364299 (2012).
Schlegel, J., Köritzer, J. & Boxhammer, V. Plasma in cancer treatment. Clin Plas Med 1, 2–7, doi:10.1016/j.cpme.2013.08.001 (2013).
Brown, N. S. & Bicknell, R. Hypoxia and oxidative stress in breast cancer. Oxidative stress: its effects on the growth, metastatic potential and response to therapy of breast cancer. Breast Cancer Res. 3, 323–327, doi:10.1186/bcr315 (2001).
Yan, D. et al. Toward understanding the selective anticancer capacity of cold atmospheric plasma–a model based on aquaporins (Review). Biointerphases 10, 040801, doi:10.1116/1.4938020 (2015).
de la Haba, C., Palacio, J. R., Martinez, P. & Morros, A. Effect of oxidative stress on plasma membrane fluidity of THP-1 induced macrophages. Biochim. BioPhys. Acta 1828, 357–364, doi:10.1016/j.bbamem.2012.08.013 (2013).
Coleman, M. L. et al. Membrane blebbing during apoptosis results from caspase-mediated activation of ROCK I. Nat. Cell Biol. 3, 339–345, doi:10.1038/35070009 (2001).
Aldo, P. B., Craveiro, V., Guller, S. & Mor, G. Effect of culture conditions on the phenotype of THP-1 monocyte cell line. Am. J. Reprod. Immunol. 70, 80–86, doi:10.1111/aji.2013.70.issue-1 (2013).
Schmidt, A. et al. Redox-regulation of activator protein 1 family members in blood cancer cell lines exposed to cold physical plasma-treated medium. Plasma Process. Polym. 13, 1179–1188, doi:10.1002/ppap.201600090 (2016).
Bekeschus, S. et al. Neutrophil extracellular trap formation is elicited in response to cold physical plasma. J. Leukoc. Biol. 100, 791–799, doi:10.1189/jlb.3A0415-165RR (2016).
Roebuck, K. A. Oxidant stress regulation of IL-8 and ICAM-1 gene expression: differential activation and binding of the transcription factors AP-1 and NF-kappaB (Review). Int. J. Mol. Med. 4, 223–230 (1999).
Bekeschus, S. et al. Distinct cytokine and chemokine patterns in chronic diabetic ulcers and acute wounds. Exp. Dermatol. 26, 145–147, doi:10.1111/exd.2017.26.issue-2 (2017).
Waugh, D. J. & Wilson, C. The interleukin-8 pathway in cancer. Clin. Cancer Res. 14, 6735–6741, doi:10.1158/1078-0432.CCR-07-4843 (2008).
Mukaida, N., Harada, A. & Matsushima, K. Interleukin-8 (IL-8) and monocyte chemotactic and activating factor (MCAF/MCP-1), chemokines essentially involved in inflammatory and immune reactions. Cytokine Growth Factor Rev. 9, 9–23, doi:10.1016/S1359-6101(97)00022-1 (1998).
Hendrickx, A. et al. Docking motif-guided mapping of the interactome of protein phosphatase-1. Chem. Biol. 16, 365–371, doi:10.1016/j.chembiol.2009.02.012 (2009).
Zhang, N., Kaur, R., Akhter, S. & Legerski, R. J. Cdc5L interacts with ATR and is required for the S-phase cell-cycle checkpoint. EMBO Rep 10, 1029–1035, doi:10.1038/embor.2009.122 (2009).
Willis, J., Patel, Y., Lentz, B. L. & Yan, S. APE2 is required for ATR-Chk1 checkpoint activation in response to oxidative stress. Proc. Natl. Acad. Sci. USA 110, 10592–10597, doi:10.1073/pnas.1301445110 (2013).
Martins, S., Eikvar, S., Furukawa, K. & Collas, P. HA95 and LAP2 beta mediate a novel chromatin-nuclear envelope interaction implicated in initiation of DNA replication. J. Cell Biol. 160, 177–188, doi:10.1083/jcb.200210026 (2003).
Hosokawa, M. et al. Cell cycle arrest and apoptosis induced by SART-1 gene transduction. Anticancer Res. 25, 1983–1990 (2005).
Faustino, R. S., Nelson, T. J., Terzic, A. & Perez-Terzic, C. Nuclear transport: target for therapy. Clin. Pharmacol. Ther. 81, 880–886, doi:10.1038/sj.clpt.6100141 (2007).
Dhar, S. K., Lynn, B. C., Daosukho, C. & St Clair, D. K. Identification of nucleophosmin as an NF-kappaB co-activator for the induction of the human SOD2 gene. J. Biol. Chem. 279, 28209–28219, doi:10.1074/jbc.M403553200 (2004).
Wang, B., Li, H., Lu, R. & Jin, Y. Function and molecular mechanism of a nucleolar protein LYAR in embryonic stem cells. Cell Res. 18, S7–S7, doi:10.1038/cr.2008.97 (2008).
Abreu-Martin, M. T., Chari, A., Palladino, A. A., Craft, N. A. & Sawyers, C. L. Mitogen-activated protein kinase kinase kinase 1 activates androgen receptor-dependent transcription and apoptosis in prostate cancer. Mol. Cell. Biol. 19, 5143–5154, doi:10.1128/MCB.19.7.5143 (1999).
Balasubramanyam, K., Swaminathan, V., Ranganathan, A. & Kundu, T. K. Small molecule modulators of histone acetyltransferase p300. J. Biol. Chem. 278, 19134–19140, doi:10.1074/jbc.M301580200 (2003).
Miyagi, Y. et al. Induction of cellular immune responses to tumor cells and peptides in colorectal cancer patients by vaccination with SART3 peptides. Clin. Cancer Res. 7, 3950–3962 (2001).
Yang, D. et al. Identification of a Gene Coding for a Protein Possessing Shared Tumor Epitopes Capable of Inducing HLA-A24-restricted Cytotoxic T Lymphocytes inCancer Patients. Cancer Res. 59, 4056–4063 (1999).
Sasatomi, T. et al. Expression of tumor rejection antigens in colorectal carcinomas. Cancer 94, 1636–1641, doi:10.1002/(ISSN)1097-0142 (2002).
Suefuji, Y. et al. Expression of SART3 antigen and induction of CTLs by SART3-derived peptides in breast cancer patients. Br. J. Cancer 84, 915–919, doi:10.1054/bjoc.2000.1690 (2001).
Koga, M. et al. Analysis of cellular localization of SART3 tumor antigen by a newly established monoclonal antibody: heterotopic expression of SART3 on the surface of B-lineage leukemic cells. Oncol. Rep. 11, 785–789 (2004).
Girard, P. M. et al. Synergistic Effect of H2O2 and NO2 in Cell Death Induced by Cold Atmospheric He Plasma. Sci. Rep. 6, 29098, doi:10.1038/srep29098 (2016).
Dobrynin, D., Arjunan, K., Fridman, A., Friedman, G. & Clyne, A. M. Direct and controllable nitric oxide delivery into biological media and living cells by a pin-to-hole spark discharge (PHD) plasma. J. Phys. D: Appl. Phys. 44, 075201, doi:10.1088/0022-3727/44/7/075201 (2011).
Uchiyama, H. et al. EPR-Spin Trapping and Flow Cytometric Studies of Free Radicals Generated Using Cold Atmospheric Argon Plasma and X-Ray Irradiation in Aqueous Solutions and Intracellular Milieu. PLoS One 10, e0136956, doi:10.1371/journal.pone.0136956 (2015).
Balzer, J. et al. Non-Thermal Dielectric Barrier Discharge (DBD) Effects on Proliferation and Differentiation of Human Fibroblasts Are Primary Mediated by Hydrogen Peroxide. PLoS One 10, e0144968, doi:10.1371/journal.pone.0144968 (2015).
Kaushik, N. et al. Responses of solid tumor cells in DMEM to reactive oxygen species generated by non-thermal plasma and chemically induced ROS systems. Sci. Rep. 5, 8587, doi:10.1038/srep08587 (2015).
Bekeschus, S., Iseni, S., Reuter, S., Masur, K. & Weltmann, K.-D. Nitrogen Shielding of an Argon Plasma Jet and Its Effects on Human Immune Cells. IEEE Trans. Plasma Sci. 43, 776–781, doi:10.1109/TPS.2015.2393379 (2015).
Priya Arjunan, K. & Morss Clyne, A. Hydroxyl Radical and Hydrogen Peroxide are Primarily Responsible for Dielectric Barrier Discharge Plasma-Induced Angiogenesis. Plasma Process. Polym. 8, 1154–1164, doi:10.1002/ppap.v8.12 (2011).
Aruoma, O. I., Halliwell, B., Hoey, B. M. & Butler, J. The antioxidant action of N-acetylcysteine: its reaction with hydrogen peroxide, hydroxyl radical, superoxide, and hypochlorous acid. Free Radic. Biol. Med. 6, 593–597, doi:10.1016/0891-5849(89)90066-X (1989).
Hasegawa, K. & Neta, P. Rate constants and mechanisms of reaction of chloride (Cl2-) radicals. The Journal of Physical Chemistry 82, 854–857, doi:10.1021/j100497a003 (1978).
Winterbourn, C. C. & Kettle, A. J. Biomarkers of myeloperoxidase-derived hypochlorous acid. Free Radic. Biol. Med. 29, 403–409, doi:10.1016/S0891-5849(00)00204-5 (2000).
Gomes, A., Fernandes, E. & Lima, J. L. Fluorescence probes used for detection of reactive oxygen species. J. Biochem. BioPhys. Methods 65, 45–80, doi:10.1016/j.jbbm.2005.10.003 (2005).
Zunino, S. J., Ducore, J. M. & Storms, D. H. Parthenolide induces significant apoptosis and production of reactive oxygen species in high-risk pre-B leukemia cells. Cancer Lett. 254, 119–127, doi:10.1016/j.canlet.2007.03.002 (2007).
Thiyagarajan, M., Anderson, H. & Gonzales, X. F. Induction of apoptosis in human myeloid leukemia cells by remote exposure of resistive barrier cold plasma. Biotechnol. Bioeng. 111, 565–574, doi:10.1002/bit.25114 (2014).
Kettle, A. J. & Winterbourn, C. C. The influence of superoxide on the production of hypochlorous acid by human neutrophils. Free Radic. Res. Commun. 12-13(Pt 1), 47–52, doi:10.3109/10715769109145766 (1991).
Klebanoff, S. J., Kettle, A. J., Rosen, H., Winterbourn, C. C. & Nauseef, W. M. Myeloperoxidase: a front-line defender against phagocytosed microorganisms. J. Leukoc. Biol. 93, 185–198, doi:10.1189/jlb.0712349 (2013).
Parker, H., Albrett, A. M., Kettle, A. J. & Winterbourn, C. C. Myeloperoxidase associated with neutrophil extracellular traps is active and mediates bacterial killing in the presence of hydrogen peroxide. J. Leukoc. Biol. 91, 369–376, doi:10.1189/jlb.0711387 (2012).
Hawkins, C. L. & Davies, M. J. Inactivation of protease inhibitors and lysozyme by hypochlorous acid: role of side-chain oxidation and protein unfolding in loss of biological function. Chem. Res. Toxicol. 18, 1600–1610, doi:10.1021/tx050207b (2005).
Stacey, M. M., Vissers, M. C. & Winterbourn, C. C. Oxidation of 2-cys peroxiredoxins in human endothelial cells by hydrogen peroxide, hypochlorous acid, and chloramines. Antioxid. Redox Signal. 17, 411–421, doi:10.1089/ars.2011.4348 (2012).
Del Giudice, A., Dicko, C., Galantini, L. & Pavel, N. V. Structural Response of Human Serum Albumin to Oxidation: Biological Buffer to Local Formation of Hypochlorite. J. Phys. Chem. B 120, 12261–12271, doi:10.1021/acs.jpcb.6b08601 (2016).
Ronsein, G. E., Winterbourn, C. C., Di Mascio, P. & Kettle, A. J. Cross-linking methionine and amine residues with reactive halogen species. Free Radic. Biol. Med. 70, 278–287, doi:10.1016/j.freeradbiomed.2014.01.023 (2014).
Wende, K. et al. Identification of the biologically active liquid chemistry induced by a nonthermal atmospheric pressure plasma jet. Biointerphases 10, 029518, doi:10.1116/1.4919710 (2015).
Ellerweg, D., Benedikt, J., von Keudell, A., Knake, N. & Schulz-von der Gathen, V. Characterization of the effluent of a He/O2microscale atmospheric pressure plasma jet by quantitative molecular beam mass spectrometry. New J. Phys. 12, 013021 (2010).
Benedikt, J. et al. Absolute OH and O radical densities in effluent of a He/H2O micro-scaled atmospheric pressure plasma jet. Plasma Sources Sci. T. 25, 045013, doi:10.1088/0963-0252/25/4/045013 (2016).
Hefny, M. M., Pattyn, C., Lukes, P. & Benedikt, J. Atmospheric plasma generates oxygen atoms as oxidizing species in aqueous solutions. J. Phys. D: Appl. Phys. 49, 404002, doi:10.1088/0022-3727/49/40/404002 (2016).
Sousa, J. S. et al. Cold atmospheric pressure plasma jets as sources of singlet delta oxygen for biomedical applications. J. Appl. Phys. 109, 123302, doi:10.1063/1.3601347 (2011).
Wilkinson, F. & Brummer, J. G. Rate constants for the decay and reactions of the lowest electronically excited singlet state of molecular oxygen in solution. J. Phys. Chem. Ref. Data 10, 809–999, doi:10.1063/1.555655 (1981).
Held, A. M., Halko, D. J. & Hurst, J. K. Mechanisms of Chlorine Oxidation of Hydrogen-Peroxide. JACS 100, 5732–5740, doi:10.1021/ja00486a025 (1978).
Foley, S. et al. Singlet oxygen quenching and the redox properties of hydroxycinnamic acids. Free Radic. Biol. Med. 26, 1202–1208, doi:10.1016/S0891-5849(98)00313-X (1999).
Jablonowski, H. and von Woedtke, Th. Research on plasma medicine-relevant plasma–liquid interaction: What happened in the past five years? Clin. Plas. Med. 3, 42–52, doi:10.1016/j.cpme.2015.11.003 (2015).
Chen, C. et al. A Model of Plasma-Biofilm and Plasma-Tissue Interactions at Ambient Pressure. Plasma Chem. Plasma Process. 34, 403–441, doi:10.1007/s11090-014-9545-1 (2014).
Poot, M. et al. Analysis of mitochondrial morphology and function with novel fixable fluorescent stains. J. Histochem. Cytochem. 44, 1363–1372, doi:10.1177/44.12.8985128 (1996).
Wende, K. et al. Proteomic Tools to Characterize Non-Thermal Plasma Effects in Eukaryotic Cells. Plasma Med. 3, 81–95, doi:10.1615/PlasmaMed.v3.i1-2 (2013).
Acknowledgements
Funding was received by the German Federal Ministry of Education and Research (grant numbers 03Z22DN11 and 13N13960); by the Ministry of Education, Science, and Culture of the State of Mecklenburg-Western Pomerania (grant number AU 15 001); and by German Science Foundation (DFG, grant number BE 4349/2-1 as part of project package PAK728).
Author information
Authors and Affiliations
Contributions
S.B., K.W., M.M.H., and J.B. designed the study; S.B., M.M.H., and K.R. carried out the experiments; S.B., M.M.H., and K.W. did the data analysis; all authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bekeschus, S., Wende, K., Hefny, M.M. et al. Oxygen atoms are critical in rendering THP-1 leukaemia cells susceptible to cold physical plasma-induced apoptosis. Sci Rep 7, 2791 (2017). https://doi.org/10.1038/s41598-017-03131-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-03131-y
- Springer Nature Limited
This article is cited by
-
Helium Plasma Effects on Polymer Surfaces: from Plasma Parameters and Surface Properties towards Bioengineering Applications
Chinese Journal of Polymer Science (2024)
-
Characterization of Non-Thermal Dielectric Barrier Discharges for Plasma Medicine: From Plastic Well Plates to Skin Surfaces
Plasma Chemistry and Plasma Processing (2023)
-
Cold atmospheric plasma: a sustainable approach to inactivating viruses, bacteria, and protozoa with remediation of organic pollutants in river water and wastewater
Environmental Science and Pollution Research (2023)
-
Removal of chemical and microbial water pollutants by cold plasma combined with Ag/TiO2-rGO nanoparticles
Scientific Reports (2022)
-
Tumor cytotoxicity and immunogenicity of a novel V-jet neon plasma source compared to the kINPen
Scientific Reports (2021)