
Abstract
Perception of environmental and habitat cues is of significance for insect survival and reproduction. Odor detection in insects is mediated by a number of proteins in antennae such as odorant receptors (ORs), ionotropic receptors (IRs), odorant binding proteins (OBPs), chemosensory proteins (CSPs), sensory neuron membrane proteins (SNMPs) and odorant degrading enzymes. In this study, we sequenced and assembled the adult male and female antennal transcriptomes of a destructive agricultural pest, the diamondback moth Plutella xyllostella. In these transcriptomes, we identified transcripts belonging to 6 chemoreception gene families related to ordor detection, including 54 ORs, 16 IRs, 7 gustatory receptors (GRs), 15 CSPs, 24 OBPs and 2 SNMPs. Semi-quantitative reverse transcription PCR analysis of expression patterns indicated that some of these ORs and IRs have clear sex-biased and tissue-specific expression patterns. Our results lay the foundation for future characterization of the functions of these P. xyllostella chemosensory receptors at the molecular level and development of novel semiochemicals for integrated control of this agricultural pest.
Similar content being viewed by others


Introduction
Olfaction plays a pivotal role in intra- and inter-specific interactions by directing insects towards food or prey, mating partners, oviposition sites, and away predators as well as toxic compounds1. The specialized organ for olfaction in insects is the antenna, on which hair-like, multi-pore sensilla are situated and peripheral olfactory signaling events occur. Olfactory receptor neurons (ORNs) and their auxiliary structures are located at the roots of the antennae2, and the entire olfactory system is dependent to a great extent on receptors expressed at the peripheral ORNs. Starting with perception of semiochemicals and ultimately ending with the translation of olfactory signals into behavior, the entire process requires orchestration of the insect’s sophisticated olfactory system at various levels. Several types of olfactory proteins are believed to participate in the selective detection and, once they have conveyed information, the rapid inactivation of trace amount of odorants, i.e. odorant receptors (ORs), ionotropic receptors (IRs), gustatory receptors (GRs), odorant binding proteins (OBPs), chemosensory proteins (CSPs) and sensory neuron membrane proteins (SNMPs)3.
Insect ORs are seven-transmembrane domain proteins with a reversed topology compared to the G-protein coupled ORs in vertebrates4,5. ORs play a central role in converting semiochemicals into electrical signal, functioning as a heterodimer with a divergent, conventional ORx and a highly conserved noncanonical OR co-receptor Orco in fruit fly, OR2 in moths and OR7 in mosquitoes3. The OR genes are expressed in the olfactory neurons housed within the olfactory sensilla (found mainly on the antenna)6.
GRs are also seven-transmembrane domain proteins, but they are more ancient than ORs. GR genes are expressed in the gustatory neurons housed within the gustatory sensilla (found on the labia, maxillary palps, antennae, legs and genitalia)7. GRs can respond to tastants such as sugars, bitter substances, CO2 and some contact pheromones8,9,10,11.
IRs belong to the ionotropic glutamate receptor (iGluR)-like protein family and can be activated by small molecules like acetates and amine-like volatile compounds12,13. It has been proven that IRs are involved in chemosensation14,15 and other functions, i.e. regulation of the circadian clock in Drosophila melanogaster 16 and induction of physical defense in Daphnia pulex 17. IRs usually contain three transmembrane domains (TMDs), a bipartite ligand-binding domain with two lobes and one ion channel, and have been proposed to act as dimmers or trimers of subunits coexpressed in the same neuron12. However, they aren’t expressed in chemosensory neurons that express ORs or Orco14.
OBPs are the liaisons between external cues and ORs18, and they selectively bind hydrophobic odorant chemicals and transport them to the surface of the dendrites of ORNs19,20,21. OBPs also function in the recognition of specific odors through activation of the ORx/Orco complex20. Another class of odorant binding proteins, CSPs, are small soluble proteins expressed predominantly in the sensilum lymph as well as in non-olfactory tissues. It is clear that CSPs bind odorant or pheromone compounds22,23,24, but their olfactory mechanisms areas yet poorly studied.
SNMPs are insect membrane proteins that are known to associate with pheromone sensitive ORNs in Lepidoptera and Diptera25. There are two types of SNMPs, SNMP1 and SNMP225. In moth, the subtype SNMP1 is coexpressed with pheromone receptors (PRs) in pheromone-responsive neurons25, whereas the subtype SNMP2 is confined to sensilla support cells25,26,27,28.
The diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae), is a destructive insect pest distributed worldwide that can cause considerable damage in cruciferous crops. It is estimated that the total loss caused by P. xylostella is about US$4-5 billion annually29. Although a bioinformatics analysis of the whole-genome sequence has explained the evolutionary success of P. xylostella with regard to its expansion in gene families associated with the perception and detoxification of plant defense compounds/insecticides at the genetic and molecular levels30, the peripheral olfactory mechanisms that contribute to the fitness of this insect pest remain poorly understood. Identification of genes expressed in the antennae will supply baseline information to understand their likely function in odorant perception in P. xylostella and insects adaptation to various host plants.
In the present study, we sequenced and analyzed the antennal transcriptome of P. xylostella adults using second-generation high-throughput Illumina RNA sequencing (RNA-seq). The purpose of our study was to identify olfaction-related genes which might be targets as a part of pest control strategies of this insect pest species that devastates cruciferous vegetables. We identified 118 candidate chemosensory genes encoding 54 ORs, 16 IRs, 7 GRs, 15 CSPs, 24 OBPs and 2 SNMPs. The sex-biased and tissue-specific expression patterns of 54 ORs and 16 IRs was also determined by semi-quantitative reverse transcription PCR. We reported the protein sequences of these chemosensory genes in Supplementary Dataset File.
Results
Sequencing and unigene assembly
By using Hiseq. 2000 sequencing approach, a total of 60,041,232 and 59,753,272 raw reads were obtained from the P. xylostella female and male antennae samples, respectively. After removing low quality and adaptor reads, female and male antennae yielded 54,430,716 and 54,059,300 clean reads and 4,898,764,440 nt and 4,865,337,000 nt clean nucleotides, respectively. After initial assembly, 124,488(mean length 278 nt) and 132,190 contigs (mean length 268 nt) were obtained from the female and male antennae libraries, respectively. Next, 62,278 female (mean length 555 nt) and 63,928 male unigenes (mean length 531 nt) were generated after contig connecting. These two unigene sets were then pooled together for further clustering, which yielded a final set of 59,844unigenesconsisting of 18,570 distinct clusters and 41,274 distinct singletons. The mean length of these unigenes was 660 nt, and N50 was 979 nt (Table 1).
Identification of candidate chemosensory receptors: ORs and GRs
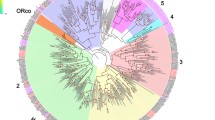
All the unigenes were searched by blastx against nr database and further by tblastn using 63 ORs from B. mori as queries, 54 candidate OR genes were identified (Table 2). Of these, 23 were predicted to have full-length open reading frames (ORFs). The length of these 23 OR genes ranges from 376 to 473 amino acid residues, and the encoded proteins are estimated to have 5–7 TMDs, which is characteristic of typical insect ORs. The remaining 31 OR genes code for at least 163 amino acids and are predicted to have more than one TMD. A phylogenetic analysis was then performed using our candidate ORs and the ORs from other Lepidopteran insects including H. armigera, H. virescens and B. mori (Fig. 1).

Phylogenetic tree of candidate Lepidopteran ORs, including the PR and Orco clades. Pxyl: P. xylostella (red), Harm: Helicoverpa armigera (black), Hvir: Heliothis virescens (green), Bmor: Bombyx mori (blue). The clade shaded in blue indicates the Orco clade. The clade shaded in red indicates the PBP clade. The bootstrap value for phylogenetic tree construction is 1000.
The OR co-receptor gene was easily identified because of extremely high conservation among species compared to other chemosensory receptors. Similar to other insect ORs, most P. xylostella (Pxyl) ORs are highly divergent and share low similarity with other Lepidopteran insect ORs, including ORs from H. armigera, H. virescens and B. mori. However, nine PxylORs had 33%~100% identity to previously characterized PRs from P. xylostella and B. mori. They formed a single subgroup in a phylogenetic tree of Lepidopteran ORs (Fig. 1). Seven of these nine PxylORs (PxylOR1 andPxylOR3-8) were predicted to have full-length ORFs. Two short sequences (PxylOR41and PxylOR45) were also clustered in the PR branch. PxylOR41 has high similarity to PxylOR4, and PxylOR45 has relatively high similarity to BmorOR6. 12 of the remaining PxylORs were clustered with their Lepidopteran orthologous genes in the phylogenetic tree. But most PxylORs appeared to be distantly related to the known insect ORs (Fig. 1). We named the Orco unigene PxylOR2 and the 7full-length candidate PR unigenes PxylOR1 and PxylOR3-PxylOR8. The other 46 OR unigenes were ranked in order of decreasing ORF length and named PxylOR9-PxylOR54. We also identified 7 candidate GRs and named them as PxylGR1-PxylGR7.
Identification of candidate IRs
IR sequences in the P. xylostella antennal transcriptome were identified based on similarity to known IRs of Lepidopteran insects, B. mori, C. pomonella, H. armigera, H. virescens and S. littoralis. Sixteen candidate IRs were identified by bioinformatic analysis, and five unigenes were predicted to have a full-length ORFs. The insect IRs typically have three TMDs. Of the 16 candidates IRs, 15 are predicted to have at least one TMD (Table 3). Twelve of the 16 putative IRs are at least 48% identical to the corresponding Lepidopteran orthologous IRs in S. littoralis and C. pomonella. The remaining four unigenes have relatively low similarity to other insect IRs: CL2177.Contig2 has 35% identity with IR1 of S. littoralis, unigene13888 has 31% identity with IR75 of C. pomonella, CL4692.Contig1 has only 25% identity with IR60a of D. melanogaster, and CL5979.Contig2 has only 24% identity with IR7c of D. melanogaster (Table 3). Phylogenetic analyses suggested that the prediction of IRs was credible. In a neighbor-joining tree of insect IRs, all candidate PxylIRs were clustered in a separate clade with their Lepidopteran orthologs (Fig. 2). All of these 16 candidate IR unigenes were named based on their homology to known IRs. For example, the IR Unigene 19385 has 55% similarity withIR75q2 and CL1791. Contig1 had 64% similarity to IR75q2. So, we named Unigene 19835 PxylIR75q2.2.

Phylogenetic tree of candidate IRs from Pluttela xylostella and other insects. Pxyl: P. xylostella (red), Harm: Helicoverpa armigera (black), Hvir: Heliothis virescens (green), Bmor: Bombyx mori (blue), Slit: Spodoptera littoralis (purple), Cpom: Cydia pomonella (cyan).The clade shaded in red indicates the IR8a/IR25a clade. The bootstrap value for phylogenetic tree construction is 1000.
Identification of putative OBPs
We identified 24 unigenes encoding OBPs from the antennal transcriptome of P. xylostella, including 3pheromone binding proteins (PBPs) and 3 general odorant binding proteins (GOBPs) (Table 4). Twenty-two of these 24 unigenes were predicted to have signal peptides, and 19 have full length ORFs. Signal peptide sequences were not detected in the remaining two putative OBPs due to incomplete N-terminal sequences. All 24 putative OBPs had high similarity to known Lepidopteran OBPs. The PBP and GOBP sequences were clustered in a separate clade in the OBP neighbor-joining tree (Fig. 3). Three candidate OBPs were classified into a PBP subgroup in the phylogenetic tree. They share 66%~100% similarity with previously characterized Lepidopteran PBPs and thus were named PBPs. We also found two GOBPs in the antennal transcriptome of P. xylostella and named them PxylGOBP1 and GOBP2. A new GOBP (PxylGOBP1.2) was identified that has 77% identity with PxylGOBP1. It was clustered in the GOBP clade and distinguished from other OBPs in the phylogenetic tree. The other 18 candidate OBPs are obviously distinct from the PBP and GOBP clades and have relatively lower similarity to OBPs from other Lepidopteran insects. Most candidate OBP sequences, such as PxylOBP2, PxylOBP3, and PxylOBP7, are closely clustered with at least one Lepidopteran ortholog, in congruence with the blastx results. Some candidate OBP sequences such as PxylOBP6, PxylOBP9, PxylOBP11 and PxylOBP17 are not clustered with OBPs from other Lepidopteran insects (Fig. 3). A possible reason may be that the orthologs of these PxylOBPs have not been identified in other Lepidopteran insects.

Phylogenetic tree of candidate Lepidopteran OBPs, including the GOBP and PBP clades. Pxyl: Pluttela xylostella (red), Harm: Helicoverpa armigera (black), Hvir: Heliothis virescens (green), Bmor: Bombyx mori (blue). The clade shaded in blue indicates the PBP clade. The clade shaded in red indicates the GOBP clade. The bootstrap value for phylogenetic tree construction is 1000.
Identification of candidate CSPs
Bioinformatic analysis led to the identification of 15 different sequences encoding candidate CSPs (Table 5). All 15 unigenes were predicted to have signal peptides and 14 have a full length ORFs. Four candidate PxylCSPs (PxylCSP1-4) match the previously identified P. xylostella CSP sequences31. The other 11 candidate CSP sequences have at least 35% identity with known CSPs from other insects, and we named them according to the length of the coding region in descending order. In a neighbor-joining tree, all 15 sequences form a cluster with Lepidopteran orthologous genes (Fig. 4).

Phylogenetic tree of candidate Lepidopteran CSPs. Pxyl: Pluttela xylostella (red), Harm: Heliocoverpa armigera (black), Hvir: Hethiothis virescens (green), Bmor: Bombyx mori (blue). The bootstrap value for phylogenetic tree construction is 1000.
Identification of candidate SNMPs
SNMPs were first identified in pheromone-sensitive neurons of Lepidoptera31 and are thought to function in pheromone detection32. Two kinds of SNMPs (SNMP1 and SNMP2) have been identified in insects and transcripts corresponding to both were found in the P. xylostella transcriptome. The sequence of CL2414Contig2 is identical to the PxylSNMP1sequence published in Genbank. CL242.Contig4 has 70% identity with SNMP2 of O. furnacalis, and we annotated this sequence as P. xylostella SNMP2 (Table 6).
Tissue- and sex-specific expression of candidate OR and IR genes in P. xylostella
To investigate the expression profile of PxylORs and PxylIRs between sexes and tissues, we determined the expression patterns of the 54 candidate ORs and 16 candidate IRs genes in the antennae and legs of male and female adult P. xylostella by semi-quantitative reverse transcription PCR (RT-PCR) (Fig. 5). As shown in Fig. 5, all of these 54 ORs were expressed in the antennae. PxylOR2 was expressed in male and female antennae and legs. The expression of the nine candidate PRs was observed only in antennae but not in legs. And of which, seven candidate PRs (PxylOR1, 3, 4, 5, 6, 7 and 41) had male-biased or male-specific expression patterns. Different from the other lepidopteran ORs, PxylOR8, was only expressed in female antennae. PxylOR45 was expressed in both male and female at a similar level. In other 44 general ORs PxylOR54 expression was much higher in female than in male antenna and the remaining 43 ORs were expressed in both male and female antennae at a similar level. In contrast to ORs, the expression of all IRs did not differ significantly between males and females. All of these 16 PxylIRs were expressed in the male and female antennae, but PxylIR7d.3 and PxylIR25a were also expressed in legs.

Tissue- and sex-specific expression patterns of candidate PxylORs and PxylIRs. M: male antennae, F: female antennae, L: legs. PxylRPS3 is the reference.
Discussion
In the present study, we profiled the antennal transcriptome of P. xylostella adults by RNA-seq technology and annotated 118 putative olfactory genes, including 54 putative ORs, 24 OBPs, 16 IRs, 15 CSPs, 7 GRs, and 2 SNMPs. Chemosensory genes have been identified in other Lepidopteran insects; 134 putative chemosensory unigenes were identified in the antennae of H. armigera, including 60 ORs, 34 OBPs, 19 IRs,18CSPs, 1 GR and 2 SNMPs, and 131 putative chemosensory unigenes were identified in H. assulta antennae, including 64 ORs, 19 IRs, 29 OBPs, 17 CSPs, and 2 SNMPs33. Our results are comparable with those from H. armigera and H. assulta in the number of genes identified. The identification of chemosensory genes from antennal transcriptomes was also reported for the moth M. sexta (91 genes, including 48 ORs, 18 OBPs, 21 CSPs and 4 IRs)34 and B. mori (138 genes, including 71 ORs, 20 OBPs, 16 CSPs and 31 IRs)35,36,37,38 and many other insect pests.
Insects utilize three groups of chemosensory receptors, ORs, IRs and GRs, to perform a variety of essential behaviors such as foraging, mating and oviposition. ORs are the centerpiece of peripheral olfactory reception and determine the sensitivity and specificity of odorant reception3. Due to the availability of insect genome databases and progress in sequencing technology, increasing numbers of OR genes have been identified from many Lepidopteran species. To date, 68, 64, 70 ORs have been identified in the genome databases of B. mori 38, Danaus plexippus 39 and Helioconius Melpomene 40, respectively. Recently, by using next-generation sequencing technology the antennal transcriptome of M. sexta was profiled, and 48 OR genes were identified34,41. In this study, we identified 54 ORs in the antennal transcriptome of adult P. xylostella. The number of ORs identified in this paper is less than that identified by You et al.30 in the genome database of P. xylostella. We might have missed some development-related OR genes because we only identified chemosensory genes in the adult antennae. Typical insect ORs are characterized by seven TMDs. We found less than seven TMDs in PxylORs, which is also observed in other Lepidopteran insects33,42,43. This is probably caused by the limited power of the software used for TMDs finding.
All of the PxylORs identified in the antennal transcriptome are highly divergent and share low similarity with other Lepidopteran insect ORs. A study showed that the common ancestor of Lepidopterans had fewer OR genes but that there were multiple gene gains and few gene losses during the evolution of Lepidoptera. This phenomenon of gene family expansion is suggested to be associated with the adaption of Lepidopteran species to host plants44. We also identified 9 (PxylOR1, PxylOR3-8, PxylOR41 and PxylOR45) candidate PRs based on their similarity to previously characterized PRs. The antennal expression pattern of PoxylPRs is consistent with that of PRs in H. armigera 42 and S. littoralis 45. Among these 9 candidate PRs, 7 showed male-biased expression, and PxylOR5 was only expressed in male antennae. In contrast, PoxylOR8 was only expressed in female antennae. Sex and tissue-specific expression of chemosensory genes is very common among Lepidoperan pests. It was found in H. assulta 33 and H. armigera 42 that some of their antennal OR genes showed sex-biased expression pattern. The male-specific expression of PxylOR5 probably plays a role in locating females, while the female-specific expression PxylOR8 likely also has ecological significance, i.e. optimization of pheromone production and spatial dispersion of females among host plants46,47 and selection of oviposition sites.
We identified one Orco unigene, named PxylOR2, which has high similarity to HarmOR2, BmorOR2 and HvirOR2. Orco is highly conserved among all insect species3 and carries out similar functions in different insects48 by forming a ligand-gated ion channel49. Orco probably functions as a chaperone and forms a dimer with the other ORs in P. xylostella.
GRs can respond to tastants such as sugars, bitter substances, CO2 and some contact pheromones50. Thus, GRs play very important roles in food selection and feeding behaviors in insects. The first insect GRs were identified in the fruit fly, D. melanogaster 51. The number of Lepidopteran GRs varies greatly; there is one GR in Cydia pomonella 52 and H. armigera 42, 2 in M. sexta 34, 3 in Heliothis virescens 53 and 5 in Spodoptera littoralis 45,54. In the antennal transcriptome of adult P. xylostella we identified 7 GRs, which is more than those in the Lepidopteran insects mentioned above, but far less than the number found in the silkworm B. mori (65 GRs)55 and the oriental tobacco budworm H. assulta (18 GRs)56. GRs are mainly expressed in gustatory organs such as the proboscis and maxillary palps, rather than in antennae8. This is a possible reason why we identified only 7GRs in P. xylostella. Two GR genes, GR21a and GR63a have been proved to be putative CO2 receptors in the antennae of the fruit fly57,58. And in mosquitos, 3 putative CO2 receptor genes (GR22, 23 and 24) have been identified in the maxillary palps of different species59,60,61. The PxylGR1 was closely related to the GR22 in mosquito and GR21a in the fruit fly and predicted to be a candidate CO2 receptor.
IRs belong to an ancient chemosensory receptor family, and two subfamilies of IRs have been identified recently, i.e. the conserved ‘antennal IRs’ and the species-specific ‘divergent IRs’62. The first IR was identified in the coeloconic sensilla of Drosophila 14 and most Drosophila IRs have clear orthologs within the genus of Lepidoptera34,42,63. IRs are ligand-gated ion channels that mediate chemical communication between neurons14. In this study, we identified 16 IRs in the antennal transcriptome of P. xylostella and named them based on homologous sequences from other insects. Similar numbers of IRs have been identified from other Lepidopteran insects: 19 IRs were identified in the antennal transcriptomes of H. armigera and H. assult 33, 15 IRs in C. pomonella 52, 20 IRs in Chio suppresalis 43, and 12 IRs in S. litoralis 36. All of these IRs are expressed in antennae, but PxylIR7d.3 and PxylIR25a are also expressed in legs, which is different from the expression patterns of these genes in H. assulta 33. Coincidently, HarmIR25a, HarmIR75d, HarmIR75p and HarmIR76p are also expressed in the cotton bollworm legs42. The function of leg-expressed IRs remains unknown and deserves in-depth investigation.
OBPs are believed to be directly involved in the activation of the ORx/Orco complex in the recognition of specific odors20. A total of 24 OBPs were identified in the antennal transcriptome of P. xylostella, including three GOBPs and three PBPs. The number of OBPs identified in the present study was comparable to those identified in transcriptomic analyses of H. armigera (34) and H. assulta (29)33, S. litura (21)64, S. littoralis (26)54, but fewer than those identified in B. mori (44)37. OBPs showed lineage-specific expansion and diversification; therefore, it is not surprising that there are some differences, or even big differences, in the number of OBPs. Previous studies have also shown that some insect OBPs and CSPs are expressed exclusively in non-antennae tissues or in larvae65. Therefore, different sampling and sequencing strategies may lead to different results. In a previous study, two GOBPs, GOBP1 and 2, were identified in P. xylostella antennae66. GOBPs were also found in the antennae of C. pomonella 67 and S. litura 68. The antennal P. xylostella GOBPs identified in this study have ecological significance, e.g. guiding P. xylstella to find better food69. The antennal S. litura GOBP1 can bind to plant odorants, while S. litura GOBP2 can bind to aldehyde-sex compounds and analogs68.
CSPs are a class of small soluble proteins expressed highly in the chemosensilla lymph70 and show high binding activity to odorants and pheromones71. We identified 15CSPs genes in the present study. The number of CSPs identified from P. xylostella was comparable to the number in B. mori (18)72, H. armigera (18) and H. assulta (17)33 and S. litura (18)64, but fewer than the number in M. sexta (21)34, Sesamia inferens (24)63 and S. littoralis (31) 45. Because CSPs are also expressed in tissues other than antennae73,74 and may participate in other physiological processes, it is possible that we have missed some CSPs in our antennal transcriptome analysis.
SNMPs are two-transmembrane domain proteins that share very high homology to members of the mammalian CD36 receptor family, which are thought to function in pheromone detection of Lepidopteran and Dipteran insects31. Two subtypes of SNMPs (SNMP1 and SNPM2) have been frequently identified in most insects, e.g. Helicoverpa armigera 33,42, Cnaphalocrocis medinalis 27, S. exigua75, S. litura 28, C. suppressalis 43, H. assulta 33, and in this study, P. xylostella. The expression of antennal SNMPs in P. xylostella suggests their role in pheromone detection, similar to what has been reported in D. melanogaster 32,76.
Conclusions
In summary, we identified 118 candidate olfactory genes that may function in odorant perception in the diamondback moth, P. xylostella by assembling and annotating transcriptomic sequence data. We carried out a comparative phylogenetic analysis to predict gene functions and examined the transcriptome patterns of the P. xylostella OR and IR genes. Genes with sex-biased and tissue-specific expression patterns, especially PxylOR5 and PxylOR8, are potential targets for environmentally-friendly management of this destructive insect pest. Our results lay the foundation for functional analysis of these receptors in both neurobiological and evolutionary studies.
Materials and Methods
Insect rearing
The laboratory-maintained P. xylostella was reared in the Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing, China. The larvae and adults were fed on Chinese cabbage and kept in cages at 27 ± 1 °C under a 16: 8 (L: D) photoperiod and 65 ± 5% relative humidity. Male and female larvae were distinguished at the last instar and placed in separate cages. Antennae of female or male adults were dissected at 1–3 days after adult emergence, immediately frozen in liquid nitrogen, and then stored at −70 °C until use.
Total RNA extraction
The frozen antennae were transferred to a liquid nitrogen-cooled mortar and ground with a pestle. One mL of TRIzol reagent was pipetted to the homogenate (Invitrogen, Carlsbad, CA, USA) and total RNA was extracted following the manufacturer’s instructions. Total RNA was resuspended in RNAse-free H2O, and RNA quantity was determined with a Nanodrop ND-2000 spectrophotometer (NanoDrop products, Wilmington, DE, USA). RNA integrity was assessed using an Agilent 2100 BioAnalyzer (Agilent Technologies, Englewood, CO, USA).
cDNA Library construction and Illumina sequencing
Tenμg of total RNA, extracted from approximately 2000 antennae of 1–3 day old adult male or female moths. The cDNA library for each sample was prepared using the NEBNext® mRNA Library Prep Reagent Set for Illumina (NEB, Ipswich, MA, USA) following the manufacturer’s instructions. Poly-A RNA for each sample was fragmented in fragmentation buffer to a length of 200 nt–700 nt. Random hexamers were used to generate first-strand cDNA, and second-strand cDNA was synthesized using RNaseH and DNA polymerase I. The double-strand cDNA (ds cDNA) samples were purified with the QIAquick PCR Purification Kit (Qiagen, Hilden, Germany) and eluted with EB buffer. The short fragments were treated with T4 DNA Polymerase and T4 Polynucleotide Kinase for end-repair and dA-tailing, then sequencing adaptors with barcodes were ligated to the dA tail of ds cDNA using T4 DNA ligase. To select insert length, ds cDNA samples were separated by agarose gel electrophoresis and bands of approximately 200 bp were excised and purified with the QIAquick Gel Extraction Kit (Qiagen, Hilden, Germany). Paired-end sequencing of the library was performed on the Illumina HiSeq™ 2000 platform (Illumina, San Diego, CA, USA) at the Beijing Genome Institute (Shenzhen, China). The read length of each end was 90 bp. The male and female libraries were sequenced in one lane, and raw reads were then sorted by barcode sequence.
Unigene generation
Raw reads were pre-processed to remove low quality reads and reads containing adapter sequences and poly-A/T tails. The publicly available program Trinity was used to perform de novo assembly of clean reads to generate a set of transcripts77. The Trinity outputs were then clustered by TGICL (TGI Clustering tools)78. The final unigene dataset consists of uniformly clustered sequences and singletons.
Gene identification and functional annotation
Unigene sequences were first searched against protein databases like nr, Swiss-Prot, KEGG and COG, using blastx with an e-value cut-off of 1e−5 79. To identify more OR genes, 63ORs from B. mori were used as queries in tblastn searches of P. xylostella antennal unigenes. Unigene ESTs were predicted using ESTScan80. Signal peptides in the protein sequences were predicted using SignaIP 4.081. The TMDs of annotated genes were predicted using TMHMM Server Version2.0 (http://www.cbs.dtu.dk/services/TMHMM).
Phylogenetic analyses
Phylogenetic trees were constructed based on the amino sequences of the candidate olfaction genes and genes from the collected data sets. The OR datasets contained OR sequences identified from Lepidopteran insects (36from H. armigera, 18 from H. virescens and 63 from B. mori)38,42,82,83. The IR datasets contained IR sequences from H. armigera (11), S. littoralis (11), Cydia pomonella (10), B. mori (18) and D. melanogaster (64)36,42,52,62. The OBP datasets contained sequences from H. armigera (26), H. virescens (17) and B. mori (34)37,42. The CSP data set contained sequences from H. armigera (13)42, H. virescens (9)84 and B. mori (16)70. All amino acid sequences were aligned using ClustalW285. The unrooted neighbor-joining trees were constructed by the Jones-Taylor-Thornton(JTT) method with 1,000 bootstrap replications as implemented in MEGA5 software86.
Expression analysis of the candidate receptors by semi-quantitative reverse transcription PCR
To illustrate and compare the expression patterns of candidate receptors in male and female antennae, semi-quantitative RT-PCR was performed using cDNA prepared from male antennae, female antennae and legs (male and female mixture). Legs were used as a control to confirm the antennae-enriched expression of candidate receptors. Total RNA was extracted as described above. Prior to cDNA synthesis, RNA was treated with DNase I (Fermentas, Vilnius, Lithuania) to remove trace amounts of genomic DNA. The cDNA was synthesized using the First Strand cDNA Synthesis Kit (Fermentas, Vilnius, Lithuania) and was used as a template in PCR reactions with gene-specific primers. The housekeeping gene RPS3 was used as a control87. Primers were designed using the Primer Premier 5 software (PREMIER Biosoft International), and the sequences are available in Supplementary Table S1. PCR was performed with the Veriti Thermal Cycler (Applied Biosystems, Carlsbad, CA, USA) under the following conditions: 94 °C for 2 min, 33 cycles of 94 °C for 30 s, 55–60 °C for 30 s, and 72 °C for 30 s, and 72 °C for 10 min. The cycle number was reduced to 27 and 30 for Actin and OR2 amplification because of their high expression level. The experiment was repeated three times using three independently isolated RNA samples. PCR amplification products were run on a 2% agarose gel and verified by DNA sequencing.
References
Sato, K. & Touhara, K. In Chemosensory Systems in Mammals, Fishes, and Insects 203–220 (Springer, 2009).
Gao, Q. & Chess, A. Identification of candidate Drosophila olfactory receptors from genomic DNA sequence. Genomics 60, 31–39 (1999).
Leal, W. S. Odorant reception in insects: roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 58, 373–391 (2013).
Clyne, P. J. et al. A novel family of divergent seven-transmembrane proteins: candidate odorant receptors in Drosophila. Neuron 22, 327–338 (1999).
Benton, R., Sachse, S., Michnick, S. W. & Vosshall, L. B. Atypical membrane topology and heteromeric function of Drosophilaodorant receptors in vivo. PLoS Biol. 4, e20 (2006).
Scott, K. et al. A chemosensory gene family encoding cadidate gustatory and olfacotry receoptors in Drosophila. Cell 104, 661–673 (2001).
Chyb, S. Drosophila gustatory receptors: from gene identification to functional expression. J. Insect Physiol. 50, 469–477 (2004).
Sparks, J. T., Vinyard, B. T. & Dickens, J. C. Gustatory receptor expression in the labella and tarsi of Aedes aegypti. Insect Biochem.Mol.Biol. 43, 1161–1171 (2013).
Jiang, X. J. et al. A gustatory receptor tuned to D-fructose in antennal sensilla chaetica of Helicoverpa armigera. Insect Biochem. Mol. Biol. 60, 39–46 (2015).
Mang, D. et al. Expression of a sugar clade gustatory receptor, BmGr6, in the oral sensory organs, midgut, and central nervous system of larvae of the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 70, 85–98 (2016).
Ning, C., Yang, K., Xu, M., Huang, L. Q. & Wang, C. Z. Functional validation of the carbon dioxide receptor in labial palps of Helicoverpa armigera moths. Insect Biochem. Mol. Biol. 73, 12–19 (2016).
Abuin, L. et al. Functional architecture of olfactory ionotropic glutamate receptors. Neuron 69, 44–60 (2011).
Ai, M. R. et al. Ionotropic glutamate receptors IR64a and IR8a form a functional odorant receptor complex in vivo in Drosophila. J. Neuroscience 33, 10741–10749 (2013).
Benton, R., Vannice, K. S., Gomez-Diaz, C. & Vosshall, L. B. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell 136, 149–162 (2009).
Silbering, A. F. et al. Complementary function and integrated wiring of the evolutionarily distinct Drosophilaolfactory subsystems. J. Neuroscience 31, 13357–13375 (2011).
Chen, C. et al. Drosophila ionotropic receptor 25a mediates circadian clock resetting by temperature. Nature 27, 516–U238 (2015).
Miyakawa, H., Sato, M., Colbourne, J. K. & Iguchi, T. Ionotropic glutamate receptors mediate inducible defense in the water flea Daphnia pulex. Plos One 10, e0121324 (2015).
Leal, W. S. Pheromone reception. In The Chemistry of Pheromones and Other Semiochemicals II 1–36 (Springer, 2005).
Pelosi, P., Zhou, J. J., Ban, L. P. & Calvello, M. Soluble proteins in insect chemical communication. Cell. Mol. Life Sci. 63, 1658–1676 (2006).
Laughlin, J. D., Ha, T. S., Jones, D. N. M. & Smith, D. P. Activation of pheromone-sensitive neurons is mediated by conformational activation of pheromone-binding protein. Cell 133, 1255–1265 (2008).
Zhou, J. J. Odorant-binding proteins in insects. Vitam Horm 83, 241–272 (2010).
Briand, L. et al. Characterization of a chemosensory protein (ASP3c) from honeybee (Apis mellifera L.) as a brood pheromone carrier. Eur. J. Biochem. 269, 4586–4596 (2002).
Yi, X. et al. Involvement of a specific chemosensory protein from Bactrocera dorsalis in perceiving host plant volatiles. J. Chem. Ecol. 40, 267–275 (2014).
Li, Z. Q. et al. Expression Analysis and Binding Assays in the Chemosensory Protein Gene Family Indicate Multiple Roles in Helicoverpa armigera. J. Chem. Ecol. 41, 473–485 (2015).
Vogt, R. G. et al. The insect SNMP gene family. Insect Biochem.Mol.Biol. 39, 448–456 (2009).
Gu, S. H. et al. Molecular identification and differential expression of sensory neuron membrane proteins in the antennae of the black cutworm moth Agrotis ipsilon. J. Insect Physiol. 59, 430–443 (2013).
Liu, S. et al. Identification and characterization of two sensory neuron membrane proteins from Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). Arch. Insect Biochem. 82, 29–42 (2013).
Zhang, J., Liu, Y., Walker, W. B., Dong, S. L. & Wang, G. R. Identification and localization of two sensory neuron membrane proteins from Spodoptera litura (Lepidoptera: Noctuidae). Insect Sci. 22, 399–408 (2015).
Furlong, M. J., Wright, D. J. & Dosdall, L. M. Diamondback moth ecology and management: problems, progress, and prospects. Ann. Rev. Entomol. 58, 517–541 (2013).
You, M. et al. A heterozygous moth genome provides insights into herbivory and detoxification. Nat. Genet. 45, 220–225 (2013).
Rogers, M. E., Krieger, J. & Vogt, R. G. Antennal SNMPs (sensory neuron membrane proteins) of Lepidoptera define a unique family of invertebrate CD36-like proteins. J. Neurobiol. 49, 47–61 (2001).
Benton, R., Vannice, K. S. & Vosshall, L. B. An essential role for a CD36-related receptor in pheromone detection in Drosophila. Nature 450, 289–293 (2007).
Zhang, J. et al. Antennal transcriptome analysis and comparison of chemosensory gene families in two closely related noctuidae moths, Helicoverpa armigera and H. assulta. PloS one 10, e0117054 (2015).
Grosse-Wilde, E. et al. Antennal transcriptome of Manduca sexta. Proc. Natl. Acad. Sci. USA 108, 7449–7454 (2011).
Xia, Q. et al. A draft sequence for the genome of the domesticated silkworm (Bombyx mori). Science 306, 1937–1940 (2004).
Olivier, V., Monsempes, C., Francois, M. C., Poivet, E. & Jacquin-Joly, E. Candidate chemosensory ionotropic receptors in a. Lepidoptera. Insect. Mol. Biol. 20, 189–199 (2011).
Gong, D. P., Zhang, H. J., Zhao, P., Xia, Q. Y. & Xiang, Z. H. The odorant binding protein gene family from the genome of silkworm. Bombyx mori. BMC Genomics 10, 332 (2009).
Tanaka, K. et al. Highly selective tuning of a silkworm olfactory receptor to a key mulberry leaf volatile. Curr. Biol. 19, 881–890 (2009).
Zhan, S., Merlin, C., Boore, J. L. & Reppert, S. M. The monarch butterfly genome yields insights into long-distance migration. Cell 147, 1171–1185 (2011).
Briscoe, A. D. et al. Female behaviour drives expression and evolution of gustatory receptors in butterflies. Plos Genet. 9, e1003620 (2013).
Howlett, N. et al. Identification of chemosensory receptor genes in Manduca sexta and knockdown by RNA interference. Bmc Genomics 13, 1 (2012).
Liu, Y., Gu, S., Zhang, Y., Guo, Y. & Wang, G. Candidate olfaction genes identified within the Helicoverpa armigeraantennal transcriptome. PloS One 7, e48260 (2012).
Cao, D. et al. Identification of candidate olfactory genes in Chilo suppressalis by antennal transcriptome analysis. Int. J. Biol. Sci. 10, 846–860 (2014).
Engsontia, P., Sangket, U., Chotigeat, W. & Satasook, C. Molecular evolution of the odorant and gustatory receptor genes in Lepidopteran insects: implications for their adaptation and speciation. J. Mol. Evol. 79, 21–39 (2014).
Legeai, F. et al. An Expressed Sequence Tag collection from the male antennae of the Noctuid moth Spodoptera littoralis: a resource for olfactory and pheromone detection research. BMC Genomics 12, 86 (2011).
Palanaswamy, P. & Seabrook, W. Behavioral responses of the female eastern spruce budwormChoristoneura fumiferana (Lepidoptera, Tortricidae) to the sex pheromone of her own species. J. Chem. Ecol. 4, 649–655 (1978).
Schneider, D., Schulz, S., Priesner, E., Ziesmann, J. & Francke, W. Autodetection and chemistry of female and male pheromone in both sexes of the tiger moth Panaxia quadripunctaria. J Comp. Physiol. a-Sensory Neural. Behav.Physiol. 182, 153–161 (1998).
Jones, W. D., Nguyen, T. A., Kloss, B., Lee, K. J. & Vosshall, L. B. Functional conservation of an insect odorant receptor gene across 250 million years of evolution. Curr. Biol. 15, 119–121 (2005).
Nakagawa, T., Sakurai, T., Nishioka, T. & Touhara, K. Insect sex-pheromone signals mediated by specific combinations of olfactory receptors. Science’s STKE 307, 1638 (2005).
Montell, C. A taste of the Drosophila gustatory receptors. Curr. Opin. Neurobiol. 19, 345–353 (2009).
Clyne, P. J., Warr, C. G. & Carlson, J. R. Candidate taste receptors in Drosophila. Science 287, 1830–1834 (2000).
Bengtsson, J. M. et al. Putative chemosensory receptors of the codling moth, Cydia pomonella, identified by antennal transcriptome analysis. PloS One 7, e31620 (2012).
Jorgensen, K., Kvello, P., Almaas, T. J. & Mustaparta, H. Two closely located areas in the suboesophageal ganglion and the tritocerebrum receive projections of gustatory receptor neurons located on the antennae and the proboscis in the moth Heliothis virescens. J. Comp. Neurol. 496, 121–134 (2006).
Jacquin-Joly, E. et al. Candidate chemosensory genes in female antennae of the noctuid moth Spodoptera littoralis. Int. J. Biol. Sci. 8, 1036–1050 (2012).
Wanner, K. W. & Robertson, H. M. The gustatory receptor family in the silkworm moth Bombyx mori is characterized by a large expansion of a single lineage of putative bitter receptors. Insect Mol. Biol. 17, 621–629 (2008).
Xu, W., Papanicolaou, A., Liu, N. Y., Dong, S. L. & Anderson, A. Chemosensory receptor genes in the Oriental tobacco budworm Helicoverpa assulta. Insect Mol. Biol. 24, 253–263 (2015).
Jones, W. D., Cayirlioglu, P., Grunwald Kadow, I. & Vosshall, L. B. Two chemosensory receptors together mediate carbon dioxide detection in Drosophila. Nature 445, 86–90 (2007).
Kwon, J. Y., Dahanukar, A., Weiss, L. A. & Carlson, J. R. The molecular basis of CO2 reception in Drosophila. Proc. Natl. Acad. Sci. USA 104, 3574–3578 (2007).
Lu, T. et al. Odor coding in the maxillary palp of the malaria vector mosquito Anopheles gambiae. Curr. Biol. 17, 1533–1544 (2007).
Robertson, H. M. & Kent, L. B. Evolution of the gene lineage encoding the carbon dioxide receptor in insects. J. Insect Sci. 9, 14 (2009).
Kent, L. B., Walden, K. K. & Robertson, H. M. The Gr family of candidate gustatory and olfactory receptors in the yellow-fever mosquito Aedes aegypti. Chem. Senses 33, 79–93 (2008).
Croset, V. et al. Ancient protostome origin of chemosensory ionotropic glutamate receptors and the evolution of insect taste and olfaction. PLoS Genet. 6, e1001064 (2010).
Zhang, Y. N. et al. Differential expression patterns in chemosensory and non-chemosensory tissues of putative chemosensory genes identified by transcriptome analysis of insect pest the purple stem borer Sesamia inferens (Walker). Plos One 8, e6971510 (2013).
Feng, B. et al. Transcriptome and expression profiling analysis link patterns of gene expression to antennal responses in Spodoptera litura. BMC Genomics 16, 269 (2015).
Poivet, E. et al. A comparison of the olfactory gene repertoires of adults and larvae in the noctuid moth Spodoptera littoralis. PloS one 8, e60263 (2013).
Zhang, Z. C., Wang, M. Q., Lu, Y. B. & Zhang, G. Molecular characterization and expression pattern of two general odorant binding proteins from the diamondback moth, Plutella xylostella. J.Chem. Ecol. 35, 1188–1196 (2009).
Garczynski, S. F. et al. Application of Cydia pomonella expressed sequence tags: Identification and expression of three general odorant binding proteins in codling moth. Insect Sci. 20, 559–574 (2013).
Liu, N.-Y. et al. Two general-odorant binding proteins in Spodoptera litura are differentially tuned to sex pheromones and plant odorants. Comp Biochem PhysiolPart A: MolIntegr Physiol. 180, 23–31 (2015).
Zhu, J. et al. General odorant-binding proteins and sex pheromone guide larvae of Plutella xylostella to better food. Insect Biochem. Mol. Biol. 72, 10–19 (2016).
Foret, S., Wanner, K. W. & Maleszka, R. Chemosensory proteins in the honey bee: insights from the annotated genome, comparative analyses and expressional profiling. Insect Biochem. Mol. Biol. 37, 19–28 (2007).
Pelosi, P., Iovinella, I., Felicioli, A. & Dani, F. R. Soluble proteins of chemical communication: an overview across arthropods. Front.Physiol. 5, 320 (2014).
Gong, D. P. et al. Identification and expression pattern of the chemosensory protein gene family in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 37, 266–277 (2007).
Jacquin-Joly, E., Vogt, R. G., Francois, M. C. & Nagnan-Le Meillour, P. Functional and expression pattern analysis of chemosensory proteins expressed in antennae and pheromonal gland of Mamestra brassicae. Chem. Senses 26, 833–844 (2001).
Liu, Y. L., Guo, H., Huang, L. Q., Pelosi, P. & Wang, C. Z. Unique function of a chemosensory protein in the proboscis of two Helicoverpa species. J. Exp. Biol. 217, 1821–1826 (2014).
Liu, C., Zhang, J., Liu, Y., Wang, G. & Dong, S. Eexpression of SNMP1 and SNMP2 genes in antennal sensilla of Spodoptera exigua (Hübner). Arch. Insect Biochem. Physiol. 85, 114–126 (2014).
Jin, X., Ha, T. S. & Smith, D. P. SNMP is a signaling component required for pheromone sensitivity in Drosophila. Proc. Natl. Acad. Sci. USA 105, 10996–11001 (2008).
Grabherr, M. G. et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nature biotechnol. 29, 644–652 (2011).
Pertea, G. et al. TIGR Gene Indices clustering tools (TGICL): a software system for fast clustering of large EST datasets. Bioinformatics 19, 651–652 (2003).
Altschul, S. F. et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402 (1997).
Iseli, C., Jongeneel, C. V. & Bucher, P. ESTScan: a program for detecting, evaluating, and reconstructing potential coding regions in EST sequences. Proc. Int. Conf. Intell. Syst. Mol. Biol. 138–148 (1999).
Petersen, T. N., Brunak, S., von Heijne, G. & Nielsen, H. SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat. Methods 8, 785–786 (2011).
Krieger, J. Genes encoding candidate pheromone receptors in a moth (Heliothis virescens). Proc. Natl. Acad. Sci. USA 101, 11845–11850 (2004).
Krieger, J. et al. A divergent gene family encoding candidate olfactory receptors of the moth Heliothis virescens. Eur. J. Neurosci. 16, 619–628 (2002).
Picimbon, J. F., Dietrich, K., Krieger, J. & Breer, H. Identity and expression pattern of chemosensory proteins in Heliothis virescens (Lepidoptera, Noctuidae). Insect Biochem. Mol. Biol. 31, 1173–1181 (2001).
Larkin, M. A. et al. Clustal W and Clustal X version 2.0. Bioinformatics 23, 2947–2948 (2007).
Tamura, K. et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739 (2011).
Wanner, K. W. et al. Female-biased expression of odourant receptor genes in the adult antennae of the silkworm, Bombyx mori. Insect Mol. Biol. 16, 107–119 (2007).
Acknowledgements
This work was funded by Beijing Nova Program (Z161100004916119), the National Natural Science Foundation of China (31672095&31471833) and the Ministry of Education of Anhui Province (gxbjZD2016016). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
S.Y., G.W. and Y.L. designed the experiments. S.Y., D.C. and Y.L. performed the experiments. S.Y., D.C. and Y.L. contributed reagents/materials/gene identification. S.Y., D.C., G.W. and Y.L. analyzed the data. S.Y., G.W. and Y.L. wrote the paper.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yang, S., Cao, D., Wang, G. et al. Identification of Genes Involved in Chemoreception in Plutella xyllostella by Antennal Transcriptome Analysis. Sci Rep 7, 11941 (2017). https://doi.org/10.1038/s41598-017-11646-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-11646-7
- Springer Nature Limited
This article is cited by
-
Genomic identification and evolutionary analysis of chemosensory receptor gene families in two Phthorimaea pest species: insights into chemical ecology and host adaptation
BMC Genomics (2024)
-
Antifeedant properties and contact toxicities of the trichocellins A-I and B-II from a Trichoderma reesei against Plutella xylostella larvae
International Journal of Tropical Insect Science (2022)
-
Antennal transcriptome analyses and olfactory protein identification in an important wood-boring moth pest, Streltzoviella insularis (Lepidoptera: Cossidae)
Scientific Reports (2019)
-
Identification and Expression Profiling of Odorant Receptor Protein Genes in Sitophilus zeamais (Coleoptera: Curculionoidea) Using RT-qPCR
Neotropical Entomology (2019)
-
Identification of candidate chemosensory genes in Mythimna separata by transcriptomic analysis
BMC Genomics (2018)