
Abstract
Epidemiological studies have found a higher prevalence of allergic symptoms and positive prick tests in patients with Meniere’s disease (MD); however the effect of allergenic extracts in MD has not been established. Thus, this study aims to determine the effect of Aspergillus and Penicillium stimulation in cytokine release and gene expression profile in MD. Patients with MD showed higher basal levels of IL-1β, IL-1RA, IL-6 and TNF-α when compared to healthy controls. We observed that IL-1β levels had a bimodal distribution suggesting two different subgroups of patients, with low and high basal levels of cytokines. Gene expression profile in peripheral blood mononuclear cells (PBMC) showed significant differences in patients with high and low basal levels of IL-1β. We found that both mold extracts triggered a significant release of TNF-α in MD patients, which were not found in controls. Moreover, after mold stimulation, MD patients showed a different gene expression profile in PBMC, according to the basal levels of IL-1β. The results indicate that a subset of MD patients have higher basal levels of proinflammatory cytokines and the exposure to Aspergillus and Penicillium extracts may trigger additional TNF-α release and contribute to exacerbate inflammation.
Similar content being viewed by others


Introduction
Meniere’s disease (MD) was first described by Prosper Menière in 1861. It is a clinical syndrome characterized by episodes of spontaneous vertigo lasting minutes to hours, usually associated with fluctuating, low to medium frequencies in sensorineural hearing loss (SNHL), tinnitus and aural fullness and it may affect one or both ears (bilateral MD). The diagnostic criteria for MD are based in clinical symptoms and they were reformulated jointly by the Classification Committee of the Bárány Society, The Japan Society for Equilibrium Research, the European Academy of Otology and Neurotology (EAONO), the Equilibrium Committee of the American Academy of Otolaryngology-Head and Neck Surgery (AAO-HNS) and the Korean Balance Society in 20151, but they do not consider the clinical heterogeneity observed among patients which may respond to different mechanisms of disease.
The prevalence of MD is about 0.5-1/1000 individuals, being most of patients considered sporadic, while 9–10% are familial cases in European population2,3,4. The pathophysiology of MD is associated with the accumulation of endolymph in the cochlear duct and vestibular organs5. Although a third of MD cases may have a dysfunction of the immune system6, the immune response in patients with MD has been seldom investigated7.
Cluster analyses using few clinical variables in a large cohort of patients with MD have identified five clinical subgroups in patients with unilateral and bilateral MD8,9. So, for unilateral MD, group 1 was the most frequently found (53%) and it was defined as sporadic MD without migraine nor autoimmune comorbidity; group 2 is defined by SNHL which precedes the vertigo episodes by months or years (delayed MD); group 3 includes familial cases of MD; group 4 is associated with migraine and group 5 is defined by another comorbid autoimmune disorder9.
Although these clinical subgroups of patients point to different potential mechanisms for the development of endolymphatic hydrops, the cellular and molecular bases of MD largely remain to be defined.
The link between allergy and MD was first described by Duke in 192310. He treated patients with epinephrine with resolution of symptoms. In 2000, Derebery conducted epidemiological studies in patients with MD finding that 58% of patients had a history of allergy and 41% presented a positive skin prick test11. Both, inhalatory and food allergens have been associated with MD12. In a non-controlled study, Topuz showed that prick test may induce aural symptoms such as tinnitus or aural fullness in 62% of patients and an increase of endolymphatic pressure measured by electrocochleography in 77% of cases13. This study suggests that certain allergens may induce endolymphatic hydrops. The role of allergy in the pathogenesis of MD was also evaluated by Keles et al.14 who found significant differences in CD4, CD4/CD8, CD23 lymphocytes subpopulations and IFN-γ and IL4 levels in patients when they were compared to controls. The prevalence of allergy was also higher in MD patients after evaluating IgE levels (10). Fuse et al. analyzed intracellular cytokine levels in patients with acute low-tone hearing loss (ALHL) and MD and they found an imbalance in the Th1/Th2 response in ALHL and an increased natural killer cell activity in MD15. In addition, there are other evidences supporting an altered immune response in MD, including the increased values of circulating immune complexes (CIC) in the serum of some patients16,17 or the association of allelic variants in the MICA, TLR10 or NFκB genes with the progression of SNHL in MD18,19,20.
The aim of this study is to determine the proinflammatory cytokines profile in PBMCs and to investigate the effect of mold allergenic extracts in the proinflammatory response in patients with MD.
Results
Clinical features in MD patients
Patients with MD experienced attacks of vertigo with a progressive SNHL during the first years of the disease. Table 1 compares the clinical features of all 113 MD patients with uni- and bilateral MD. Patients with bilateral SNHL had a longer duration of the disease (P = 8.0 × 10−4), worse hearing stage (P = 0.03), higher frequency of cardiovascular risk factors (high blood pressure (P = 4 × 10−3) and type 2 diabetes (P = 0.024), and a higher frequency of episodes of sudden falls without spinning termed Tumarkin crisis (P = 6 × 10−3).
PBMC from MD patients have higher basal levels of cytokines when compared to controls
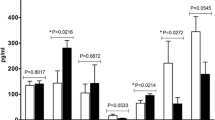
Basal levels of proinflammatory cytokines in the supernatant of 113 patients and 54 healthy controls were tested by Bead-based multiplex immunology assays. When compared to controls, MD patients had 2.8 fold higher expression of Interleukin-1 beta (IL-1β; P = 3.8 × 10−4); 3.8 fold higher Interleukin-1 receptor antagonist (IL-1RA, P = 2.74 × 10−4); 7.1 fold higher Interleukin-6 (IL-6; P = 7.83 × 10−4) and 2.6 fold more Tumor necrosis factor alpha (TNF-α; P = 3.5 × 10−5) (Fig. 1A).

Cytokines at basal levels and after mold stimulation in PBMC. (A) All cases and controls at basal levels according to the levels of IL-1β. (B) Scattered-plot of MD patients according to the levels IL-1β, TNF-α and IL-6. Dots in green represent the patients with low levels of IL-1β (≤5.9 pg/mL) whereas dots in purple are the patients with high levels of IL-1β (>5.9 pg/mL) (C) All cases and controls after stimulation with Aspergillus. (D) All cases and controls after stimulation with Penicillium. *Significant p-values. IL-1RA, IL-1 receptor antagonist.
Figure 1B shows the distribution of MD patients according to the levels IL-1β, TNF-α and IL-6. The basal levels of IL-1β in patients with MD showed a bimodal distribution, suggesting that two different subgroups of patients with MD can be distinguished according to the levels of cytokines in PBMC supernatant. So, 89 patients had low basal levels of IL-1β and 24 of 113 patients (21%) showed high basal levels of IL-1β (>mean + 2 standard deviation = 5.9 pg/mL; Supplementary Table S1). However, all control individuals had very low basal values of all measured cytokines.
Table 2 includes all patients with high basal levels of IL-1β. Twelve of them (50%) had been diagnosed as unilateral MD and another have 12 bilateral MD. Most of these patients were sporadic cases (22/24) and they were classified within the subgroup 1 (classic MD unilateral without migraine and they did not presented another comorbid autoimmune condition). Twenty-one of 24 (87.5%) showed elevated levels of IL-6, 19/24 (79.2%) has also higher levels of TNF-α and 17/24 (70.8%) presented an elevation of IL-1RA.
Mold allergenic extracts induce a higher TNF-α production in PBMCs from MD patients when they are compared to controls
PBMCs from all MD patients and healthy controls were stimulated with mold protein extracts (5μg/mL) for 16 hours and were compared to unstimulated PBMCs. When PBMCs were stimulated with Alternaria alternata no differences were found in IL-1β, IL-6, IL-1RA or TNFα levels between MD patients and healthy controls (Data not shown). However, when PBMCs were stimulated with Aspergillus (Fig. 1C) or Penicilium (Fig. 1D), MD patients showed higher levels TNF-α and IL-6, but no significant elevation in IL-1β.
When compared to healthy controls, PBMC of patients with high basal levels of IL-1ß had 3.2 fold higher expression of TNF-α in response to Aspergillus mix (p= 2.9 × 10−3), and 2.2 fold higher expression of TNF-α in response to Penicillium mix (p = 0.02). Patients with low levels of IL-1β presented high levels of TNF-α (2.7, P = 5.0 × 10−6) in response to Aspergillus and 2.7 fold higher (p = 6 × 10−6) in response to Penicillium. Of note, when we compared between MD patients with high and low basal of IL-1β after stimulation with either Aspergillus or Penicillium we found no differences in their cytokines release.
Gene expression in PBMCs from MD patients with low and high levels of IL-1ß
When we compared the gene expression profiles of stimulated and non-treated PBMCs from 15 MD patients and 11 healthy individuals we found 1099 differentially expressed genes (DEG, >2-fold change, p < 0.02).
In order to examine the complexities of the multivariate data we performed a principal component analysis (PCA) revealing great differences in the expression within each group (Fig. 2). Hence, because of the inter-individual variability, we decided to perform pairwise analysis in order to compare groups. Figure 2A shows a hierarchical cluster with 2 major groups: gene expression before and after stimulation. Loading plots represent DEGs in both, patients and controls (Fig. 2B), controls (Fig. 2C) and MD patients (Fig. 2D) before and after Penicillium and Aspergillus stimulation. Healthy controls showed the most significant differences in their gene expression profiles.

Gene expression in PBMC. (A) Dendrogram showing hierarchical clustering of patients, according to the gene expression in PBMCs. (B) Three dimensional loading plot showing principal component analysis (PCA) eigenvectors using all datasets. (C) Loading plots in controls before and after stimulation. (D) Loading plots in MD patients before and after stimulation.
PBMC gene expression at basal levels
We compared the gene expression profile in non-treated PBMCs from patients with high (N = 3) and low (N = 4) basal levels of IL-1β. We found that PBMCs from patients with high basal levels of IL-1β have 58 DEG (p-value cutoff 0.05 and Fold Change >1.2) when they were compared to patients with low levels of cytokines, being the most significant gene CXCL5 with a 2.5 Fold change (adjusted p = 0.03). We then performed a Core analysis using IPA® software and three different networks were retrieved. The top ranked network had a score of 38 and involved 16/58 genes (Fig. 3). All 16 genes are involved in cellular function and maintenance, protein synthesis and tissue morphology. Remarkably, 21/58 genes are involved in immunological diseases and 32/58 in organismal Injury and abnormalities such as inflammation.

Top Network from MD patients PBMCs at basal levels. Network retrieved after comparing by pairwise analysis gene expression data from MD patients PBMCs with high and low basal levels of IL-1β. Genes in red were up-regulated, while genes in green were down-regulated.
Mold-induced gene expression in PBMC
We compared the gene expression profile in Aspergillus or Penicillium –treated and non-treated PBMCs from 15 MD patients and 11 healthy individuals. Figure 4A shows a Venn diagram showing the number of DEG in controls and patients with low and high levels of IL-1β, after Aspergillus stimulation (Supplementary Table S2). One hundred and fifty-three genes were differentially expressed in the 3 groups and a top network with 29 focus molecules was retrieved, after performing a core analysis (score = 55; Fig. 4B). Of note, 132 DEG were found in PBMC from patients with low levels of IL-1β retrieving a top network with 20 focus molecules (score = 35; Fig. 4C), and 75 unique DEG in PBMC from patients with high levels of IL-1β, retrieving a top network with 19 focus molecules (score = 41; Fig. 4D).

Differentially expressed genes after Aspergillus stimulation. (A) Venn diagram showing the number of differentially expressed genes in controls, MD patients with low basal levels of IL-1β and patients with high basal levels of IL-1β. (B) Network obtained after taking common genes to all groups. (C) Network retrieved in MD patients with low basal levels of IL-1β unique DEG. (D) Network obtained after taking MD patients with high basal levels of IL-1β unique DEG. Genes in red were up-regulated, while genes in green were down-regulated.
Next, we compared the gene expression profiles after stimulating PBMC with Penicilium extract (Supplementary Table S3). Figure 5 shows a Venn diagram from controls and patients with high and low levels of IL-1β. One hundred and forty-six DEG were common in all three groups. A top network with 25 focus molecules was obtained (score = 45; Fig. 5B). Remarkably, 95 unique DEG were found in PBMC from patients with low levels of IL-1β and 680 genes were differentially expressed in patients with high levels of IL-1β. Figure 5C (Focus molecules = 21; score = 41) and 5D (Focus molecules = 28; score = 37) include the networks retrieved for each group, respectively.

Differential expressed genes after Penicillium stimulation. (A) Venn diagram showing the number of differentially expressed genes in controls, MD patients with low basal levels of IL-1β and patients with high basal levels of IL-1β. (B) Network obtained after taking common genes to all groups. (C) Network obtained after taking MD patients with low basal levels of IL-1β unique DEG. (D) Network obtained after taking MD patients with high basal levels of IL-1β unique DEG. Genes in red were up-regulated, while genes in green were down-regulated.
Discussion
Our study has found that basal levels of proinflammatory cytokines may be increased in some patients with MD. This observation is found in both patients, with uni- or bilateral MD. Furthermore, we have found that both Aspergillus and Penicillium extracts may induce a release of TNF-α, which is not observed in controls.
This cytokine profile may represent two subgroups of MD patients with intrinsic differences in the immune response or two functional states of the immune system in patients with MD. Previous studies have defined five clinical subgroups in patients with uni and bilateral MD9,21. Unilateral MD type 1 is the most common clinical variant and it includes patients without migraine, no concomitant autoimmune disease nor familial history of MD. Most of the patients with increased IL-1β were classified as MD type 1 (83.4%), 8.3% were MD type 3 and 8.3% were MD type 4. None of the patients with elevated proinflammatory cytokines had an acute infection or another autoimmune condition that could explain this finding. So, this subset of MD patients with high basal levels of cytokines and negative autoantibodies may be considered as an autoinflammatory condition involving the inner ear. The field of autoinflammatory disorders has expanded from rare monogenic diseases to polygenic autoinflammatory diseases such Crohn disease, Behcet’s disease or sarcoidosis22,23. The characteristics of autoinflammatory disease of the inner ear may include an innate immune response mediated by monocytes, high levels of IL-1β and low titers/non-specific autoantibodies24.
Moreover, our findings show that Aspergillus and Penicillium extracts trigger a significant TNF-α release in PBMC from patients with MD that contribute to exacerbate the inflammatory response.
The human genes in the IL-1 complex encode for three cytokines: IL-1α, IL-1β and the antagonist receptor IL-1RA. The severity of a given inflammation is influenced by the balance between the levels of IL-1β and that of IL-1 RA. In healthy individuals, IL-1 RA is readily detectable in plasma and IL-1 β levels are usually undetectable25. Remarkably, IL-1RA levels were found to be much higher elevated in most patients with MD regardless of the levels of IL-1β. Since TLR10 is an anti-inflammatory pattern-recognition receptor able to up-regulate IL-1RA26, and the TLR10 allelic variant I369L rs11096955 has been associated with faster hearing loss progression in bilateral MD19, the activation of TLR10 could explain the increased levels of IL-1RA found in patients with MD. Moreover, individuals bearing I369L rs11096955 displayed increased levels of IL-1β, TNF-α and IL-6 upon ligation of TLR226.
This study demonstrates that allergenic extracts from Aspergillus and Penicillium induce a proinflammatory immune response in most patients with MD involving TNF-α, which it is not observed in controls. This mechanism could initiate or exacerbate the inflammatory response in the endolymphatic sac or the spiral ligament leading to the development of endolymphatic hydrops. Previous studies also have established a relationship between mold extracts and proinflammatory cytokines in autoimmune inner ear disease (AIED)27. In AIED patients, isolated PBMC exposed to mold increased IL-1β gene expression, and enhanced IL-6 and IL-1β secretion, that could act as a Pathogen-Associated Molecular Pattern (PAMP) in a subset of them. Since most of our patients with MD were classified as MD type 1 and did not have a comorbid autoimmune disease, and we also excluded patients with response to inhalatory allergens in the prick test (including Penicillium and Aspergillus), the elevation of TNF-α observed in patients with MD after exposure of PBMC to Penicillium and Aspergillus cannot be explained by another autoimmune condition or previous sensitization to other allergens.
Furthermore, it is well established that molds can cause respiratory symptoms and may worsen the course of bronchial asthma, which could lead to increase secretion of TNF-α, IL-6, IFN-γ or IL-1β28,29.
Our data show that Aspergillus and Penicillium trigger similar immune responses, according to the gene expression profile in PBMC, but few genes were specifically triggered by Penicillium in patients with high basal levels of IL-1β such as TNFRSF6B, CXCR4 or CD14 (up-regulated) and FCGR3B or HLA-DRB4 (down-regulated). In addition, some genes were also specifically expressed in patients with low basal levels of IL-1β such as VEGFA, LAIR2 or CD82 (up-regulated) and CX3CR1 or SNAP23 (down-regulated). We found that IL6 and CXCL5 genes were key mediators in molds-trigger response in all controls and patients with MD, regardless of the IL-1β levels.
We also observed that only 58 genes were differentially expressed in MD patients with high basal levels of IL-1β, after stimulation with either Aspergillus or Penicillium. This gene expression profile was not found neither in controls or patients with low levels of IL-1β, supporting the hypothesis that this abnormal immune response with high levels of IL-1β may facilitate the identification of a subgroup of MD patients with elevated proinflammatory cytokines and a specific mold-induced gene expression profile. A major part of the genes retrieved from our data revealed networks associated to cellular movement, immune cell trafficking, cell morphology and molecular transport. Thus, the probable pathways involved within these networks would be Granulocyte/Agranulocyte Adhesion and Diapedesis, Communication between Innate and Adaptative Immune cells and Role of Cytokines in mediating communication between immune cells.
Our results add new evidence to support that MD is a heterogeneous disorder and the levels of proinflammatory cytokines and the differences in the immune response to certain pathogens, including molds, may explain that some patients respond to drugs with antihistaminic effects, such as betahistine, but most patients do not show a response different from placebo30. Moreover, since patients using immunotherapy have reported an improvement of the duration and frequency of the vertigo episodes compared to controls11, the assessment of allergic response in patients with MD, and the use of antihistaminic or immunotherapy as part of the treatment plan to control vertigo attacks should be considered.
The major limitation of our study is that the number of investigated patients was low (N = 113), and further patients will need to be tested to validate our findings. However, we found that 21% of MD patients had higher basal levels of IL-1β. Pilot clinical trials will determine if these individuals may benefit from target therapies to block IL-1β such as anakinra (recombinant anti-IL1RA) and canakinumab (anti-IL1β monoclonal antibody), as it has been shown in AIED31.
Conclusions
1.A subset of patients with Meniere disease has elevated levels of proinflamatory cytokines and they may have an autoinflammatory inner ear disease.
2.Aspergillus and Penicillium trigger the release of TNF-α in MD patients and this could initiate or exacerbate the inflammatory response in the inner ear.
Materials and Methods
Human subjects
This study included a total of 113 patients with definite MD according to the diagnostic criteria of the Barany Society1 and 54 healthy controls. All patients had, at least, 3 years of follow-up since the onset of the disease.
Individuals over 18 years old were recruited from 9 otoneurology clinics from January 2015 to December 2016. Control subjects matching sex and age with MD patients were selected (mean age 54 ± 12, 55.3% women). All participants underwent allergic screening by dermal prick test (ALK Abello, A/S, Hørsholm, Denmark) to respiratory allergens. All individuals showed negative response to the prick test and those under steroid therapy during the last 3 months were excluded from the study.
All patients and healthy individuals gave their informed consent for the study. Blood samples were collected during the quiescent phase of the disease. The study was carried out according to the principles of the Declaration of Helsinki revised in 201332 for investigation with humans. The Almeria Ethical Review board for clinical research approved the research protocol PI13-01242.
PBMC culture and stimulation
Peripheral blood mononuclear cells (PBMC) were isolated and cultured in RPMI 1640 supplemented with 10% (v/v) fetal bovine serum (Biowest, Nuaillé, France) as previously described33. Cells were plated at 1 × 106 cells/mL in 12-well plates. In some experiments, fungus extracts were added (2 different Allergenic Extract-Mix fungus containing, in one case, 4 equal parts of Mix Aspergillus (oryzae, repens, niger and terreus; code#1034497) and in the other case, 4 equal parts of Mix Penicillium (brevicompactum, expansum, notatum and roqueforti; code#1034760)). Additionally, Alternaria alternata was used in some experiments (ALK Abelló A/S, Hørsholm, Denmark). Each combination was dialyzed to remove phenol by using Tube-o-dialyzer (VWR Intranational, Radnor, PA, USA) against deionized distilled water. Fungus protein concentration was analyzed by Bradford dye-binding method protein assay (Bio-Rad Laboratories, Hercules, CA, USA)34. One × 106 PBMC per mL was stimulated with 5 μg/mL of each dialyzed mix fungus. PBMC were incubated during 16 h at 37 °C in 7% CO2 and compared with cultured, unstimulated PBMC. The optimal concentrations of these mixtures were previously identified by culturing PBMC with increasing concentrations (5, 10, 20 μg/mL) of these reagents to identify the best concentration (mold mix: 5 μg/mL) that could be used without distressing cell viability (viability >80%). At the end of all incubations, PBMC were centrifuged, RNA and proteins harvested and supernatants were collected and stored at −20 °C.
Cytokine Measurement
Conditioned supernatants were collected and stored at −20 °C until a sufficient number of samples were acquired. Frozen samples were thawed immediately prior to analysis and none of the samples underwent repetitive freeze-thawing cycles prior to analysis. IL-1β, IL-6, IL-1RA and TNFα levels in conditioned supernatant were quantified simultaneously with a commercially available Multiplex Bead-Based Kit (EMD Millipore, Billerica, MA, USA), in accordance with the kit-specific protocols provided by Millipore, using a Luminex 200 (Luminex Corp., Austin, TX, USA) and read with Luminex × PONENT 3.1 software.
The minimum detection limit for the assay was 0.14 pg/mL and the maximum 10000 pg/mL. Samples with readings below or above these levels were assigned values of 0 pg/mL for the minimum value or 10000 pg/mL for the maximum value. The percentages of samples with levels below 0.14 pg/mL (detection limit for unstimulated samples) were: IL-1β 0.08%, IL-1RA 0.03%, IL-6 0.006% and TNF-α 0%. The percentages of samples with levels above 10000 pg/mL (detection limit for samples stimulated with Aspergillus) were: IL-1β 0.006%, IL-1RA 0%, IL-6 0.06% and TNF-α 0.047%; and for samples stimulated with Penicillium were: IL-1β 0.012%, IL-1RA 0%, IL-6 0.05% and TNF-α 0.6%.
We measured the cytokines in duplicate and determined the reliability of the measurements by calculating the Intraclass Correlation Coefficient (ICC). ICC for each cytokine was greater than 80% in samples stimulated with Mix Apergillus: IL-1β 97.8%, IL-1RA 98.2%, IL-6 81% and TNF-α 94%; and samples stimulated with Mix Penicillium: IL-1β 94.9%, IL-1RA 90.2%, IL-6 84.5% and TNF-α 91.7%. In addition, we had two quality controls for each cytokine (provided by the manufacturer with the kit), run in duplicate, which were within the expected range with coefficients of variation less than 8%.
IL-1β, IL-6 ELISA
IL-1β and IL-6 levels were quantified, in 30 conditioned supernatant samples, using a sandwich ELISA (R&D Systems, Minneapolis, MN, USA) as described by the manufacturer’s procedure. All samples were run in duplicate. The minimum detection limit for the assays was 0.17 pg/mL and the maximum 450 pg/mL. Samples with readings below or above these levels were assigned values of 0 pg/mL for the minimum value. No sample was above the maximum value. Correlation between ELISA and Multiplex Bead-Based assay was greater than 85% for both cytokines.
RNA extraction
RNA was isolated using the High Pure RNA isolation Kit (Hoffmann-La Roche, Basel, Switzerland) according to the manufacturer’s protocol. RNA concentration was measured on a Nanodrop (NanoDrop Technologies Inc.,Wilmington, DE, USA). RNA quality was checked using Agilent 2100 Bioanalyzer (Agilent Technologies, Waldbronn, Germany).
Expression array
HumanHT-12 v4 Expression BeadChip (Illumina Inc., San Diego, CA, USA) provides genome-wide transcriptional coverage of more than 47,000 probes, including well-characterized genes, gene candidates, and splice variants. In short biotin-labeled
cRNA samples were prepared as described by manufacturer’s protocol using 500ng of purified total RNA as a template for the reaction and then, processed with the high resolution scanner iScan (Illumina Inc., San Diego, CA, USA). Probe intensity data were analyzed using Illumina’s GenomeStudio software (Gene Expression Module). In order to determine gene average expression levels a median normalization was used according to control probe profiles.
Statistical analysis
A descriptive statistical analysis for clinical data was performed using SPSS software v.22 (SPSS Inc, Chicago, IL, USA). Data are shown as means ± standard deviation (SD). Quantitative variables were compared using Student’s unpaired T-test. Qualitative variables were compared using crosstabs and Fisher’s exact test. Nominal p-values < 0.05 were considered statistically significant.
The R statistical software version 3.2.3 and RStudio were used for all analyses in gene expression data (www.r-project.org). Limma R package from Bioconductor was used for expression data analysis, normalization and differential expression analysis. We used a pairwise t-statistic approach for each comparison and followed Benjamini and Hochberg’s correction in order to control the false discovery rate. An adjusted P value cutoff of 0.02 was considered for differentially expressed genes (DEG).
Signaling pathway analysis was performed using Ingenuity Pathways Analysis software (IPA®, Qiagen, Venlo, Netherlands, http://www.ingenuity.com/products/ipa). Core analysis tool was executed using DEG.
A PCA was performed for gene expression data using DEG (dots = genes; lines with arrows = MD patients and controls before and after stimulation; 3-axis = PC1, PC2 and PC3). Loading plots were generated to visualize the relationship between groups. Using this method, the basal levels and after stimulation were separated using PC1, PC2, and PC3, according to DEG. IPA network scores are derived from p-values. If are n genes in the network and f of them are Focus Genes. The p-value is the probability of finding f or more Focus Genes in a set of n genes randomly selected from the Global Molecular Network database in IPA. It is calculated using Fisher’s exact test as described in IPA user’s instructions. Therefore, the p-score is defined as: p-score = −log10 (p-value).
We compared the gene expression profile of stimulated and non-stimulated PBMCs using the web-based software GeneSifter performing a 2-way ANOVA, in order to define a significant effect of Penicillium and Aspergillus stimulation, or any difference between patients with low and high basal levels of IL-1β (P < 0.02; threshold 2-fold change).
Data availability
The datasets generated in this study are available from the corresponding author on reasonable request and can be obtained from Gene Expression Omnibus with the accession number GSE109558.
References
Lopez-Escamez, J. A. et al. Diagnostic criteria for Meniere’s disease. Journal of vestibular research: equilibrium & orientation 25, 1–7, https://doi.org/10.3233/ves-150549 (2015).
Morrison, A. W., Bailey, M. E. & Morrison, G. A. Familial Meniere’s disease: clinical and genetic aspects. The Journal of laryngology and otology 123, 29–37, https://doi.org/10.1017/s0022215108002788 (2009).
Vrabec, J. T. Genetic investigations of Meniere’s disease. Otolaryngologic clinics of North America 43, 1121–1132, https://doi.org/10.1016/j.otc.2010.05.010 (2010).
Requena, T. et al. Familial clustering and genetic heterogeneity in Meniere’s disease. Clinical genetics 85, 245–252, https://doi.org/10.1111/cge.12150 (2014).
Hallpike, C. S. & Cairns, H. Observations on the Pathology of Ménière’s Syndrome: (Section of Otology). Proceedings of the Royal Society of Medicine 31, 1317–1336 (1938).
Greco, A. et al. Meniere’s disease might be an autoimmune condition? Autoimmunity reviews 11, 731–738, https://doi.org/10.1016/j.autrev.2012.01.004 (2012).
Kim, S. H. et al. Autoimmunity as a candidate for the etiopathogenesis of Meniere’s disease: detection of autoimmune reactions and diagnostic biomarker candidate. PloS one 9, e111039, https://doi.org/10.1371/journal.pone.0111039 (2014).
Frejo, L., Giegling, I., Teggi, R., Lopez-Escamez, J. A. & Rujescu, D. Genetics of vestibular disorders: pathophysiological insights. Journal of neurology 263(Suppl 1), 45–53, https://doi.org/10.1007/s00415-015-7988-9 (2016).
Frejo, L. et al. Extended phenotype and clinical subgroups in unilateral Meniere disease: A cross-sectional study with cluster analysis. Clinical otolaryngology: official journal of ENT-UK; official journal of Netherlands Society for Oto-Rhino-Laryngology & Cervico-Facial Surgery, https://doi.org/10.1111/coa.12844 (2017).
Duke, W. W. Ménière’s syndrome caused by allergy. Journal of the American Medical Association 81, 2179–2181, https://doi.org/10.1001/jama.1923.02650260021006 (1923).
Derebery, M. J. Allergic management of Meniere’s disease: an outcome study. Otolaryngology–head and neck surgery: official journal of American Academy of Otolaryngology-Head and Neck Surgery 122, 174–182 (2000).
Di Berardino, F. & Cesarani, A. Gluten sensitivity in Meniere’s disease. The Laryngoscope 122, 700–702, https://doi.org/10.1002/lary.22492 (2012).
Topuz, B., Ogmen, G., Ardic, F. N. & Kara, C. O. Provocation of endolymphatic hydrops with a prick test in Meniere’s disease. Advances in therapy 24, 819–825 (2007).
Keles, E. et al. Meniere’s disease and allergy: allergens and cytokines. The Journal of laryngology and otology 118, 688–693, https://doi.org/10.1258/0022215042244822 (2004).
Fuse, T. et al. Immunological responses in acute low-tone sensorineural hearing loss and Meniere’s disease. Acta oto-laryngologica 123, 26–31 (2003).
Derebery, M. J. Allergic and immunologic features of Meniere’s disease. Otolaryngologic clinics of North America 44, 655–666, ix, https://doi.org/10.1016/j.otc.2011.03.004 (2011).
Brookes, G. B. Circulating immune complexes in Meniere’s disease. Archives of otolaryngology–head & neck surgery 112, 536–540 (1986).
Gazquez, I. et al. MICA-STR A.4 is associated with slower hearing loss progression in patients with Meniere’s disease. Otology & neurotology: official publication of the American Otological Society, American Neurotology Society [and] European Academy of Otology and Neurotology 33, 223–229, https://doi.org/10.1097/MAO.0b013e31824296c8 (2012).
Requena, T. et al. Allelic variants in TLR10 gene may influence bilateral affectation and clinical course of Meniere’s disease. Immunogenetics 65, 345–355, https://doi.org/10.1007/s00251-013-0683-z (2013).
Cabrera, S. et al. Intronic variants in the NFKB1 gene may influence hearing forecast in patients with unilateral sensorineural hearing loss in Meniere’s disease. PloS one 9, e112171, https://doi.org/10.1371/journal.pone.0112171 (2014).
Frejo, L. et al. Clinical subgroups in bilateral Meniere disease. Frontiers in Neurology 7, https://doi.org/10.3389/fneur.2016.00182 (2016).
Ciccarelli, F., Martinis, M. D. & Ginaldi, L. An Update on Autoinflammatory Diseases. Current Medicinal Chemistry 21, 261–269, https://doi.org/10.2174/09298673113206660303 (2013).
Pathak, S., McDermott, M. F. & Savic, S. Autoinflammatory diseases: update on classification diagnosis and management. Journal of clinical pathology 70, 1–8, https://doi.org/10.1136/jclinpath-2016-203810 (2017).
Vambutas, A. & Pathak, S. AAO: Autoimmune and Autoinflammatory (Disease) in Otology: What is New in Immune-Mediated Hearing Loss. Laryngoscope investigative otolaryngology 1, 110–115, https://doi.org/10.1002/lio2.28 (2016).
Hurme, M. & Santtila, S. IL-1 receptor antagonist (IL-1Ra) plasma levels are co-ordinately regulated by both IL-1Ra and IL-1β genes. European Journal of Immunology 28, 2598–2602, https://doi.org/10.1002/(SICI)1521-4141(199808)28:08 2598::AID-IMMU2598 3.0.CO;2-K (1998).
Oosting, M. et al. Human TLR10 is an anti-inflammatory pattern-recognition receptor. Proceedings of the National Academy of Sciences of the United States of America 111, E4478–4484, https://doi.org/10.1073/pnas.1410293111 (2014).
Pathak, S., Hatam, L. J., Bonagura, V. & Vambutas, A. Innate immune recognition of molds and homology to the inner ear protein, cochlin, in patients with autoimmune inner ear disease. Journal of clinical immunology 33, 1204–1215, https://doi.org/10.1007/s10875-013-9926-x (2013).
Cembrzynska-Nowak, M. et al. TNF-alpha, IL-6 and IFN-gamma secreted by bronchoalveolar leukocytes isolated from patients with bronchial asthma, complicated by fungal airways infections. Archivum immunologiae et therapiae experimentalis 46, 381–386 (1998).
Rosenblum Lichtenstein, J. H. et al. Environmental Mold and Mycotoxin Exposures Elicit Specific Cytokine and Chemokine Responses. PloS one 10, e0126926, https://doi.org/10.1371/journal.pone.0126926 (2015).
Adrion, C. et al. Efficacy and safety of betahistine treatment in patients with Meniere’s disease: primary results of a long term, multicentre, double blind, randomised, placebo controlled, dose defining trial (BEMED trial). BMJ (Clinical research ed.) 352, h6816, https://doi.org/10.1136/bmj.h6816 (2016).
Vambutas, A. et al. Early efficacy trial of anakinra in corticosteroid-resistant autoimmune inner ear disease. The Journal of clinical investigation 124, 4115–4122, https://doi.org/10.1172/jci76503 (2014).
Helsinki, W. M. A. D. o. World Medical Association Declaration of Helsinki: ethical principles for medical research involving human subjects. Jama 310, 2191–2194, https://doi.org/10.1001/jama.2013.281053 (2013).
Satoh, H., Firestein, G. S., Billings, P. B., Harris, J. P. & Keithley, E. M. Tumor Necrosis Factor-α, an Initiator, and Etanercept, an Inhibitor of Cochlear Inflammation. The Laryngoscope 112, 1627–1634, https://doi.org/10.1097/00005537-200209000-00019 (2002).
Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical biochemistry 72, 248–254 (1976).
Acknowledgements
This work was supported by Grants from Meniere’s Society, UK and PI13/1242 Grant from ISCIII by FEDER Funds from the EU. Lidia Frejo was a PhD student oft he Biomedicine Program at the University of Granada. The authors would like to thank all patients and healthy controls participating in this study.We also would like to thank the Supercomputing and Bioinnovation Center (SCBI) from University of Málaga for the sharing of their resources (IPA®).
Author information
Authors and Affiliations
Contributions
J.A.L.E. study concept and design, study supervision and manuscript preparation. E.M.S., J.C.A.D., A.S.V., S.S.P., J.M.E.S., A.B.C., I.A., J.F., M.R.I., J.A.L.E. patient recruitment and clinical data acquisition. L.F. and T.R. P.B.M.C. isolation and mold stimulation, cytokines measurement, gene expression studies. L.F. and A.G.M. data acquisition, analysis and interpretation of data and manuscript preparation.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Frejo, L., Gallego-Martinez, A., Requena, T. et al. Proinflammatory cytokines and response to molds in mononuclear cells of patients with Meniere disease. Sci Rep 8, 5974 (2018). https://doi.org/10.1038/s41598-018-23911-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-23911-4
- Springer Nature Limited
This article is cited by
-
Cytokine profiling and transcriptomics in mononuclear cells define immune variants in Meniere Disease
Genes & Immunity (2024)
-
Serum/glucocorticoid-inducible kinase 1 deficiency induces NLRP3 inflammasome activation and autoinflammation of macrophages in a murine endolymphatic hydrops model
Nature Communications (2023)
-
Untargeted Metabolomic Analysis in Endolymphatic Sac Luminal Fluid from Patients with Meniere’s Disease
Journal of the Association for Research in Otolaryngology (2023)
-
Types of Inheritance and Genes Associated with Familial Meniere Disease
Journal of the Association for Research in Otolaryngology (2023)
-
Elevated G-CSF, IL8, and HGF in patients with definite Meniere’s disease may indicate the role of NET formation in triggering autoimmunity and autoinflammation
Scientific Reports (2022)