
Abstract
Soil respiration (Rs) is an important source of atmospheric CO2 flux and is sensitive to changes in soil nutrient and water contents. Despite extensive studies on the effects of enhanced atmospheric nitrogen (N) deposition and changes in precipitation (P) on Rs, few studies have taken into account the effects of interactions between these factors on Rs of alpine grasslands. To address these questions, we investigated the effects of N addition (10 g N m−2 yr−1), changes in precipitation (±50% precipitation), and their interaction on soil respiration and its components, including heterotrophic respiration (Rh) and autotrophic respiration (Ra),in a Tibetan alpine steppe during three consecutive growing seasons. We found that Rs differed in its response to N addition and precipitation regimes. Specifically, decreased precipitation led to a significant reduction in Rs during the last two years, whereas N addition minimally impacted Rs. Another important finding was that soil respiration components differed in their response to N addition and precipitation regimes. Nitrogen addition significantly enhanced Ra, whereas Rh was not altered in response to N addition. By contrast, the precipitation regime led to marked changes in Rh, but exhibited marginally significant effects on Ra. Therefore, our findings highlighted that soil respiration differed in its response to N addition and precipitation regimes mainly due to the different responses of soil respiration components to these factors. Therefore, carbon dynamics should take soil respiration components into account under global change scenarios.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
As the second-largest carbon (C) flux, soil respiration (Rs) plays a critical role in the regulation of C cycling in terrestrial ecosystems. Researchers have estimated that Rs accounts for 77 petagrams (Pg) of C released to the atmosphere each year1,2. Because grasslands account for approximately 37% of the world’s terrestrial region2, C releases from grassland soils significantly contribute to the global carbon cycle. Rs is composed of autotrophic respiration (Ra), which is related to roots, mycorrhizae and other rhizospheric microorganisms, and heterotrophic respiration (Rh) from non-rhizospheric microorganisms and free-living soil3,4,5. In the context of global changes, the response of Rsis mainly mediated by Ra and Rh related to biotic and abiotic factors directly or indirectly6,7. However, in recent years, as the importance of monitoring Ra and Rh been increasingly recognized, more and more experiments have started to measure Ra and Rh8,9. Numerous biotic and abiotic factors are assumed to regulate Rs and its components. N addition will likely affect Rs via impacts on these drivers10. Consequently, Ra and Rh are indispensable for evaluating the carbon balance of terrestrial ecosystems. However, we know little about how these factors directly and indirectly regulate Ra and Rh in the context of N addition.
Nitrogen (N) availability regulates global carbon balance11. Over recent decades, the burning of fossil fuel and use of fertilizer have increased the input of N into terrestrial ecosystems12,13, and this trend will continue to increase in the future14. The effects of N addition on Rs have been reported in different ecosystems, such as forest ecosystems15,16,17, temperate grasslands18,19,20,21 and other ecosystems. Nevertheless, the results are disputable and may be positive22,23, negative16 or neutral15,24,25. The reason for these differences may result from varied experimental duration and N addition rates. For instance, Ren found that N addition increased the Rs at an earlier stage but reduced Rs in a later period6. The effects of N availability on Rs were generally evaluated in previous studies, but the responses of these processes on the interaction of N additions and precipitation changes remain limited, particularly in this semi-arid grassland.
Compared with numerous studies examining the effects of N on Rs, our understanding of precipitation changes on Rs and its components is very limited, and information from the Qinghai-Tibetan Plateau is particularly lacking. More extreme precipitation events and drought are predicted to occur in the future26, and such changes will likely significantly affect the soil moisture and thus Rs. Nevertheless, compared with drought, few studies have evaluated the influence of heavy precipitation on Rs27,28,29. To date, the number of precipitation manipulation studies separating Rs components is very limited30. In addition, these studies simulated increased rainfall amounts through water addition, which may not necessarily reflect the real conditions of natural extreme rainfall events that may exceed past and current variation ranges31,32,33. Knapp developed a conceptual model called the Bucket Model to predict the response of terrestrial ecosystems to increased intra-annual precipitation variability characterized by extreme precipitation events and longer dry intervals29. The Bucket Model predicts that the primary productivity and Rs in arid lands is increased by the increased precipitation variability34. To date, it remains unknown how precipitation changes affect soil respiration in cold and dry environments, such as the Tibetan alpine steppe.
As the largest and highest plateau in the world, Qinghai-Tibetan Plateau has a mean elevation of greater than 4000 m and an area of 2.0 × 106 km2,35 and is one of the most sensitive areas to climate change. As the dominant ecosystems on the plateau, alpine steppe and alpine meadow occupied greater than 60% of the total region. Atmospheric N deposition is significant in the eastern Plateau due to regional economic development, ranging from 8.7 to 13.8 kg N ha−1 yr−1 36. Unlike N deposition, the precipitation pattern varied across the Qinghai-Tibetan Plateau, which increased in the northern area and decreased in the southern area37. Nevertheless, few studies have concentrated on the influence of precipitation changes and N addition on Rs in alpine steppe.
In this study, we conducted a field precipitation and N manipulative experiment in an alpine steppe, and we aimed to investigate the responses of soil respiration and its components to precipitation changes and N additions. Our objectives were to (i) reveal the response patterns of Rs and its constituents to N addition and precipitation changes, and (ii) evaluate the impacts of biotic and abiotic variables on Ra and Rh through N addition and precipitation changes.
Results
Seasonal variations in Microclimates
Figure 1 presents the seasonal patterns of soil temperature, soil water content and precipitation from September 2014 to October 2016. The ten-day mean soil temperature ranged from −7.67 to 18.20 °C at a 0- to 10-cm soil depth (Fig. 1a), and mean soil moisture ranged from 7.93 to 26.97 V/V% at a 0- to 10-cm soil depth (Fig. 1b). The 10-day precipitation ranged from 0 to 128.8 mm. During the growing season, the total precipitation was 502.5 mm in 2014, 367 mm in 2015 and 473.4 mm in 2016 (Fig. 1c).

Seasonal and annual variation in (a) soil temperature at a depth of 5 cm; (b) volumetric soil moisture at a 5-cm depth; (c) precipitation from May in 2014 to October in 2016. Error bars indicate the standard errors.
Soil temperature and moisture
Given control of soil moisture and soil temperature over soil respiration, we examined the dynamic changes in soil temperature and soil moisture in the growing season. Our results demonstrated that N addition significantly reduced soil temperature rather than soil moisture (Table 1). By contrast, increased precipitation led to a significant increase in soil moisture, whereas reduced precipitation significantly reduced soil moisture (Table 1). However, precipitation did not affect soil temperature (Table 1). Moreover, we found that N- and precipitation-induced changes in soil respiration were closely associated with soil temperature and moisture (Fig. 2).

Relationships of Rs with seasonal mean soil temperature and moisture across the 30 plots. Black solid lines represent regression curves.
Effects of N addition and precipitation change on soil nitrogen availability
Consecutive N addition significantly enhanced soil N availability. For example, NH4+-N and NO3-N concentrations in soil were enhanced by 49.01% and 164.89%, respectively. However, NH4+-N and NO3-N concentrations in soil exhibited differential responses to precipitation. Specifically, precipitation had a significant effect on NO3-N concentrations in soil but no impact on NH4+-N concentrations in soil (Table 2). For example, decreased precipitation enhanced soil NO3-N concentrations by 38.01%, whereas enhanced precipitation reduced NO3-N concentrations by 20.16% (Table 2). Moreover, the stimulatory effects of precipitation on NO3-N concentration were much stronger in 2015 compared with 2016 (Table 2).
Effects of N addition and precipitation change on aboveground and below-ground biomass
Above-ground biomass (AGB) of the community was significantly enhanced by N addition, whereas N addition had no effect on below-ground biomass (BGB). Similar to N addition, decreased precipitation significantly reduced AGB. However, decreased precipitation marginally reduced BGB, whereas enhanced precipitation had no effect on BGB (Fig. 3).

Effects of nitrogen and precipitation changes on (a) aboveground biomass (AGB), (b) belowground biomass (BGB) in 2016. N1 indicates CK, nitrogen addition (N2), 50% reduction in precipitation (P1), natural precipitation (P2), 50% additional precipitation (P3), ′P < 0.1, *P < 0.05, ***P < 0.001.Error bars indicate the standard errors.
Soil respiration and its Constitute during the growing season
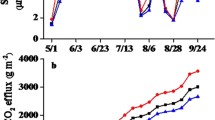
The soil respiration rate exhibited a unimodal pattern in the growing season. The pattern was consistent in the consecutive 3-year observations from different treatments (Fig. 4). For example, soil respiration peaked in July to August and subsequently declined thereafter (Fig. 4). Notably, the peak of soil respiration in 2015 was much greater than that in 2014 and 2016 (Fig. 4). Moreover, N addition and precipitation exhibited differential effects on soil respiration. For example, consecutive N addition had no effects on soil respiration (Fig. 5). By contrast, increased precipitation significantly increased soil respiration, where as reduced precipitation led to a significant reduction in soil respiration (Fig. 5).

Seasonal variation in soil respiration rates under different treatments in (a) 2014; (b) 2015; (c) 2016. N1P1 indicates the 50% precipitation reduction treatment, natural precipitation (N1P2), 50% additional precipitation treatment (N1P3), 50% reduction in precipitation with nitrogen addition treatment (N2P1), natural precipitation without nitrogen addition (N2P2), 50% additional precipitation with nitrogen addition (N2P3). Error bars indicate the standard errors.

Effects of precipitation changes on mean soil respiration during the growing seasons from 2014 to 2016. P1 indicates 50% reduction in precipitation, natural precipitation (P2), 50% additional precipitation (P3). Error bars indicate the standard errors.
In the growing seasons, precipitation changes and N addition were observed from 2014 to 2016. N addition (N2) enhanced Rs by 4.16% in 2014 and reduced Rs by 0.19% and 1.13% in 2015 and 2016, respectively; however, these effects were not significant (Table 3; Fig. 6a,b). Similarly, the effect of P3 treatment on Rs was significant (Table 3; Fig. 6e,f). Compared with the P2 treatments, P1 treatments significantly decreased Rs by 17.5% (P = 0.082) in 2016, and in no significant effects were noted in 2014 and 2015 (Table 3; Fig. 6c,d).

Effects of precipitation changes and N addition on soil respiration (a,c,e) and changes (%) in soil respiration (b,d,f) during the growing seasons from 2014 to 2016. N1 indicates CK, nitrogen addition (N2), 50% reduction in precipitation (P1), natural precipitation (P2), 50% additional precipitation (P3), ′P < 0.1, *P < 0.05, ***P < 0.001, Error bars indicate the standard errors.
To further investigate the effects of N addition and precipitation on soil respiration components, we monitored autotrophic and heterotrophic respiration. Our results demonstrated that the two types of soil respiration differed in their responses to N addition. Specifically, N addition significantly enhanced autotrophic respiration but had no effects on heterotrophic respiration (Fig. 7a,b; Table 4). Moreover, despite the lack of response of autotrophic respiration to precipitation, decreased precipitation led to a significant reduction in heterotrophic respiration. In particular, in 2016, decreased precipitation had minimal impacts on autotrophic respiration (Fig. 7c,d; Table 4). In addition, only the contribution of Rh to the total Rs was reduced by N2 (Rh/Rs, by 6.85%, Table 5).

Effects of nitrogen addition and precipitation changes on (a) autotrophic respiration (Ra) in 2015, (b) autotrophic respiration (Ra) in 2016, (c) heterotrophic respiration (Rh) in 2015 and (d) heterotrophic respiration (Rh) in 2016. N1 indicates CK, nitrogen addition (N2), 50% precipitation reduction (P1), natural precipitation (P2), 50% additional precipitation (P3), ′P < 0.1, *P < 0.05, *** P < 0.001. Error bars indicate the standard errors.
Discussion
In the present study, we found that the addition of and altered precipitation had differential effects on soil respiration in a Tibetan alpine steppe plateau (Fig. 6). Soil respiration in the growing season exhibited no response to consecutive N addition for three years (Fig. 6a,b). By contrast, increased precipitation led to a significant increase in soil respiration, whereas reduced precipitation significantly reduced soil respiration (Fig. 6 and Table 3), suggesting that changes in soil respiration in this ecosystem might be driven by precipitation limitations.
Effects of N addition
A recent meta-analysis demonstrated that N addition, increased Rs and Ra but reduced Rh in grasslands22. However, we found that the response of soil respiration to N addition was not significant (Table 3; Fig. 6a,b), and these results were similar to previous studies24,38,39. Although the effects of N addition are not significant, a slight trend is observed in the first year wherein the addition of N increased Rs. However, Rs values during the subsequent two years were not significantly altered or decreased (Fig. 4a,b). Similar temporal responses were observed in an alpine meadow, in which N addition increased Rs during the first year, had no effect during the following two years, and decreased Rs rates in the last year6,40,41. This finding suggests that the response of soil respiration to N addition is strongly dependent on the duration of N addition. These results are consistent with recent investigations in the same region10. Of note, our study only presented the responses of Rs to short-term N additions (three years), but it remains undetermined whether long-term N enrichment would yield some differences. Therefore, experiments with a longer duration are required to better understand Rs and Ra/Rh relationships in future studies.
Rs was not altered by N addition (Fig. 6a,b). However, another important finding is that Ra was significantly increased by N addition (Table 4). The Ra response was similar to the response of aboveground biomass and belowground biomass (Fig. 2). Belowground biomass had a direct effect on Ra because larger root biomass represents a larger root surface area for respiration10,22. Similar responses to N addition have been observed in a meta-analysis by Xia & Wan42. The meta-analysis demonstrated that total root biomass increased by 23%, suggesting that N addition increased biomass. Greater root biomass indicates that more C was allocated below ground43, which is typically associated with increased root respiration44. However, upon continual nitrogen input into alpine steppe ecosystems, nitrogen saturation will ultimately be achieved, which will cause the root biomass to plateau10,22. We observed that N addition did not significant increase BGB in 2016. However, a recent meta-analysis found that the addition of N significantly increased BGB on the Tibetan Plateau45. Although the BGB was not significantly altered, we observed an increasing trend for N addition in 2016 in this study (17.01%; Fig. 2a,b). The different findings may be attributed to different soil temperature (Table 1), the soil temperature in our experiment was significantly decreased by N addition. For vegetation growth, temperature is a limiting factor in the alpine steppe on the Tibetan Plateau, and thus higher temperature may stimulate vegetation productivity46. Moreover, the soil’s inorganic N concentration increased with the addition of N (Table 2). The high inorganic N accumulated in 0–10 cm of soil may be toxic to microbial activities and growth47.
A previous study in an alpine steppe indicated that both plant growth and microbial activity were limited by N48. However, long-term N input to N-limited soils should initially stimulate soil microbial activity and lead to carbon-limited conditions after the microbial requirements for N are achieved47. This limitation of C to microorganisms under conditions of supplementation with N would lead to reductions in Rh and inhibition of microbial activity25,49. Furthermore, we observed that soil temperature exhibited a decreasing trend with increasing N additions (Table 1). Soil temperature not only directly explained the variations in Ra and Rh by adjusting respiration enzymes50 but also indirectly influenced these values via affecting microbial and root growth16. These effects were particularly noted in cold areas where temperature is generally considered a limiting factor for living things51. However, a recent meta-analysis found that N addition did not significantly affect soil temperature on the Tibetan Plateau45. This finding is potentially explained by the fact that the addition of N increased aboveground plant growth, and a more closed canopy prevented the incoming solar radiation from reaching the soil surface52. Moreover, we observed a significant correlation between soil respiration and soil moisture/temperature (Fig. 2). To our knowledge, soil moisture is mainly mediated by soil temperature53. For example, increased soil temperature often reduces soil moisture53. However, N-induced reductions in soil temperature rarely reduce soil moisture. Therefore, soil respiration in the Tibetan alpine steppe plateau was regulated by soil moisture. This may represent an important mechanism by which N addition and altered precipitation affect soil respiration.
Effects of altered precipitation
Our findings provide insights into the effects of precipitation changes on Rs in an alpine ecosystem. We found that soil respiration in an alpine steppe exhibited a drastic response to reduced precipitation and a mild response to increased precipitation. Previous studies have suggested that the direction of the Rs response was likely influenced by the water status of an ecosystem with either reductions or increases in precipitation54. Our results supported the hypothesis that altered precipitation would change Rs29, but differential responses were found for different treatments. In addition, the carbon cycles in arid and semi-arid regions are more sensitive to changes in precipitation55,56. In an alpine steppe ecosystem that lacks precipitation, precipitation becomes more important. However, in this study, interestingly, Rs in an alpine steppe ecosystem exhibited a drastic response to precipitation decreases in the Qinghai-Tibetan Plateau.
With the increasing frequency of extreme climate conditions, the mechanism by which drought affects terrestrial carbon cycling has received great attention57. Compared with the results of several previous studies in other arid ecosystems58,59, reducing precipitation significantly decreased Rs and its component (Ra and Rh) in an alpine steppes (Fig. 4c,d) in our study. Considering the close relationship between Ra and plant photosynthetic activity60, the reduced Ra could be attributed to the inhibition of plant growth, which may supply less C substrate for root/rhizospheric respiration61,62. Similar to N addition, a larger root biomass means a greater root surface area for respiration10,22. Moreover, soil moisture impacts the sensitivity of plant growth, which reduces Ra in this region. The response to soil moisture suggests that decreased precipitation can largely restrict soil respiration and its components, which is more significant if the infrequency of precipitation and number of drought events due to climatic changes were increased63,64,65.
Compared with Ra, decreased precipitation markedly affected Rh, with cumulative Rh reduced in drought conditions. The reduction in Rh could be partly due to the reduction in microbial biomass. A previous study found that decreasing precipitation significantly reduced microbial biomass in a similar grassland community66. In addition, reduced precipitation treatment strongly decreased Rh, and drought significantly suppressed Rh. The results conducted in a pine forest are consistent with those of a girdling experiment67. Other studies have found that Rs (including Rh) is sensitive to soil moisture in old field systems68. Soil water content can change Rh by altering the activity of decomposer microbes and substrate availability69. Decreases in microbial biomass and activities under conditions of reduced water availability have been reported in different ecosystems70. Therefore, we concluded that the reduced precipitation treatment constrained microbial activity and plant growth, subsequently reducing both Ra and Rh.
Compared with reduced precipitation, the response of Rs and its component to increasing precipitation is not significant (Table 2, Fig. 4e,f). Then, an interesting question arises. Why is Rs not altered under conditions of increased precipitation? We explored several potential reasons that could explain the lack of response of Rs to increased precipitation. First, our study only reported the responses of Rs to short-term increased precipitation (three years), but it remains unclear whether long-term precipitation increased would cause some differences. Second, soil respiration is significantly related to AGB in alpine grasslands on the Tibetan Plateau71. Thus, the non-significant response of AGB (Fig. 2a) to increased precipitation may also explain the non-significant response of soil respiration to increased precipitation. Third, the precipitation distribution varied significantly during the last two growing seasons (Fig. 1c). In particular, for the peak plant growth in August, the precipitation amount in 2015 (79.1 mm) was significantly reduced compared with that in 2016 (200.2 mm). The response of AGB and soil moisture to increased precipitation depends on the amount of increased precipitation in alpine grasslands on the Tibetan Plateau72 given that both AGB and soil moisture can affect soil respiration73. Antecedent precipitation and soil moisture conditions before Rs measurements can also affect the response of soil respiration to precipitation, and these factors also might result in non-significant changes in soil respiration to precipitation74. Moreover, the non-significant response of Rs to increased precipitation may also be related to the non-significant difference in soil respiration temperature sensitivity between the control and increased precipitation treatments74. Moreover, the highly inorganic N that accumulated in 0–10 cm of soil may be toxic to microbial growth and activities47, and we found that soil NO3−N decreased in the wet year (2016). In addition, high soil moisture potentially stimulated root and microbial activities52, and additional precipitation could increase Ra due to enhanced plant growth (Fig. 2a,b), thus leading to non-significant changes in Rs during this period. Finally, the positive effect of nitrogen addition on AGB and nitrate nitrogen increases with increasing the nitrogen addition rate45,75. Thus, the non-significant response of soil respiration to nitrogen addition may also be related to its relatively low nitrogen addition rate. Based on these points, it is possible that precipitation-associated water increases collectively remain very important in this alpine steppe. Experiments with longer durations are necessary to better understand Rs and Ra/Rh relationships in future studies. Our findings suggest that Rs might be altered based on precipitation changes in alpine steppe ecosystems in the future.
Increasing precipitation could increase N2O76, N addition could also increase N2O77. N2O emissions from soils are caused principally by microbial nitrification and denitrification by soil water-filled pore space, mineral nitrogen concentration, temperature and precipitation76. Moreover, both CO2 and N2O as two major greenhouse gases could render global warming. Therefore, more attention should be paid to the N2O in future studies.
Conclusions
Overall, both N addition and altered precipitation did not significantly influence Rs in the first year, but reduced precipitation drastically repressed Rs during the subsequent years. Furthermore, we found that the addition of N significantly increased Ra but had no obvious effects on Rh during the last two years. In addition, altered precipitation significantly influenced Rh but minimally affected Ra. Our results revealed the distinct effects of the addition of N and altered precipitation regimes on heterotrophic and autotrophic respiration. Soil respiration is regulated by soil moisture rather than soil temperature in the Tibetan alpine steppe. These findings suggest that the response of Rs to climate change may be more complex, and our ability to predict the C-N-precipitation interactions will become more challenging in Tibetan alpine steppe ecosystems. In addition, the changes in Rs and its components in response to N addition and precipitation changes merit further attention in future experimental and modelling analyses.
Methods
Study site
The study was conducted on the Sanjiaocheng Sheep Breeding Farm (37°18′N, 100°15′E) located in Qinghai province, China. The elevation is 3,286 m. The long-term mean annual precipitation is 387 mm, and the mean annual temperature is 0.08 °C (1980–2012). The soil is classified as chestnut soil78 with 61.0% sand, 33.4% silt, and 4.9% clay. The 0- to 30-cm soil properties are as follows: pH 9.5, total organic carbon 20.5 g kg−1, total nitrogen 2.5 g kg−1 and total phosphorous 0.6 g kg−1.
The plant community is dominated by Stipapurpurea, Poa crymophila, Artemisia scoparia, and Carexivanovaeat at the experimental site. Abundant species include Heteropappusaltaicus, Koeleriaglauca, Agropyroncristatum, Leymussecalinus, Dracocephalumheterophyllum, Taraxacummongolicum, Cirsiumjaponicum, Potentilla multifida, and Pedicularis alaschanica.
Experimental design
The experimental area was fenced to prevent grazing disturbances, and the experiment was started in May 2013. Thirty plots of 3.3 × 2.7 m2 were randomly assigned to 6 treatments within 5 blocks using a randomized block design. Each plot was cut 15 cm around the border with iron sheets (3.0 × 2.4 m). To prevent surface runoff, we inserted an algam into soil at a depth of 20 cm in each plot. Blocks and plots were separated by a 2-m buffer zone. Each plot was split into 3 subplots (subplot 1 for measuring plant biomass and soil characteristics, subplot 2 for measuring carbon fluxes and subplot 3for observing the phenology).
For N treatments, NH4NO3 (10 g N m−2 y−1) was evenly sprayed using a sprayer with 1 litre of water into the plots on June and July 15th during each year from 2013 to 2016, and the same amount of water (1 L) was evenly distributed in the control treatments. Previous studies using similar methods to add N indicated that the saturated N load was 8 g m−2 year−1 at same region in a Tibetan alpine steppe79, 8–10 g m−2 year−1 in temperate grasslands in Inner Mongolia80. Thus, the current N addition should be adequate to simulate N saturation in our study site. Precipitation treatments were performed using a steel structure bracket and sunlight-penetrable concave polyvinyl chloride (PVC) boards (1 mm). The steel structure is inclined (north 2.1 m and south 1.4 m), and five PVC boards (each PVC is 24 cm × 352 cm) were alternately arranged with five equal and empty areas, which occupied 50% of the total area. Compared with 50% precipitation decreased treatment, both the contrast treatment and 50% precipitation increased treatment are cut open at the bottom of the PVC boards. Rainwater evenly distributed by a sprayer into the 50% precipitation increased treatment which collected in 50% precipitation decreased treatment through PVC pipes. In addition, previous studies also used ±50% precipitation changes in same region in a Tibetan alpine steppe81. Six treatments were included in this study: 50% precipitation reduction treatment (N1P1), natural precipitation (N1P2), 50% additional precipitation treatment (N1P3), 50% reduced precipitation with N addition treatment (N2P1), natural precipitation without N addition (N2P2), and 50% additional precipitation with N addition (N2P3).The six treatments are described in Supplement Table 1.
Soil respiration measurements
In June 2013, two polyvinyl chloride collars (20 cm in diameter) were inserted into the soil at least 30 cm away from the edge. In each plot, the Rs collar was inserted into the soil to a depth of 3 cm, and the Rh collar was inserted into the soil at least 50 cm. Before measurements, we removed living plants inside the collars once a week. During the growing season (May-October), Rs and Rh were measured with a portable soil CO2 flux system (Li-8100, Li-COR, Inc., Lincoln, NE, USA) each month between 9:00 and 12:00 am from 2014 to 2016. Ra was calculated as Rs minus Rh6. During the CO2 measurements, the soil temperature and volumetric moisture at a 0–10 soil depth were monitored adjacent to the collar using a digital thermometer and a portable TDR-100 soil moisture probe (Spectrum Technologies, Inc., Plainfield, IL, USA), respectively. In addition, soil temperature and moisture were continuously recorded using an EM-50 (Decagon, USA) at a depth of 5 cm every 15 min in each plot.
Above- and belowground sampling
In this study, aboveground and belowground biomasses were collected during peak growth in the middle of August in 2016. We randomly selected three 0.25 m × 0.25 m quadrants and harvested each quadrant in subplot 1. All plant samples were oven-dried at 65 °C to a constant mass and then weighed to estimate aboveground biomass (AGB).
After AGB sampling, three root augers (4 cm in diameter) were collected in the same quadrants to a depth of 30 cm (10 cm intervals). Then, three cores were mixed as one sample and soaked in water using a 0.5-mm sieve to remove the soil. After BGB presented, root mass was measured and oven dried at 65 °C.
Soil sampling and inorganic N analyses
Soil samples were collected in the middle of August in each experimental year. Within each plot, soil cores (3 cm in diameter) were collected from the surface layer (0–10 cm) and combined to yield one composite sample. Soil samples were passed through a 2-mm screen in the field after the organic debris and roots were removed, stored in a cooler and transported to the laboratory for the following analyses. The soil’s inorganic N (NO3−-N and NH4+-N) was extracted with a 2 M KCl solution and analysed using a AQ1 Discrete Multi-Chemistry Analyzer (SEAL Analytical, USA).
Statistical analysis
In this study, three-way ANOVA was used to compare the treatment differences based on year, N, precipitation, total NH4+-N and NO3-N. Repeated measures analysis of variance (ANOVA) was used to examine the effects of sampling date, precipitation and non-soil temperature, soil moisture, Rs and its components (Ra and Rh) for the three years as well as their contribution to total Rs (Ra/Rs, Rh/Rs) in 2015 and 2016. All statistical analyses were conducted using SPSS 16.0 (IBM SPSS, Inc., Chicago, Illinois, USA). Unless otherwise stated, the differences were considered statistically significant at P < 0.05.
References
O’Mara, F. P. The role of grasslands in food security and climate change. Annals of botany 110, 1263–1270 (2012).
Raich, J. W. & Potter, C. S. Global patterns of carbon dioxide emissions from soils. Global Biogeochemical Cycles 9, 23–36 (1995).
Sun, Z. et al. The effect of nitrogen addition on soil respiration from a nitrogen-limited forest soil. Agricultural and forest meteorology 197, 103–110 (2014).
Hanson, P., Edwards, N., Garten, C. T. & Andrews, J. Separating root and soil microbial contributions to soil respiration: a review of methods and observations. Biogeochemistry 48, 115–146 (2000).
Bond‐Lamberty, B., Wang, C. & Gower, S. T. A global relationship between the heterotrophic and autotrophic components of soil respiration? Global Change Biology 10, 1756–1766 (2004).
Ren, F. et al. Contrasting effects of nitrogen and phosphorus addition on soil respiration in an alpine grassland on the Qinghai-Tibetan Plateau. Scientific reports 6 (2016).
Wang, X. et al. Soil respiration under climate warming: differential response of heterotrophic and autotrophic respiration. Global change biology 20, 3229–3237 (2014).
Li, D., Zhou, X., Wu, L., Zhou, J. & Luo, Y. Contrasting responses of heterotrophic and autotrophic respiration to experimental warming in a winter annual‐dominated prairie. Global change biology 19, 3553–3564 (2013).
Suseela, V. & Dukes, J. S. The responses of soil and rhizosphere respiration to simulated climatic changes vary by season. Ecology 94, 403–413 (2013).
Peng, Y. et al. Nonlinear response of soil respiration to increasing nitrogen additions in a Tibetan alpine steppe. Environmental Research Letters 12, 024018 (2017).
Fernández-Martínez, M. et al. Nutrient availability as the key regulator of global forest carbon balance. Nature Climate Change 4, 471–476 (2014).
Reay, D. S., Dentener, F., Smith, P., Grace, J. & Feely, R. A. Global nitrogen deposition and carbon sinks. Nature Geoscience 1, 430–437 (2008).
Penuelas, J. et al. Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe. Nature communications 4, 2934 (2013).
Galloway, J. N. et al. Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science 320, 889–892 (2008).
Allison, S. D., Czimczik, C. I. & Treseder, K. K. Microbial activity and soil respiration under nitrogen addition in Alaskan boreal forest. Global Change Biology 14, 1156–1168 (2008).
Janssens, I. et al. Reduction of forest soil respiration in response to nitrogen deposition. Nature Geoscience 3, 315–322 (2010).
Mo, J. et al. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Global Change Biology 14, 403–412 (2008).
Zhu, C. et al. Divergent effects of nitrogen addition on soil respiration in a semiarid grassland. Scientific reports 6, 33541 (2016).
Yan, L., Chen, S., Huang, J. & Lin, G. Differential responses of auto‐and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe. Global Change Biology 16, 2345–2357 (2010).
Liu, W., Xu, W., Han, Y., Wang, C. & Wan, S. Responses of microbial biomass and respiration of soil to topography, burning, and nitrogen fertilization in a temperate steppe. Biology and fertility of soils 44, 259–268 (2007).
Peng, Q. et al. Effects of nitrogen fertilization on soil respiration in temperate grassland in Inner Mongolia, China. Environmental Earth Sciences 62, 1163–1171 (2011).
Zhou, L. et al. Different responses of soil respiration and its components to nitrogen addition among biomes: a meta‐analysis. Global change biology 20, 2332–2343 (2014).
Verburg, P. S. et al. Net ecosystem carbon exchange in two experimental grassland ecosystems. Global Change Biology 10, 498–508 (2004).
Yue, K. et al. Stimulation of terrestrial ecosystem carbon storage by nitrogen addition: a meta-analysis. Scientific reports 6, 19895 (2016).
Liu, L. & Greaver, T. L. A global perspective on belowground carbon dynamics under nitrogen enrichment. Ecology Letters 13, 819–828 (2010).
Solomon, S. Climate change 2007-the physical science basis: Working group I contribution to the fourth assessment report of the IPCC. Vol. 4 (Cambridge University Press, 2007).
Harper, C. W., Blair, J. M., Fay, P. A., Knapp, A. K. & Carlisle, J. D. Increased rainfall variability and reduced rainfall amount decreases soil CO2 flux in a grassland ecosystem. Global Change Biology 11, 322–334 (2005).
Zhou, X., Sherry, R. A., An, Y., Wallace, L. L. & Luo, Y. Main and interactive effects of warming, clipping, and doubled precipitation on soil CO2 efflux in a grassland ecosystem. Global Biogeochemical Cycles 20 (2006).
Knapp, A. K. et al. Consequences of more extreme precipitation regimes for terrestrial ecosystems. AIBS Bulletin 58, 811–821 (2008).
Liu, L. et al. A cross‐biome synthesis of soil respiration and its determinants under simulated precipitation changes. Global change biology 22, 1394–1405 (2016).
Chen, Z. et al. Extreme rainfall and snowfall alter responses of soil respiration to nitrogen fertilization: a 3‐year field experiment. Global change biology (2017).
Beier, C. et al. Precipitation manipulation experiments–challenges and recommendations for the future. Ecology Letters 15, 899–911 (2012).
Kayler, Z. E. et al. Experiments to confront the environmental extremes of climate change. Frontiers in Ecology and the Environment 13, 219–225 (2015).
Thomey, M. L. et al. Effect of precipitation variability on net primary production and soil respiration in a Chihuahuan Desert grassland. Global Change Biology 17, 1505–1515 (2011).
Li, W. & Zhou, X. Ecosystems of Qinghai-Xizang (Tibetan) Plateau and approach for their sustainable management. Guangdong Science and Technology Press, Guangzhou, 56–101 (1998).
Lü, C. & Tian, H. Spatial and temporal patterns of nitrogen deposition in China: synthesis of observational data. Journal of Geophysical Research: Atmospheres 112 (2007).
Duan, K., Yao, T., Wang, N., Tian, L. & Xu, B. The difference in precipitation variability between the North and South Tibetan Plateaus. J Glaciol Geocryol 30, 726–732 (2008).
Micks, P., Aber, J. D., Boone, R. D. & Davidson, E. A. Short-term soil respiration and nitrogen immobilization response to nitrogen applications in control and nitrogen-enriched temperate forests. Forest Ecology and Management 196, 57–70 (2004).
Ambus, P. & Robertson, G. The effect of increased N deposition on nitrous oxide, methane and carbon dioxide fluxes from unmanaged forest and grassland communities in Michigan. Biogeochemistry 79, 315–337 (2006).
Han, Y., Zhang, Z., Wang, C., Jiang, F. & Xia, J. Effects of mowing and nitrogen addition on soil respiration in three patches in an oldfield grassland in Inner Mongolia. Journal of plant ecology 5, 219–228 (2011).
Xu, W. & Wan, S. Water-and plant-mediated responses of soil respiration to topography, fire, and nitrogen fertilization in a semiarid grassland in northern China. Soil Biology and Biochemistry 40, 679–687 (2008).
Xia, J. & Wan, S. Global response patterns of terrestrial plant species to nitrogen addition. New Phytologist 179, 428–439 (2008).
Peng, Y. & Yang, Y. Allometric biomass partitioning under nitrogen enrichment: Evidence from manipulative experiments around the world. Scientific reports 6 (2016).
Bai, W. et al. Increased temperature and precipitation interact to affect root production, mortality, and turnover in a temperate steppe: implications for ecosystem C cycling. Global change biology 16, 1306–1316 (2010).
Fu, G. & Shen, Z.-X. Response of alpine plants to nitrogen addition on the Tibetan Plateau: A meta-analysis. Journal of plant growth regulation 35, 974–979 (2016).
Piao, S., Fang, J. & He, J. Variations in vegetation net primary production in the Qinghai-Xizang Plateau, China, from 1982 to 1999. Climatic Change 74, 253–267 (2006).
Aber, J. D., Nadelhoffer, K. J., Steudler, P. & Melillo, J. M. Nitrogen saturation in northern forest ecosystems. BioScience 39, 378–286 (1989).
Jiang, C. et al. Nutrient resorption of coexistence species in alpine meadow of the Qinghai-Tibetan Plateau explains plant adaptation to nutrient-poor environment. Ecological Engineering 44, 1–9 (2012).
Li, Y., Liu, Y., Wu, S., Niu, L. & Tian, Y. Microbial properties explain temporal variation in soil respiration in a grassland subjected to nitrogen addition. Scientific reports 5, 18496 (2015).
Atkin, O. K. & Tjoelker, M. G. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends in plant science 8, 343–351 (2003).
Lloyd, J. & Taylor, J. On the temperature dependence of soil respiration. Functional ecology, 315–323 (1994).
Yiqi, L. & Zhou, X. Soil respiration and the environment. (Academic press, 2010).
Suseela, V., Conant, R. T., Wallenstein, M. D. & Dukes, J. S. Effects of soil moisture on the temperature sensitivity of heterotrophic respiration vary seasonally in an old‐field climate change experiment. Global Change Biology 18, 336–348 (2012).
Sowerby, A., Emmett, B. A., Tietema, A. & Beier, C. Contrasting effects of repeated summer drought on soil carbon efflux in hydric and mesic heathland soils. Global Change Biology 14, 2388–2404 (2008).
Knapp, A. K. & Smith, M. D. Variation among biomes in temporal dynamics of aboveground primary production. Science 291, 481–484 (2001).
Flanagan, L. B., Wever, L. A. & Carlson, P. J. Seasonal and interannual variation in carbon dioxide exchange and carbon balance in a northern temperate grassland. Global Change Biology 8, 599–615 (2002).
Knapp, A. K. et al. Characterizing differences in precipitation regimes of extreme wet and dry years: implications for climate change experiments. Global change biology 21, 2624–2633 (2015).
Mariko, S., Urano, T. & Asanuma, J. Effects of irrigation on CO 2 and CH 4 fluxes from Mongolian steppe soil. Journal of Hydrology 333, 118–123 (2007).
Patrick, L. et al. Effects of an increase in summer precipitation on leaf, soil, and ecosystem fluxes of CO2 and H2O in a sotol grassland in Big Bend National Park, Texas. Oecologia 151, 704–718 (2007).
Wan, S. & Luo, Y. Substrate regulation of soil respiration in a tallgrass prairie: results of a clipping and shading experiment. Global biogeochemical cycles 17 (2003).
HoÈgberg, P. et al. Large-scale forest girdling shows that current photosynthesis drives soil respiration. Nature 411, 789–792 (2001).
Kuzyakov, Y. & Cheng, W. Photosynthesis controls of rhizosphere respiration and organic matter decomposition. Soil Biology and Biochemistry 33, 1915–1925 (2001).
Cook, E. R., Woodhouse, C. A., Eakin, C. M., Meko, D. M. & Stahle, D. W. Long-term aridity changes in the western United States. Science 306, 1015–1018 (2004).
Manabe, S., Wetherald, R. T., Milly, P., Delworth, T. L. & Stouffer, R. J. Century-scale change in water availability: CO 2-quadrupling experiment. Climatic change 64, 59–76 (2004).
Dore, M. H. Climate change and changes in global precipitation patterns: what do we know? Environment international 31, 1167–1181 (2005).
Liu, W., Zhang, Z. & Wan, S. Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland. Global Change Biology 15, 184–195 (2009).
Scott‐Denton, L. E., Rosenstiel, T. N. & Monson, R. K. Differential controls by climate and substrate over the heterotrophic and rhizospheric components of soil respiration. Global Change Biology 12, 205–216 (2006).
Wan, S., Norby, R. J., Ledford, J. & Weltzin, J. F. Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old‐field grassland. Global Change Biology 13, 2411–2424 (2007).
Williams, M. A. Response of microbial communities to water stress in irrigated and drought-prone tallgrass prairie soils. Soil Biology and Biochemistry 39, 2750–2757 (2007).
Fierer, N., Schimel, J. P. & Holden, P. A. Variations in microbial community composition through two soil depth profiles. Soil Biology and Biochemistry 35, 167–176 (2003).
Fu, G., Zhang, X.-Z., Zhou, Y.-T., Yu, C.-Q. & Shen, Z.-X. Partitioning sources of ecosystem and soil respiration in an alpine meadow of Tibet Plateau using regression method. Polish Journal of Ecology 62, 17–24 (2014).
Fu, G., Shen, Z.-X. & Zhang, X.-Z. Increased precipitation has stronger effects on plant production of an alpine meadow than does experimental warming in the Northern Tibetan Plateau. Agricultural and Forest Meteorology 249, 11–21 (2018).
Shen, Z.-X. et al. The soil drying along the increase of warming masks the relation between temperature and soil respiration in an alpine meadow of Northern Tibet. Polish Journal of Ecology 64, 125–129 (2016).
Shen, Z.-X., Li, Y.-L. & Fu, G. Response of soil respiration to short-term experimental warming and precipitation pulses over the growing season in an alpine meadow on the Northern Tibet. Applied Soil Ecology 90, 35–40 (2015).
Fu, G. & Shen, Z.-X. Response of alpine soils to nitrogen addition on the Tibetan Plateau: A meta-analysis. Applied Soil Ecology 114, 99–104 (2017).
Dobbie, K. E. & Smith, K. A. Nitrous oxide emission factors for agricultural soils in Great Britain: the impact of soil water‐filled pore space and other controlling variables. Global Change Biology 9, 204–218 (2003).
Brumme, R. & Beese, F. Effects of liming and nitrogen fertilization on emissions of CO2 and N2O from a temperate forest. Journal of Geophysical Research: Atmospheres 97, 12851–12858 (1992).
Gong, Z. Chinese soil taxonomy. (Science press, 2001).
Peng, Y. et al. Linkages of plant stoichiometry to ecosystem production and carbon fluxes with increasing nitrogen inputs in an alpine steppe. Global change biology 23, 5249–5259 (2017).
Tian, Q. et al. A novel soil manganese mechanism drives plant species loss with increased nitrogen deposition in a temperate steppe. Ecology 97, 65–74 (2016).
Zhang, K. et al. Corrigendum: Effects of Short-Term Warming and Altered Precipitation on Soil Microbial Communities in Alpine Grassland of the Tibetan Plateau. Frontiers in Microbiology 8, 667 (2017).
Acknowledgements
This study is funded by the National Science and Technology Support Project (Grant No. 2014BAC05B01), National Natural Science Foundation of China (Grant No. 40801076), National Program on Basic Work Project of China (Grant No. 2015FY11030001) and Strategic Priority Research Program of CAS (Grant No. XDA0505030304). We thank Zebing Zhong, Hechun Liu, Feng Xiong and Yi Ning for facilitating our field surveys of the Tibetan Plateau (2014–2015) and providing laboratory assistance.
Author information
Authors and Affiliations
Contributions
G.Z. designed the study and supervised the project. C.L., X.N., L.Y., F.L., Y.X. and K.F. collected the data. C.L., Y.P. and Y.Y. wrote the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Li, C., Peng, Y., Nie, X. et al. Differential responses of heterotrophic and autotrophic respiration to nitrogen addition and precipitation changes in a Tibetan alpine steppe. Sci Rep 8, 16546 (2018). https://doi.org/10.1038/s41598-018-34969-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-34969-5
- Springer Nature Limited
Keywords
This article is cited by
-
Response of soil respiration to water and nitrogen addition and its influencing factors: a four-year field experiment in a temperate steppe
Plant and Soil (2022)
-
Seasonal variation in soil CO2 emission and leaf gas exchange of well‐managed commercial Citrus sinensis (L.) orchards
Plant and Soil (2021)
-
Spatial differentiation of the NPP and NDVI and its influencing factors vary with grassland type on the Qinghai-Tibet Plateau
Environmental Monitoring and Assessment (2021)