
Abstract
Dams commonly restrict fish movements in large rivers but can also help curtail the spread of invasive species, such as invasive bigheaded carps (Hypophthalmichthys spp). To determine how dams in the upper Mississippi River (UMR) affect large-scale invasive and native fish migrations, we tracked American paddlefish (Polyodon spathula) and bigheaded carp across > 600 river km (rkm) and 16 navigation locks and dams (LD) of the UMR during 2 years with contrasting water levels. In 2022, a low-water year, both native paddlefish and invasive bigheaded carp had low passage rates (4% and 0.6% respectively) through LD15, a movement bottleneck being studied for invasive carp control. In contrast, flooding in 2023 led to open-river conditions across multiple dams simultaneously, allowing 53% of paddlefish and 46% of bigheaded carp detected in Pool 16 to move upstream through LD15. Bigheaded carp passed upstream through LD15 rapidly (μ = 32 rkm per day) a maximum of 381 rkm, whereas paddlefish moved an average of 9 upstream rkm per day (maximum of 337 rkm). Our results can inform managers examining trade-offs between actions that enhance native fish passage or deter movements of invasive species. This understanding is critical because current climate change models project increases in flooding events like that observed during 2023.
Similar content being viewed by others
Introduction
The migratory behaviors of native and invasive fishes are affected by dams in rivers throughout the world1. Dams can limit the movements and dispersal of fish populations in ways that restrict access to important habitats used for spawning and feeding2. Furthermore, dams can restrict the connections between different segments of fish populations, thereby creating challenges for natural resource managers working to conserve and restore threatened or endangered species3,4. Alternatively, dams can also function as effective barriers to the dispersal of harmful invasive species by limiting their spread5,6,7,8. The tension between the need to permit or enhance the movements of native fishes and efforts to curtail the movements of invasive species represents a difficult challenge for riverine fisheries managers9,10,11,12.
The upper Mississippi River (UMR) basin has a high degree of channel fragmentation13 due to a series of 29 navigation locks and dams that can affect fish migration. Fish passage through a dam is influenced by the design features of individual structures. Each dam features a series of adjustable spillway gates (i.e., Tainter and/or roller gates) that can be raised and lowered to regulate upstream water levels, as well as a navigation lock to facilitate movement of vessels between pools. The gated portions of the UMR navigation dams are operated to maintain a minimum 9-foot (2.75-m) navigation channel14 in the river and can be raised fully out of the river during high-water events (i.e., open-river condition15,16) to permit the dispersal of floodwaters, thereby creating a period when fish can readily pass through these dams. As the gates are lowered into the river, upstream passage becomes more difficult. When the gates are lowered, hydraulic head (i.e., the difference in water elevation above the dam relative to below the dam) and current velocity through the partially closed gates are commonly so high that upstream passage is restricted to the navigation lock of each lock and dam4,15,17.
Previous research has documented the important roles specific dams play in regulating fish dispersal in the UMR because each lock and dam (LD) differs in the range of hydraulic head and the percentage of time in open-river condition. High hydraulic head at LD 5–LD 8 effectively halts upstream dispersal by migratory American paddlefish (Polyodon spathula) whenever dam gates are partially closed4. Upstream dispersal of invasive species is also affected by hydrologic conditions. Bigheaded carp (i.e., bighead carp Hypophthalmichthys nobilis and silver carp H. molitrix) at Starved Rock Lock and Dam on the Illinois River generally passed upstream through the gated portion of the dam during open-river conditions and made upstream passage through the lock chamber during periods with high hydraulic head7. A study on passage rates of 11 species through UMR LD 20–LD 26 and the Chain of Rocks weir showed that, in general, fish tend to move upstream through dam gates in greater proportions to lock passages whenever dam gates are raised from the river during open-river conditions18. Numerous studies have documented that a diverse range of fish species appeared to be able to make downstream passage through the gated portion of dams under a wide range of hydraulic conditions18,19,20,21,22.
Restricted upstream passage at certain dams has limited the spread of invasive bigheaded carp. LD 19, which has a hydroelectric facility and the highest vertical lift of the active navigation locks on the UMR, has a hydraulic head that is too great to permit fish passage through the gated portion of the dam23,24,25. Farther upstream, LD 15 and LD 14 are infrequently in open-river conditions. Together, the restrictions on upstream passage at LD 19 and LD 15 have played a large role in reducing the population of invasive carp upstream from LD 15. Upstream from LD 19, adult abundance is considered relatively low, with captures comprising less than 25% of samples, while adults are common downstream from LD 19 (25–90% of samples)26. Reproduction occurs intermittently between LD 19 and LD 15, and annually downstream from LD 19. A variety of socioecological problems accompany increasing bigheaded carp populations, from safety hazards to boaters to food web disruptions27,28. To prevent further upstream spread, the portion of the UMR between LD 19 and LD 15 is considered a focal point for invasive carp control.
Managers need to balance the tradeoffs of reducing invasive carp passage and facilitating passage of native species; to do so effectively, they need information on how fish passage is affected by dam operations and other influential factors, such as temperature. For instance, paddlefish remain sedentary during winter months but begin to actively migrate upstream into areas with deep water and rocky substrates once water temperatures reach 10 °C29. Bigheaded carps were documented moving into the tailwaters downstream from Starved Rock Lock and Dam during spring, when water temperatures began to rise above 5 °C7. Any differences in movement cues between native species and invasive bigheaded carp could be useful for managers to consider for selective passage. Additionally, climate change may result in changes to movement cues, such as the timing of seasonal flood pulses. In the UMR, floods have increased in magnitude and duration in recent decades, and some models indicate this trend may continue in the coming decades30,31.
The purpose of this study was to examine movements of paddlefish and bigheaded carp with respect to dams and the abiotic factors affecting both dam operations and fish physiology. We used consecutive years with contrasting conditions to explore how fish responses changed as conditions varied between 2022, a low-water year, and 2023, and a high-water year. This work builds upon earlier efforts to describe the timing of dam passage22 and the hydraulic effects of dam operation on fish passage through dam gates19. This study describes invasive and native fish large-scale migrations in a large navigable river with a study area that encompasses over 600 km of riverine habitats and 16 navigation locks and dams. A large-scale comparison of native and invasive fish passage is an important step to facilitate a stronger understanding of how climatic variables such as precipitation and warming temperatures affect the abilities of native and invasive species to traverse the existing infrastructure of the UMR. Compared to existing studies on fish passage that typically studied movements around a single structure, this regional-scale understanding is critical for managers tasked with conserving native fish population connectivity while taking measures to slow the advance of non-native species. Our objectives are to (1) compare the proportion of available fish with surgically implanted internal acoustic transmitters migrating upstream through a key fish movement bottleneck dam (i.e., LD 15), (2) evaluate the relation of upstream migrations to environmental factors (i.e., hydrologic conditions, water temperature), and (3) evaluate the maximum upstream migratory distance and rate of movement exhibited by invasive bigheaded carp and native paddlefish.
Methods
Study area
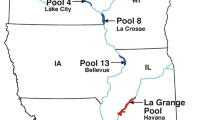
The study area encompassed > 600 river kilometers (rkm) of the Mississippi River, ranging from Pool 4 near Lake City, Minnesota, downstream to Pool 19 near Keokuk, Iowa (Fig. 1). The pools of the Mississippi River are named sequentially moving downstream and are created by lock and dam structures at the downstream end of each pool. Widespread drought occurred in the upper Mississippi River watershed throughout much of 202232, aside from brief heavy rain events during May 2022. In contrast, widespread and extended flooding occurred during April and May 2023 due to snowmelt runoff33.

Map of the study area, including the Lock and Dam (LD) locations along the Mississippi River and the length of each pool between each lock and dam. The pools of the Mississippi River are named sequentially moving downstream and are created by lock and dam structures at the downstream end of each pool. Created with ArcGIS Pro 3.0.0; https://hub.arcgis.com/content/290e4ab8a07f4d2c8392848d011add32/about; https://esri.maps.arcgis.com/home/item.html?id=8c2d6d7df8fa4142b0a1211c8dd66903.
Fish collection and implantation of acoustic transmitters
We captured adult silver carp, bighead carp, and paddlefish using gill nets. Most fish were captured in Pools 14–19 of the UMR during 2017–2022. Additional silver carp (N = 16) were surgically implanted with transmitters in Pool 5A and Pool 8 during 2020–2023. Surgical implantation of acoustic transmitters generally followed Lubejko et al.7. We implanted fish with 69-kHz acoustic transmitters with random ping rates of 30–90 s or 80–160 s (Innovasea, model V16-4H and V16-6H). Implanted transmitters did not exceed 2% of the fish’s weight34. We tested transmitters to ensure proper functionality and soaked them for ≥ 2 min in an alcohol disinfectant solution prior to surgical implantation. Fish were sedated with high purity carbon dioxide gas from a compressed gas cylinder that was administered into river water in a 140-L polyethylene sedation bath via direct gas injection until the fish reached Stage 4 anesthesia35,36,37,38. All fish were weighed and measured after sedation. The incision was closed with three non-absorbable sutures (3–0 [669H], Ethicon Inc., Somerville, New Jersey). Fish recovered in a tank at the surgery site until equilibrium was restored and were released within 0.5 rkm of the original capture location. Captured fish were handled in accordance with the U.S. Geological Survey Upper Midwest Environmental Sciences Center Institutional Animal Care and Use Committee approved procedures, under IACUC protocol number ESB-19-LD19CARP-01. This study complied with ARRIVE guidelines and best animal welfare practices39,40.
Acoustic receiver arrays
Fish movements within the UMR were monitored with 69-kHz acoustic telemetry receiver arrays (VR2Tx, VR2W, VR2C; Innovasea, Nova Scotia, Canada). The focal array was a large-scale longitudinal receiver array (> 600 river km) that is maintained by the U.S. Fish and Wildlife Service, Minnesota Department of Natural Resources, Illinois Natural History Survey, and U.S. Geological Survey to detect fish movement across dams in Pools 4–19 of the Mississippi River. Receivers in the longitudinal array were deployed using a combination of mounts and deployment strategies. Receivers were affixed to UMR navigation channel buoys with rebar mounts orienting the hydrophone towards the river bottom. Shore-based receiver mounts consisted of a metal frame deployed on the river bottom and tethered to a tree or other shore structure, with the hydrophone pointing away from shore. Receiver placement was chosen based on both channel morphology and availability and accessibility of appropriate mounting structures to maximize the likelihood of fish detection. Passive receivers were downloaded every 4–8 weeks during March–November each year. Shore-based receiver mounts were left in place overwinter to record movement data while ice conditions prevented the use of channel buoy mounted receivers.
In addition to the longitudinal acoustic receiver array, we used data from two small-scale 69-kHz telemetry arrays deployed at LD 14 and LD 15 to determine route of upstream passage at these bottleneck dams (i.e., through the lock chamber or through the gated portion of the dam). Receivers were installed year-round in ladder wells to prevent damage and collisions with barges and other vessels. The placement of receivers in ladder recesses did reduce the field of view, but the receivers were still able to detect test transmitters throughout the focal zones of interest (i.e., downstream lock approach, lock chamber, and upstream lock approach; Supplementary Fig. S1 online). Upstream passage route though the lock chamber was identified with a detection sequence in the downstream lock approach, lock chamber, and upstream lock approach. If a fish moved upstream through a dam without following this detection sequence, the passage was presumed to have occurred through the gated portion of the dam. We conducted range tests to validate receiver detection efficiency using transmitters with a 7-s fixed interval plus a 3-s transmission period. The detection rate was calculated as the number of observed detections divided by the number of expected detections, multiplied by 100, where number of expected detections is calculated from the known delay between transmissions and the duration of the test41. Range testing was conducted in May and November of 2022 and July and November of 2023 (Supplementary Fig. S1 online). The average detection rate of available transmissions (± standard error [SE]) was 85 ± 4% for receivers in the downstream lock approach, 89 ± 3% for receivers in the lock chamber, 73 ± 11% for receivers in the auxiliary lock chamber, and 87 ± 2% for receivers in the upstream lock approach at LD 14 and 91 ± 2% for receivers in the downstream lock approach, 91 ± 3% for receivers in the lock chamber, 86 ± 3% for receivers in the auxiliary lock chamber, and 84 ± 4% for receivers in the upstream lock approach at LD 15.
Data processing and statistical analyses
To identify the proportion of fishes passing upstream, we converted raw detection data into discrete residency events using the ‘residence event’ function in the VTrack package in R (v1.2142). All analyses were performed in R (v.4.2.143). A residency event was defined by at least two detections at a receiver within 12 h. Residency events were assigned to the pool in which the corresponding receiver was deployed. Identification of dam passage events followed Vallazza et al.21 and identified the last location and date in the first pool and the first location and date in the second pool. For locations that had receivers with potential overlap in detection range on opposite sides of a dam (i.e., LD 14, LD 15), passage through those dams were confirmed with subsequent detections on receivers farther from the dam. Occasionally, fish were not detected within a pool but were subsequently detected farther upstream, thus confirming successful movement through the previous pool. This occurred more frequently in pools with fewer receivers and more braided channel morphology. In these instances, the fish that passed upstream without being detected in the intermediate pool were assigned a date within the intermediate pool that was the midpoint between their last confirmed detection in the previous pool and their first confirmed detection in the upstream pool.
We summarized hydraulic head at a daily level using data from the U.S. Army Corps of Engineers, who maintain records on the locks and dams used in this study. For the period of January 1, 2022, to December 11, 2023, we calculated hydraulic head as the difference between the pool elevation upstream from a dam and the tailwater elevation downstream from a dam. The temporal resolution of the data varied among dams, with daily values available at LD 4 through LD 10 and 30-min data available at LD 11 through LD 18. We used the daily measurements of hydraulic head at LD 4 through LD 10, and the daily minimum hydraulic head for LD 11 through LD 18 because we thought the lowest level available to fish in a given day was the most relevant for upstream passage at dams. We also analyzed water temperature at LD 15, where water temperatures were recorded at 30-min intervals. We used the daily mean water temperature to characterize water temperatures at LD 15.
To assess the influence of hydraulic head and water temperature on fish passage at LD 15, we used generalized linear mixed models (GLMM; binomial distribution, logit link) in R package lme4 (v1.1-35.144). For each day an individual fish was detected in Pool 16, it was classified as passing upstream into Pool 15 (1) or not passing upstream (0). In some cases, the timing of passage at LD 15 was uncertain, such as when the last detection downstream from LD 15 and the first detection upstream of LD 15 were separated by more than a day. We assigned the date of upstream passage in these instances as the midpoint between the last detection downstream from LD 15 and the first detection upstream from LD 15. We included transmitter ID as a random effect to account for individuals that made more than one upstream passage. We used the R package DHARMa (v0.4.645) to evaluate model assumptions. Previous research in this system indicated that a small proportion of fish available to pass upstream from LD 15 do so during years without floods21,22. The low rates of upstream passage at LD 15 in previous years21,22 strongly indicates that abiotic factors limited fish movements, rather than characteristics of fish themselves. We therefore modeled two of the most important environmental controls for fish passage—i.e., hydraulic head and water temperature—rather than including variables for individuals, such as total length, weight, or sex. We used Kendall’s tau to evaluate correlation between hydraulic head and water temperature. We assessed statistical significance using α = 0.05. All data are publicly available through ScienceBase (https://doi.org/10.5066/P14GQIVU)46.
To describe the magnitude of fish migrations, we examined physical distances traveled through two scenarios: overall range of distance traveled and rapid migration. A range of distance traveled was calculated for each fish using the homerange function in the R package riverdist (v0.16.147), which charts the migration path using observed detections and measures the total extent of its boundaries. The overall range of distance traveled by individual fish was described between a low-water year in 2022 and a high-water year in 2023. This range calculation was applied only to fish that were present in UMR Pools 16–19 at any point during 2022 or 2023 to retain a focus on the effects of the bottleneck of LD 15. For the rapid migration that occurred during 2023, we identified fish that were present in UMR Pools 16–19 and then subsequently completed upstream passage through LD 15. The distance and duration each fish traveled once upstream from the bottleneck of LD 15 was calculated by using the difference in the river kilometer assigned to each receiver and the times of detection when the fish was recorded. To precisely capture the rate of these rapid movements, we used a 7-day timeout, where the time between detections among receivers must occur within 7 days or less. Detection intervals that were separated by more than 7 days were not able to precisely capture the snapshot in time during the rapid movement. These extended interval data were not used in the calculation of the rate and extent of the rapid movement, even if a fish showed farther progression upstream at a later date. The time of the first detection upstream from LD 15 was used to define the beginning of the upstream movement and concluded at the time of arrival on the farthest upstream receiver. These sequential detection records convey the sustained rate of movement these fishes displayed in 2023.
To visually depict the days in which each lock and dam operated in open-river conditions, we used a threshold of 0.46 m or less as the hydraulic head value to denote putative open-river conditions. At LD 11 through LD 18, information also was available on the days in which the gates were pulled out of the water, and we therefore also classified any day in which the gates were pulled out of the water as open-river conditions at these dams. The design of each dam varies, and no single value characterizes open-river conditions based on hydraulic head across all the dams in the study15; however, the hydraulic head threshold we chose is one used as a guideline based on input from U.S. Army Corps of Engineers48. Although we chose to use a value of 0.46 m as the hydraulic head threshold between open-river and controlled operation, we acknowledge this is a simplification for the purposes of plotting figures. All statistical analyses used daily values for hydraulic head and water temperature.
Results
Proportion of fishes passing upstream from LD 15 in 2022 and 2023
Between January 1, 2022, and December 12, 2023, 480 bigheaded carp (400 silver carp, 65 bighead carp, and 15 hybrids) and 124 paddlefish were detected between Pool 4 and Pool 19 of the UMR across 93 passive acoustic telemetry receivers. In 2022, 94% of implanted bigheaded carp were detected downstream from LD 15, in Pools 16–19. During 2022, 174 bigheaded carp were detected in Pool 16, and only one passed upstream at LD 15 (Supplementary Table S1). The route of upstream passage was through the LD 15 main lock chamber; this same individual subsequently completed upstream passage through the main lock chamber of LD 14. In contrast, 46% of implanted bigheaded carp (75 of 163) in Pool 16 passed upstream through LD 15 in 2023 (Fig. 2; Supplementary Fig. S2 online), representing 21% of all implanted bigheaded carp detected in Pools 16–19 (75 of 354). We determined that all bigheaded carp upstream passages through LD 15 during 2023 occurred through the gated portion of the dam because no bigheaded carp were detected making upstream passage through the main or auxiliary lock. Paddlefish displayed a similar pattern, with 4% (1 of 25) of individuals detected in Pool 16 during 2022 passing upstream through LD 15 (via the lock chamber), whereas 53% (17 of 32) passed upstream through LD 15 in 2023 (Fig. 3). Most paddlefish (15 of 17) completed upstream passage through the gated portion of the dam, with two individuals completing upstream passage through the auxiliary lock chamber.

Upstream passages by implanted bigheaded carp from Lock and Dam 4 to Lock and Dam 18 in 2022 (a) and 2023 (b). Gray bars indicate dates of putative controlled river conditions for each lock and dam, based on a threshold hydraulic head value of 0.46 m, with white spaces between indicating dates of putative open-river conditions. The bottom of each blue arrow indicates the last date a fish was detected in the downstream pool, and the point of each arrow indicates the first date it was detected in the upstream pool. Upstream passages with a detection interval between upper Mississippi River pools of 21 days or less are depicted to align the river conditions most precisely with the timing of upstream passage. Expanded information on upstream and downstream passages can be accessed in Supplementary Fig. S2.

Upstream passages by implanted paddlefish between Lock and Dam 4 and Lock and Dam 18 in 2022 (a) and 2023 (b). Gray bars indicate dates of putative controlled river conditions for each lock and dam, based on a threshold hydraulic head value of 0.46 m, with white spaces between indicating dates of putative open-river conditions. The bottom of each blue arrow indicates the last date a fish was detected in the downstream pool, and the point of each arrow indicates the first date it was detected in the upstream pool. Upstream passages with a detection interval between upper Mississippi River pools of 21 days or less are depicted to align the river conditions most precisely with the timing of upstream passage. Expanded information on upstream and downstream passages can be accessed in Supplementary Fig. S2.
Relation of environmental factors to passage at LD 15
Upstream passage at LD 15 by bigheaded carp significantly increased as hydraulic head decreased (n = 216; z = − 8.901; p < 0.001) but was not strongly related to water temperature (n = 216; z = 1.678; p = 0.09; Fig. 4; Supplementary Fig. S4 online). Similarly, upstream passage at LD 15 by paddlefish significantly increased with a decrease in hydraulic head (n = 34; z = − 4.196; p < 0.001) but was not related to water temperature (n = 34; z = − 0.346; p = 0.73; Fig. 5). Upstream passage at other dams in the study (including LD 5) also tended to occur during periods of low hydraulic head and open-river conditions (Supplementary Fig. S3 online). There was a weak, positive correlation between hydraulic head and water temperature (Kendall’s tau = 0.11; 95% confidence interval [CI] 0.07–0.15; z = 4.36, p < 0.001).

Hydraulic head at Lock and Dam 15 in 2022 (a) and 2023 (b), with the number of bigheaded carp making upstream passage each day in 2022 (c) and 2023 (d) and the number of bigheaded carp detected in Pool 16 each day in 2022 (e) and 2023 (f). The horizontal dashed line in a and b indicates a hydraulic head value of 0.46 m, a threshold that generally represents open-river conditions across dams. Asterisk indicates a large increase in the number of tagged bigheaded carp detected in Pool 16 from a tagging event that occurred in Pool 16 during April 2022.

Hydraulic head at Lock and Dam 15 in 2022 (a) and 2023 (b), with the number of paddlefish making upstream passage each day in 2022 (c) and 2023 (d) and the number of paddlefish detected in Pool 16 each day in 2022 (e) and 2023 (f). The horizontal dashed line in a and b indicates a hydraulic head value of 0.46 m, a threshold that generally represents open-river conditions across dams.
Rate of movement and maximum upstream migration
Comparisons between study years showed the mean range of distance traveled for bigheaded carp and paddlefish was larger in 2023. Mean and maximum range of distance traveled for bigheaded carp were approximately three times greater in 2023 relative to 2022 (Table 1). We quantified the migratory distance and rate of upstream movement for bigheaded carp and paddlefish that successfully completed upstream passage through LD 15 in 2023 and continued moving upstream. There was no rapid upstream migration through LD15 during 2022. The mean distance upstream from LD 15 that bigheaded carp traveled in 2023 was 270 rkm (range: 48–381 rkm; n = 53). Paddlefish tracked migrating upstream through LD 15 in 2023 had a shorter mean distance traveled upstream from LD 15 (120 rkm, range 16–337, n = 6). The mean daily rate covered by migrating fishes in 2023 was 32 rkm/day (range 7–50 rkm/day) for bigheaded carp and 9 rkm/day (range 2–16 rkm/day) for paddlefish. The mean distance of 32 km/day converts to a continuous 0.37 m/s groundspeed for bigheaded carp, and the maximum distance of 50 km/day converts to continuous groundspeed of 0.58 m/s. These measurements represent the prolonged swimming groundspeed and stamina of bigheaded carp over the course of numerous days in a large navigable river during flood conditions.
Discussion
Our results highlight the extent to which dam operations and hydrologic conditions affect fish movements across a vast spatial scale in the UMR. Dam operations during controlled hydrologic conditions can limit native and invasive fish upstream passage. Conversely, during uncontrolled hydrologic conditions, when the dams allowed for open-river conditions, fish moved rapidly upstream. Characterizing the magnitude and relative duration of periods when upstream migrations are inhibited by dams is important for both the conservation of native fishes and control of harmful invasive species. Likewise, understanding fish behavior during brief windows of unregulated hydrological conditions, when dam gates are raised from the water, is also important for natural resource managers to consider when developing fisheries management plans.
Paddlefish and bigheaded carp responded similarly to the different hydrologic conditions occurring during 2022 and 2023. Dams that do not enter open-river conditions under most river discharge levels (e.g., LD 15, LD 14, LD 5) appear to block upstream movements of paddlefish and bigheaded carp. Both taxa appeared to opportunistically exploit relatively short periods of open-river conditions to make rapid upstream migrations. The period of intense flooding that occurred during spring 2023 offered a unique opportunity to observe these species’ behavior during a period when nearly all UMR dams were functioning in open-river conditions. Paddlefish and, especially, bigheaded carp initiated rapid upstream movements en masse when they were able to navigate through the open dam gates.
Only hydraulic head, not water temperature, was significantly associated with upstream passage through LD 15. However, water temperature may still play an important role in initiating these upstream migrations. Water temperatures were nearly identical during the period of fish migrations in 2022 and 2023 (between 10 and 22 °C). Generally, fish appeared to be more motivated to move upstream as the rate of warming increased as evidenced by the timing of the congregation of individuals in the tailwaters of LD 15 and LD 5 during periods of similar temperatures in May 2022 and 202349. However, the availability of movement corridors through or around barriers to movement (i.e., locks and dams) appears to be the most important factor in predicting fish passage through bottleneck dams as bigheaded carp made upstream passage through LD 15 in much greater numbers during 2023 during a period of open-river conditions. Likewise, a single implanted silver carp was also observed passing LD 5 during open-river conditions indicating that a similar pattern of fish migration may be occurring at this upstream barrier.
The large-scale migratory responses of paddlefish and bigheaded carp to unregulated river conditions observed in this study indicates that these species respond rapidly to changing hydrologic conditions. While outside the focus of this study, one can infer that the upstream movement during 2023 may be indicative of how native UMR fish species might have behaved prior to the installation of this network of locks and dams. These structures began to be installed during the mid-nineteenth century and slowly expanded until the completion of the current system of dams in 1940. Prior to these developments, paddlefish and other native species could migrate freely throughout the UMR, aside from sections of major rapids and waterfalls50,51. Fish passages through natural barriers were likely enhanced during periods of flooding. Paddlefish and other native fishes likely evolved behaviors to exploit these relatively narrow windows of opportunity to cross these natural barriers52. Likewise, bigheaded carp undoubtedly experienced similar intermittent impediments to upstream migrations in their native range53 and evolved similar behavioral cues to confront these environmental challenges (e.g., jumping behaviors or high burst swim speeds54). Directional cues can arise through group interactions based on the theory of collective navigation55. Berdahl et al.56 defines collective navigation as “the outcome of navigating within a social context.” Okasaki et al.57 provides evidence that chinook salmon (Oncorhynchus tshawytscha) benefit from collective navigation when attempting to traverse fish ladders in the Columbia River. It is possible that similar behavioral adaptations have emerged in paddlefish and bigheaded carp that enhance their abilities to navigate through these dams in groups. Future studies could investigate the potential role of collective navigation in facilitating mass movements of fishes through gated dams like those in the UMR.
A proportion of the available populations of paddlefish and bigheaded carp were able to rapidly transition upstream through a series of gated dams during hydrologic conditions that necessitated dam gates to be fully open. The relative speed of these upstream migrations during intense flooding periods provides strong indication that both species possess behavior patterns to facilitate rapid upstream movements when human-made barriers are inundated. Recent efforts to evaluate finer-scale movements of native fishes and bigheaded carp through navigation locks and dam gates indicate that these species can use either pathway to make upstream passage7,18,19,20,22. The dam gates at LD 5, 14, and 15 are infrequently in open-river condition and restrict upstream fish movements to the navigation lock during much of the year15,58. Downstream movements of fish through the gated portion of dams are less affected by partially closed gates15,21,25,59,60. Although the temporal extent of our data collection may be relatively short, our study indicates that instances of upstream passage through the navigation lock at LD 15, a known migratory bottleneck in the UMR, are relatively isolated compared to the mass upstream movements observed through the dam gates at that same location during open-river conditions (current study;22). Of our implanted population detected in Pool 16 during 2023, 46% (75 of 163) of bigheaded carp and 53% (17 of 32) of paddlefish made upstream passage through LD 15 compared to < 1% and 4% of available bigheaded carp and paddlefish, respectively, in 2022. The differing frequencies of fish passage for both species between 2022 and 2023 provides a contrast in how those species respond to different hydrologic conditions and relatively brief windows of favorable opportunities to pass human-made barriers.
The implanted bigheaded carp we observed appeared to halt their upstream migration in Pool 5A. This may reflect the cessation of open-river conditions at LD 5 rather than the potential stamina limit of the maximum upstream distance that a fish could swim if in open water. Hydraulic head at LD 5 exceeded 0.46 m and the dam was operating in controlled conditions by the date bigheaded carp passing upstream from Pool 16 traveled the 400 km to reach LD 5 (Fig. 2). Given the observed upstream passage through LD 5 and LD 4 during open-river conditions by a bigheaded carp that was originally captured and implanted in Pool 8, it is reasonable to suspect the bigheaded carp migration could have continued beyond Pool 5A if open-river conditions were present at the date of their arrival. Additionally, most of the bigheaded carp that passed upstream from LD 15 did not return downstream of LD 15 by the end of the study period (Supplementary Fig. S2 online). These bigheaded carp, now in closer proximity to LD 5, may be able to rapidly move upstream if LD 5 again enters open-river conditions.
Our study supports the conclusions of previous studies that periods of elevated discharge may serve as important opportunities for population connection for native species, such as paddlefish and lake sturgeon3,4,22,61. Efforts to develop and deploy deterrent technologies designed to limit the movements of bigheaded carp and disconnect their populations should balance the needs of native species that also benefit from these intermittent pathways created by dam operations. These concerns represent a difficult trade-off for river managers tasked with protecting important native species and controlling the spread of harmful non-native species6,9,62. Additionally, studies that estimate bigheaded carp movements through navigation locks and dams that can experience open-river conditions20,22 would benefit from considering the capacity of these injurious species to rapidly move en masse through dam gates during flooding conditions, thereby reducing or negating the benefit of reducing upstream passages through lock chambers. Timing of operation of deterrents, development of selective deterrents for the gated portion of dams, and the development and implementation of selective fish passage technologies warrant consideration63.
The movements we observed in 2023 indicate floods represent potentially profound vulnerabilities to the control of invasive bigheaded carp moving upstream. Controlled studies in swim flumes64,65 and conceptual hydrological models66 indicate specific dam gate operation regimes may be used to inhibit upstream passage of bigheaded carp. However, it is possible that the relatively rare instances of open-river conditions may be the most influential events that govern the populations of bigheaded carp at the leading edge of their distribution. The specific timing of intermittent river flooding is unpredictable. This makes it difficult for natural resource managers to accurately predict when fishes may attempt upstream passage, particularly if climatic conditions become more variable in the future30,31,67.
The insights from the present study were made possible because of extensive efforts to implant acoustic transmitters into bigheaded carp in areas of relatively high density, as well as the effort to deploy a network of acoustic receivers across more than 600 river km. Without the large quantity of native and invasive fishes implanted with acoustic transmitters and extensive receiver coverage, we would not have been able to identify the precise timing and associated hydrologic conditions of fish passage. The interpretation and application of our findings may be limited by the fact that only two contrasting years’ data were used for these analyses. Therefore, river managers and researchers may benefit from maintaining and possibly expanding telemetry networks in the UMR to provide further documentation of fish migrations under differing environmental conditions. Future studies could also examine long-term telemetry data to further investigate the roles of water temperature and other environmental factors in influencing the migratory behaviors of bigheaded carp and paddlefish in the UMR. Greater understanding of these broad-scale migration patterns can enhance the future management of these species that routinely traverse jurisdictional boundaries. Furthermore, increasing the understanding of how fishes move through lock and dam structures is critical for developing deterrent strategies that effectively limit the spread of aquatic invasive species while protecting migration corridors for native fishes.
Data availability
All data are publicly available through ScienceBase at https://doi.org/10.5066/P14GQIVU46.
References
McIntyre, P. et al. Conservation of migratory fishes in freshwater ecosystems. In Conservation of Freshwater Fishes (eds Closs, G. et al.) 324–360 (Cambridge University Press, 2016).
Kruk, A. & Penczak, T. Impoundment impact on populations of facultative riverine fish. Int. J. Limnol. 39, 197–210 (2003).
Knights, B. C., Vallazza, J. M., Zigler, S. J. & Dewey, M. R. Habitat and movement of lake sturgeon in the upper Mississippi River system, USA. Trans. Am. Fish. Soc. 131, 507–522 (2002).
Zigler, S. J., Dewey, M. R., Knights, B. C., Runstrom, A. L. & Steingraeber, M. T. Hydrologic and hydraulic factors affecting passage of paddlefish through dams in the upper Mississippi River. Trans. Am. Fish. Soc. 133, 160–172 (2004).
Rahel, F. J. Intentional fragmentation as a management strategy in aquatic systems. BioScience 63, 362–372 (2013).
Rahel, F. J. & McLaughlin, R. L. Selective fragmentation and the management of fish movement across anthropogenic barriers. Ecol. Appl. 28, 2066–2081 (2018).
Lubejko, M. V. et al. Evaluating upstream passage and timing of approach by adult bigheaded carps at a gated dam on the Illinois River. River Res. Appl. 33, 1268–1278 (2017).
Jones, P. E. et al. The use of barriers to limit the spread of aquatic invasive animal species: A global review. Front. Ecol. Evol. 9, 611631 (2021).
Cooper, A. R. et al. Prioritizing native migratory fish passage restoration while limiting the spread of invasive species: A case study in the Upper Mississippi River. Sci. Total. Environ. 791, 148317 (2021).
Fausch, K. D., Rieman, B. E., Dunham, J. B., Young, M. K. & Peterson, D. P. Invasion versus isolation: Trade-offs in managing native salmonids with barriers to upstream movement. Conserv. Biol. 23, 859–870 (2009).
Zielinski, D. P., McLaughlin, R. L., Pratt, T. C., Goodwin, R. A. & Muir, A. M. Single-stream recycling inspires selective fish passage solutions for the connectivity conundrum in aquatic ecosystems. BioScience 70, 871–886 (2020).
Franklin, P. A. et al. Aquatic connectivity: Challenges and solutions in a changing climate. J. Fish Biol. https://doi.org/10.1111/jfb.15727 (2024).
Nilsson, C., Reidy, C. A., Dynesius, M. & Revenga, C. Fragmentation and flow regulation of the world’s large river systems. Science 308, 405–408 (2005).
Alexander, J. S., Wilson, R. C. & Green, W. R. A brief history and summary of the effects of river engineering and dams on the Mississippi River system and delta: U.S. Geological Survey Circular 1375 (2012). https://doi.org/10.3133/cir1375.
Wilcox, D. et al. Improving fish passage through navigation dams on the Upper Mississippi River system (U.S. Army Corps of Engineers, 2004).
Theiling, C. H. & Nestler, J. M. River stage response to alteration of Upper Mississippi River channels, floodplains, and watersheds. Hydrobiologia 640, 17–47 (2010).
Osvalt, R. & Grace, J. Velocity field in roller/tainter gate bays of Upper Mississippi River structures (U.S. Army Corps of Engineers, 1984).
Tripp, S., Brooks, R., Herzog, D. & Garvey, J. Patterns of fish passage in the upper Mississippi River. River Res. Appl. 30, 1056–1064 (2014).
Whitty, J. M., Riesgraf, A. T., Zielinski, D. P. & Sorensen, P. W. Movements of a model fish, the common carp, through a generic Mississippi River lock and dam demonstrate how fish swimming performance, behavior, and discharge-driven flow-fields determine fish passage rates in ways that can be predicted and modified using fish passage models. River Res. Appl. 38, 670–683 (2022).
Finger, J. S., Riesgraf, A. T., Zielinski, D. P. & Sorensen, P. W. Monitoring upstream fish passage through a Mississippi River lock and dam reveals species differences in lock chamber usage and supports a fish passage model which describes velocity-dependent passage through spillway gates. River Res. Appl. 36, 36–46 (2020).
Vallazza, J. M. et al. Timing and hydrological conditions associated with bigheaded carp movement past navigation dams on the upper Mississippi River. Biol. Invasions 23, 3409–3425 (2021).
Turney, D. D. et al. Hydrological and lock operation conditions associated with paddlefish and bigheaded carp dam passage on a large and small scale in the Upper Mississippi River (Pools 14–18). PeerJ 10, e13822 (2022).
Coker, R. E. Keokuk Dam and the fisheries of the Upper Mississippi River. Bull. U.S. Bur. Fish. 45, 87–139 (1929).
U.S. Army Corps of Engineers. Upper Mississippi River basin master reservoir regulation manual, Appendix 19, Lock & Dam No. 19 (U.S. Army Corps of Engineers, 1981).
Fritts, A. K. et al. Lock operations influence upstream passages of invasive and native fishes at a Mississippi River high-head dam. Biol. Invasions 23, 771–794 (2021).
Jackson, N. & Runstrom, A. Upper Mississippi River Basin Asian Carp Control Strategy Framework. (Upper Mississippi River Asian Carp Partnership, Upper Mississippi River Conservation Committee Fisheries Technical Section, 2018).
DeBoer, J. A., Anderson, A. M. & Casper, A. F. Multi-trophic response to invasive silver carp (Hypophthalmichthys molitrix ) in a large floodplain river. Freshw. Biol. 63, 597–611 (2018).
Kolar, C. S. et al. Bigheaded Carps: A Biological Synopsis and Environmental Risk Assessment (American Fisheries Society, Special Publication 33, Bethesda, 2007).
Russell, T. R. Biology and life history of the paddlefish—A review. In The Paddlefish: Status, Management and Propagation (eds Dillard, J. G. et al.) 2–20 (American Fisheries Society, North Central Division, 1986).
Byun, K., Chiu, C.-M. & Hamlet, A. F. Effects of 21st century climate change on seasonal flow regimes and hydrologic extremes over the Midwest and Great Lakes region of the US. Sci. Total Environ. 650, 1261–1277 (2019).
Houser, J. N., (ed.) Ecological Status and Trends of the Upper Mississippi and Illinois Rivers (U.S. Geological Survey Open-File Report 2022–1039, 2022).
NOAA National Centers for Environmental Information. Monthly Drought Report for Annual 2022, accessed 24 July 2024; https://www.ncei.noaa.gov/access/monitoring/monthly-report/drought/202213 (2023).
NOAA National Centers for Environmental Information. Monthly Drought Report for Annual 2023, accessed 24 July 2024; https://www.ncei.noaa.gov/access/monitoring/monthly-report/drought/202313 (2024).
Winter, J. D. Advances in underwater biotelemetry. In Fisheries Techniques (eds Murphy, B. R. & Willis, D. W.) 555–590 (American Fisheries Society, 1996).
Ross, L. G. & Ross, B. Anaesthetic and Sedative Techniques for Aquatic Animals (Blackwell Publishing, 2008).
Trushenski, J. T., Bowker, J. D., Gause, B. R. & Mulligan, B. L. Chemical and electrical approaches to sedation of hybrid striped bass: Induction, recovery, and physiological responses to sedation. Trans. Am. Fish. Soc. 141, 455–467 (2012).
Oberg, E. W., Perez, K. O. & Fuiman, L. A. Carbon dioxide is an effective anesthetic for multiple marine fish species. Fish. Res. 165, 22–27 (2015).
Summerfelt, R. & Smith, L. Methods for fish biology. In Anaesthesia, Surgery and Related Techniques (eds Schreck, C. B. & Moyle, P. B.) 213–272 (American Fisheries Society, 1990).
Underwood, W. & Anthony, R. AVMA Guidelines for the Euthanasia of Animals 2020th edn. (American Veterinary Medical Association, 2020).
Bowker, J. & Trushenski, J. Guide to Using Drugs, Biologics, and Other Chemicals in Aquaculture (American Fisheries Society Fish Culture Section, 2019).
Kim, J. & Mandrak, N. E. Assessing the potential movement of invasive fishes through the Welland Canal. J. Great Lakes Res. 42, 1102–1108 (2016).
Campbell, H. A., Watts, M. E., Dwyer, R. G. & Franklin, C. E. V-Track: Software for analysing and visualising animal movement from acoustic telemetry detections. Mar. Freshw. Res. 63, 815 (2012).
R Core Team. R: A language and environment for statistical computing (R Foundation for Statistical Computing, 2022).
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48 (2015).
Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-level/Mixed) Regression Models (R package version 0.4.6., 2016).
Fritts, M. et al. Data Release Associated with Research that Describes How Major Flooding and Dam Operations Contribute to Large Upstream Migrations of Native Paddlefish and Invasive Bigheaded Carp. U.S. Geological Survey data release (2024). https://doi.org/10.5066/P14GQIVU.
Tyers, M. riverdist: River Network Distance Computation and Applications. R Package version 0.16.1. (2023).
U.S. Army Corps of Engineers. USACE St. Paul District—Water Management; https://www.mvp-wc.usace.army.mil/ (2024).
Minnesota Department of Natural Resources. Invasive Species of Minnesota—2023 Annual Report; https://files.dnr.state.mn.us/aboutdnr/reports/legislative/2024/2023-invasive-species-annual-report.pdf (2023).
Carlander, H. B. A History of Fish and Fishing in the Upper Mississippi River (Upper Mississippi River Conservation Committee, 1954).
Schramm, H. L. Jr. The fishery resources of the Mississippi River: A model for conservation and management. Fisheries 42, 574–585 (2017).
Brönmark, C. et al. There and back again: Migration in freshwater fishes. Can. J. Zool. 92, 467–479 (2014).
Chen, J. Evolutionary process of the Yangtze River. In Evolution and Water Resources Utilization of the Yangtze River (ed. Chen, J.) 47–122 (Springer, 2020).
Stell, E., Hoover, J. J., Fuller, L. & Parsons, G. R. Analyzing leap characteristics and water escape velocities of silver carp using in situ video analysis. N. Am. J. Fish. Manag. 40, 163–174 (2020).
Berdahl, A., Torney, C. J., Ioannou, C. C., Faria, J. J. & Couzin, I. D. Emergent sensing of complex environments by mobile animal groups. Science 339, 574–576 (2013).
Berdahl, A. M. et al. Collective animal navigation and migratory culture: From theoretical models to empirical evidence. Phil. Trans. R. Soc. B 373, 20170009 (2018).
Okasaki, C., Keefer, M. L., Westley, P. A. H. & Berdahl, A. M. Collective navigation can facilitate passage through human-made barriers by homeward migrating Pacific salmon. Proc. R. Soc. Lond. B 287, 20202137 (2020).
Bouska, K. L. Percentage of Annual Days that River Stage Exceeds ‘open river’ Conditions for Lock and Dams on the Upper Mississippi River, 1985–2015: U.S. Geological Survey data release (U.S. Geological Survey, 2021).
Southall, P. D. & Hubert, W. A. Habitat use by adult paddlefish in the upper Mississippi River. Trans. Am. Fish. Soc. 113, 125–131 (1984).
Moen, C. T., Scarnecchia, D. L. & Ramsey, J. S. Paddlefish movements and habitat use in Pool 13 of the upper Mississippi River during abnormally low river stages and discharges. N. Am. J. Fish. Manag. 12, 744–751 (1992).
Tripp, S. J. et al. Sturgeon and paddlefish migration: Evidence to support the need for interjurisdictional management. Fisheries 44, 183–193 (2019).
Bergman, J. N. et al. Historical, contemporary, and future perspectives on a coupled social–ecological system in a changing world: Canada’s historic Rideau Canal. Environ. Rev. 30, 72–87 (2022).
Minnesota Department of Natural Resources. Minnesota Invasive Carp Action Plan; https://files.dnr.state.mn.us/natural_resources/invasives/aquaticanimals/invasivecarp/invasive-carp-action-plan-2024.pdf (2024).
Newbold, L. R., Shi, X., Hou, Y., Han, D. & Kemp, P. S. Swimming performance and behaviour of bighead carp (Hypophthalmichthys nobilis): Application to fish passage and exclusion criteria. Ecol. Eng. 95, 690–698 (2016).
Hoover, J. J., Zielinski, D. P. & Sorensen, P. W. Swimming performance of adult bighead carp Hypophthalmichthys nobilis (Richardson, 1845) and silver carp H. molitrix (Valenciennes, 1844). J. Appl. Ichthyol. 33, 54–62 (2017).
Zielinski, D. P., Voller, V. R. & Sorensen, P. W. A physiologically inspired agent-based approach to model upstream passage of invasive fish at a lock-and-dam. Ecol. Model. 382, 18–32 (2018).
Byun, K. & Hamlet, A. F. Projected changes in future climate over the Midwest and Great Lakes region using downscaled CMIP5 ensembles. Int. J. Climatol. 38, e531–e553 (2018).
Acknowledgements
We thank Levi Suchla, Wes Bouska, and the commercial fishers for assisting with fish collection. Funding sources were the U.S. Geological Survey Ecosystems Mission Area Biological Threats and Invasive Species Research Program and U.S. Fish and Wildlife Service. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Author information
Authors and Affiliations
Contributions
M.F., D.G.R., and A.F. conceived the study; M.F., C.H., K.L., A.M., M.B., J.L., D.T., Z.W., E.S., G.L., J.S., K.Z., D.O., R.J.H., and A.F. designed the fieldwork and participated in field sampling; D.G.R., D.A., and A.F. analyzed the data; M.F., D.G.R., D.A., and A.F. wrote the original manuscript text; D.G.R. and D.A. prepared figures; all authors contributed to manuscript editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Fritts, M., Gibson-Reinemer, D., Appel, D. et al. Flooding and dam operations facilitate rapid upstream migrations of native and invasive fish species on a regulated large river. Sci Rep 14, 20609 (2024). https://doi.org/10.1038/s41598-024-70076-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-70076-4
- Springer Nature Limited