
Abstract
Background
Effects of confounders on associations between diet and colorectal cancer (CRC) in observational studies can be minimized in Mendelian randomization (MR) approach. This study aimed to investigate observational and genetically predicted associations between dietary intake and CRC using one-sample MR.
Methods
Using genetic data of over 93 million variants, we performed a genome-wide association study to find genomic risk loci associated with dietary intake in participants from the UK Biobank. Then we calculated genetic risk scores of diet-related variants and used them as instrumental variables in the two-stage least square MR framework to estimate the hazard ratios (HRs) and 95% confidence intervals (CIs) for associations. We also performed observational analyses using age as a time-scale in Cox proportional hazard models.
Results
Allele scores were calculated from 399 genetic variants associated with the consumption of of red meat, processed meat, poultry, fish, milk, cheese, fruits, vegetables, coffee, tea, and alcohol in participants from the UK Biobank. In MR analysis, genetically predicted fruit intake was significantly associated with a 21% decreased risk of CRC (HR = 0.79, 95% CI = 0.66–0.95), and there was a marginally inverse association between vegetable intake and CRC (HR = 0.85, 95% CI = 0.71–1.02). However, null findings were observed in multivariable analysis, with HRs (95% CIs) of 0.99 (0.98–1.01) and 0.99 (0.98–1.00) per increment of daily servings of fruits and vegetables, respectively.
Conclusion
Dietary habits were attributable to genetic variations, which can be used as instrumental variables in the MR framework. Our study supported a causal relationship between fruit intake and a decreased risk of CRC and suggested an effective strategy of consuming fruits in the primary prevention of CRC.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
With a global burden of 1.9 million new cases and 0.9 million deaths estimated in 2020, colorectal cancer (CRC) is the third most common cancer type and the second most common cancer death due to this malignancy in the world [1]. Regarding the prevention of CRC, the World Cancer Research Fund/American Institute for Cancer Research (WCRF/AICR) launched the guidance every 10 years based on up-to-date systematic reviews and meta-analyses and reported the level of evidence for the association of different dietary factors with CRC risk [2]. Observational studies may be vulnerable to residual confounding by factors that cannot be measured, and this may limit it in interpreting such an observed association as a causal relationship [3, 4]. In the meantime, by examining genetic variants such as single nucleotide polymorphisms (SNPs) as instrument variables (IVs) that act as proxies for environmental factors, Mendelian randomisation (MR) was suggested to provide a useful approach to minimise the bias of the effect estimate between risk factors and CRC risk [5,6,7].
A previous MR study comprehensively examined the causal inference of modifiable factors with the CRC risk [8]. Among 39 risk factors, only coffee consumption was included in the analysis due to unavailable or unsuitable SNPs for the use as instrumental variables (IVs) for other dietary factors [8]. Given a substantial proportion of the preference for foods was explained by genetic variations, individual food preferences and dietary habits have been identified to be affected by the senses of taste and smell and metabolic processes [9,10,11]. Additionally, a previous comprehensive genome-wide association study (GWAS) reported hundreds of significant loci for single foods and dietary patterns in participants of the UK Biobank [12]. However, underlying biological mechanisms contributing to genetic variations for the intake of several food items (e.g., pork vs. beef vs. lamb/mutton, oily vs. nonoily fish, fresh vs. dried fruits, cooked vs. raw vegetables) have been still unclear. Therefore, we first carried out a GWAS of food intake to identify genetic variants associated with the intake of total red meat, processed meat, poultry, total fish, milk, cheese, total fruits, total vegetables, coffee, tea, and alcohol, using updated data of more than double number of SNPs compared to the previous study. We then performed a one-sample MR study to elucidate the association between genetically predicted dietary intake and CRC risk using GWAS-identified genomic risk loci as IVs.
Materials and Methods
Study population
The UK Biobank is a prospective cohort study that included 502,389 participants aged 37–73 years who resided within 25 miles of 22 recruiting centers between 2006 and 2010. The study was approved by the North West Multi-centre Research Ethics Committee. The methodological details and rationale of the UK Biobank have been published elsewhere [13,14,15].
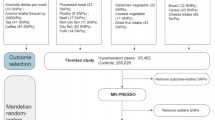
In the present study, we mutually excluded participants without genetic information (N = 15,208), sex mismatch (N = 367), putative sex chromosome aneuploidy (N = 651), and those who were either genetically identified or self-reported as having ethnic backgrounds other than White British (including White, British, Irish, and any other White backgrounds) (N = 78,378). After exclusion, the sample available for the genome-wide association analysis was restricted to 408,093 individuals. Finally, we excluded participants who were diagnosed with any cancers at enrolment (N = 34,078) and those who withdrew from the study during the follow-up (N = 11), leaving a total of 374,001 individuals (Fig. 1).

Flow diagrams of study participants and analytical framework
Genotyping and quality control
Genotyping was performed using either the custom UK Biobank Axiom Array or the Affymetrix Axiom Array, as described elsewhere [14, 15]. Genotyping data were imputed using both the UK10K and 1000 Genomes Phase 3 and the Haplotype Reference Consortium reference panel, which resulted in a total of 93,095,623 markers [14]. Following the quality control procedure, we excluded SNPs with low imputation quality (imputed score < 0.3, n = 15,368,777), high missingness (geno > 0.05, n = 909,502), low minor allele frequency (maf < 0.0002, n = 55,398,429) and those that deviated from the expected Hardy–Weinberg equilibrium (p < 1e-6, n = 8,717,604) [16]. A total of 27,503,596 SNPs that passed the quality filtering remained.
Dietary intake assessment
A touchscreen food frequency questionnaire (FFQ) was used to assess food and beverage intake in the preceding year [17]. Details of the questionnaire were publicly available [18]. In this study, we included foods that were documented in the WCRF report for their associations with CRC risk at various levels of evidence. We also selected foods for which consumption could reasonably be attributed to genetic variations (Additional file 1: eInformation). A linear mixed model was applied to adjust for familial relatedness in genome-wide association analysis of food intake; thus, we converted dietary outcomes into quantitative traits (Table 1). Of these, frequency traits of beef, pork, lamb, processed meat, poultry, oily fish, nonoily fish, cheese, and alcohol intake, and quantitative traits of fresh and dried fruits, cooked and raw vegetables, and coffee and tea consumption were included in our analyses. For categorical phenotypes, we used the corresponding numeric values (times/week) for the analysis. To justify the selection of dietary factors, we combined food items into more common food groups which are similar to those from the WCRF report. We grouped single items to obtain the total intake of red meat (including pork, beef, and lamb), total fish (including oily and nonoily fish), total fruits (including fresh and dried fruits), and total vegetables (including cooked and raw vegetables) [19]. Milk consumption (mL/day) was estimated based on the type of milk, breakfast cereal, coffee, and tea intake [19]. The 24-h dietary data were used to validate the estimation of milk intake, and 94% of the total milk consumption was found to come from milk added to breakfast cereal, coffee, and tea [19]. Overall, the Shapiro–Wilk test was applied to assess the normality of the data, and for data not following a normal distribution, the median and interquartile range was reported for data that did not follow normal distribution.
Outcome ascertainment
Incident CRC cases were determined via the ICD-10 code, in which CRC was defined as either colon cancer (C18.0-C18.9) or rectal cancer (C19 and C20). Time to follow-up was defined as the date of study enrolment until the date of CRC diagnosis, death, lost-to-follow-up, or end of follow-up (June 25, 2021), whichever came first.
Instrumental variables for dietary phenotype
To identify genetic variants associated with dietary traits, we performed a GWAS for food intake (Additional file 1: eMethod). In brief, we performed a genome-wide association analysis under the linear mixed model approach [20]. We incorporated age, sex, and the first 6 first principal component scores released by the UK Biobank [14] as covariates. In the large-scale UK Biobank dataset, more than 30% of study participants were genetically defined to relate with another participant [14]. Therefore, we further adjusted for the cryptic relatedness among participants by calculating the sparse genetic relatedness matrix (GRM) using genotyping data of 93,183 SNPs, which were used for the final kinship inference of the released UK Biobank data [14, 21, 22]. The list of genomic risk loci and their functions were determined under the SNP2GENE and GENE2FUNC functions of the web-based FUMA tool [23].
In sensitivity analysis, we excluded genetic variants, which were associated with more than two dietary traits from the list of IVs for dietary intake to minimise the possibility of horizontal pleiotropy. Additionally, for the exclusion restriction assumption, we further excluded SNPs that were associated with CRC risks (p-value < 0.05) from the list of candidate IVs to minimise the possibility of genetic variants affecting CRC other than through dietary intake. Details on the estimation of beta coefficients for the effect of variants on CRC risks adjusting for familial relatedness were available at Additional file 1: eMethod.
The internally weighted allele score for each participant was calculated by multiplying the number of effect alleles that the participant carried by the corresponding beta-coefficient of the association between the genetic variant and dietary intake estimated from the genome-wide association. Then we summed up the weighted allele score of individual genetic variants and used them as IVs in the MR analysis.
To assess the weak instrument problem, an F-statistic was implemented for IVs of allele scores and their corresponding individual genetic variants [24]. F-statistic was approximated by a squared estimate for IVs on dietary intake frequency divided by its variance.
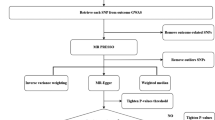
Mendelian randomisation analysis
We carried out a one-sample MR in the UKB to assess the effect of dietary intake on CRC using the two-stage least square method [25, 26]. In the first stage, we regressed each food frequency consumption on its respective allele score using a linear regression model to obtain a set of fitted values for exposure of interest. In the second stage, we regressed the CRC outcome on the fitted values obtained in stage 1 using an age-scale Cox proportional hazard model. Additionally, we used the MR pleiotropy residual sum and outlier test (MR-PRESSO) to detect the presence of pleiotropy [27] and the MR-Egger regression to identify whether directional pleiotropy may influence the causal estimates [28]. Subgroup analyses were conducted by sex and CRC subsites.
In sensitivity analysis, we carried out a multivariable MR, which included multiple dietary factors which their allele scores were substantially correlated or had relatively high genetic correlations.
Observational association
We sought to evaluate the association between dietary intake (in a continuous form) and CRC risk using age as a time-scale in Cox proportional hazard models. In the multivariable analysis, we adjusted for confounders, including sex, family history of CRC, household income, smoking, alcohol consumption (except for alcohol consumption exposure), body mass index, and physical activity, which were associated with CRC risk in the univariate analysis.
Results
Study population characteristics
Table 2 summarises the general characteristics and dietary habits of 174,576 men and 199,428 women without any cancers at enrolment. At recruitment, participants were aged 56.6 years (mean ages 56.5 years for men and 56.8 years for women). After a median follow-up of 12.4 years (interquartile range 11.6–13.1 years), 3,131 colon cancer and 1,555 rectal cancer cases were newly detected.
Loci and annotation of SNPs related to dietary intake
The results from the genome-wide association analysis for significant SNPs (p < 5 × 10–8) associated with food intake are presented as Manhattan plots (Fig. 2). We identified a total of 402 genomic risk loci for the consumption of red meat (n = 15), processed meat (n = 12), poultry (n = 1), total fish (n = 28), milk (n = 50), cheese (n = 59), total fruits (n = 82), total vegetables (n = 50), coffee (n = 33), tea (n = 40), and alcohol (n = 57) in the linear mixed model adjusting for familial relatedness (Additional file 2: Table S1). Of these, variants rs2199936 (chromosome 4, ABCG2 gene), rs139797380 (chromosome 6, SLC35D3 gene), and rs4410790 (chromosome 7, AC003075.4 gene) were associated with milk, coffee, and tea consumption. Variant 2:27,748,992 (chromosome 2, GCKR gene) was associated with the consumption of milk, coffee, and alcohol. Variant rs8103840 (chromosome 19, FUT1 gene) was associated with the intake of processed meat, fish, and fruits. In addition, some SNPs were associated with two dietary factors, including rs201406724 (milk and tea), rs11940694 (milk and alcohol), rs2465018 (milk and tea), rs17685 (milk and tea), rs4726481 (tea and alcohol), rs7012814 (cheese and tea), 8:73,433,232 (milk and tea), rs11032362 (processed meat and fruits), 12:11,271,915 (coffee and tea), rs12591786 (milk and tea), rs12909335 (milk and tea), rs9937521 (tea and alcohol), rs12459249 (milk and coffee), and rs429358 (fish and fruits).

Manhattan plot of genome-wide association analyses of A red meat, B processed meat, C poultry, D fish, E milk, F cheese, G fruit, H vegetable, I coffee, J tea, and K alcohol consumption using linear mixed model. X-axis shows chromosome positions, Y-axis shows -log10 of p-values. Red dashed lines indicate significant threshold (p = 5e-8)
Biological processes, molecular functions, and Wikipathways that may involve in insights into genetic effects on the intake of fish, milk, cheese, fruits, coffee, tea, and alcohol are presented in Additional file 2: Figures S1-S7. Overall, the heritability was highest for the consumption of cheese (h2 = 10.48%), alcohol (h2 = 9.71%), and milk (h2 = 9.01%), followed by tea (h2 = 8.34%) and fruits (h2 = 7.83%). Other foods had a heritability of approximately 5%-6%, except poultry (h2 = 3.50%) (Additional file 2: Table S2). Furthermore, we found a relatively high genetic relationship for the intake between milk and tea (r = 0.86), fish and vegetables (r = 0.52), fruits and vegetables (r = 0.49), red meat and processed meat (r = 0.48), processed meat and fruits (r = -0.46), cheese and alcohol (r = 0.44), and red meat and poultry (r = 0.43) (Additional file 2: Figure S8). The highest Pearson correlation coefficients between food consumption were found for coffee and tea (r = -0.32) and milk and tea (r = 0.30) (Additional file 2: Figure S8).
Mendelian randomisation analysis of dietary intake and colorectal cancer risk
All genetic instruments of SNPs and allele scores predicted dietary intake frequency, with F-statistics greater than 10, are presented in Tables 3, Additional file 2: Tables S1 and S3. Since only one variant was associated with poultry intake, we did not calculate the MR estimate for the effect of poultry intake on CRC.
Table 4 shows the estimates of the causal effect of dietary intake on CRC risks in the one-sample MR approach using the full lists of genetic variants. Overall, genetically proxied fruit intake was associated with 21% decreased risks of both CRC (HR = 0.79, 95% CI = 0.66–0.95) and colon cancer (HR = 0.79, 95% CI = 0.63–0.99). Findings for other dietary factors were not significant: red meat (HR = 0.72, 95% CI = 0.40–1.28), processed meat (HR = 0.57, 95% CI = 0.29–1.11), fish (HR = 1.05, 95% CI = 0.72–1.53), milk (HR = 1.19, 95% CI = 0.86–1.63), cheese (HR = 0.98, 95% CI = 0.78–1.23), coffee (HR = 1.16, 95% CI = 0.96–1.40), tea (HR = 0.95, 95% CI = 0.82–1.11), and alcohol (HR = 1.01, 95% CI = 0.86–1.20). These associations remained after excluding genetic variants associated with more than one dietary phenotype or related to CRC risks (Additional file 2: Table S4). In sex-specific subgroups, CRC reduction was only observed in women for an increment of 1 serving/day of consuming fruits in both the main analysis of including all eligible variants and the sensitivity analysis of the reduced list of variants, with HRs (95% CIs) of 0.72 (0.53–0.98) and 0.69 (0.50–0.96), respectively. Furthermore, genetically proxied alcohol consumption was associated with a 22% increased risk of CRC in men. However, this association disappeared in the sensitivity analysis using the reduced list of variants.
Marginally inverse associations were found for vegetable intake and CRC (HR = 0.85, 95% CI = 0.71–1.02) and colon cancer (HR = 0.80, 95% CI = 0.64–1.01) risks. Using genetic variants associated with a single dietary phenotype and not related to CRC, the magnitude of associations was similar to that of all eligible variants, with HRs (95% CIs) of 0.84 (0.70–1.01) and 0.80 (0.63–1.01) for CRC and colon cancer, respectively.
In the sensitivity analysis of using multivariable MR with the inclusion of multiple dietary factors which their allele scores were substantially correlated (r > 0.10, Additional file 2: Figure S9A) or had relatively high genetic correlations (r > 0.30, Additional file 2: Figure S9B), the sets of red meat and processed meat; fish, total fruit, and total vegetables; milk, tea, and coffee; and cheese and alcohol were considered in the model. Accordingly, genetically predicted consumption of red meat, processed meat, and cheese was associated with an increased risk of CRC, with HRs (95% CIs) of 1.30 (1.19–1.43), 1.29 (1.18–1.41), and 1.36 (1.21–1.53), respectively (Additional file 2: Table S9). Furthermore, inverse associations were observed for associations between genetically predicted vegetable (HR = 0.94, 95% CI = 0.90–0.98) and tea (HR = 0.97, 95% CI = 0.95–0.99) consumption (Additional file 2: Table S9).
Evaluation of pleiotropy effects
Although MR-PRESSO global tests suggested a possible bias from horizontal pleiotropy in associations of processed meat intake in men and coffee consumption in women with rectal cancer (Table 3, ppleiotropy = 0.01), the estimates after correcting for outliers remained in similar directions of associations, with HRs (95% CIs) of 0.30 (0.03–3.21) and 0.72 (0.35–1.47), respectively. The MR-PRESSO distortion test showed that the distortion in the effect estimates before and after removing outliers was not significant. These possible pleiotropy effects disappeared in our sensitivity analysis of restricting genetic variants for IVs (Additional file 2: Table S4).
Observational association
Additional file 2: Table S5 shows the observational effect of dietary intake on the risk of CRC. Red meat (HR = 1.05, 95% CI = 1.03–1.07, per 1 time/week), processed meat (HR = 1.03, 95% CI = 1.01–1.05, per 1 time/week), and alcohol consumption (HR = 1.03, 95% CI = 1.01–1.04, per 1 time/week) were positively associated with CRC risks. In contrast, more frequently milk (HR = 0.95, 95% CI = 0.92–0.97, per 100 mL/day) and tea (HR = 0.98, 95% CI = 0.97–0.99, per 1 cup/day) consumers had decreased risk of CRC. However, null associations were observed in multivariable analysis, with HRs (95% CIs) of 0.99 (0.98–1.01) and 0.99 (0.98–1.00) per increment of daily servings of fruits and vegetables, respectively. When stratified by sex, the effects of red meat, processed meat, and alcohol consumption remained for the men subgroup, whereas only the inverse association between milk intake and CRC risk was observed in women. Nevertheless, null findings were observed in multivariable analysis, with HRs (95% CIs) of 0.99 (0.98–1.01) and 0.99 (0.98–1.00) per increment of daily servings of fruits and vegetables, respectively.
In the analysis by CRC subsites, positive associations of red meat intake and inverse associations of milk and tea consumption were observed with both colon cancer and rectal cancer (Additional file 2: Tables S6-S7). Furthermore, processed meat (HR = 1.03, 95% CI = 1.01–1.05, per 1 time/day) and alcohol (HR = 1.03, 95% CI = 1.01–1.04, per 1 time/day) consumption showed an increased risk of colon cancer.
Discussion
In this study, we identified 399 genomic risk loci for self-reported traits reflecting daily consumption of food items included in the WCRF report for CRC prevention (Additional file 2: Figure S10). Using these genomic risk loci in the one-sample MR framework, we found that genetically predicted dietary intake of fruits was associated with a lower risk of CRC, with a similar magnitude of an inverse association with colon cancer. Additionally, marginally inverse associations between vegetable intake with CRC and colon cancer were observed in the total study population. When compared with our observational analysis of a prospective cohort study design, these associations appeared to be weaker and did not reach the level of significance (Additional file 2: Figure S11).
When we searched PubMed up to September 2023 for the GWAS of dietary traits, a total of 23 GWAS were identified, and seven studies included the population of the UK Biobank (Additional file 2: Table S8). Our study extended to the previous research by accounted for familial relatedness, which was not adjusted in most previous GWAS. Besides, to justify the selection of dietary factors, we combined food items into more common food groups that underlying biological mechanisms contributing to genetic variations existed. In addition, we analysed updated data with more than double SNPs from the most comprehensive GWAS for dietary intake [12]. Moreover, we carried out functional analyses to inform possible biological mechanisms between genetic factors and food consumption. A detailed comparison of the identified variants and the heritability of genetic factors between our present GWAS and Cole’s study is further provided in Additional file 3: Appendix.
By obtaining dietary habits from the questionnaire, we considered the amount of food consumption in the continuous form and applied the linear mixed model. A previous study converted food-liking traits into numerical values (range 0–9) without justification [29]. Given the transformation of food preference phenotypes into the hedonic scale into numeric values is not appropriate, the proportional odds logistic mixed model (POLMM) has been shown to handle ordinal categorical phenotypes, especially when the phenotype is extremely imbalanced [30]. The authors applied the POLMM for the frequent consumption of food items (never or almost never, once every few months, once a month, once a week, 2–4 times per week, and almost daily) in the UK Biobank without converting into numeric values [30]. In our present study, modelling dietary intake frequencies as continuous variables may violate the assumption of linearity relationship between SNPs and food consumption due to the restriction of outcome variable ranges. Nevertheless, findings on the top 10 genes were similar to those identified from our current study (e.g., CCDC171 for beef, pork, and lamb, XKR6 for processed meat, LY6H for poultry, and MLLT10 for oily fish).
The anti-cancer effects of fruits and vegetables were suggested due to their bioactive compounds, such as fiber, folate, vitamins, minerals, and flavonoids [31]. Of these, fiber is fermented by several bacteria to produce short-chain fatty acids (SCFAs), including acetate (central appetite regulation), propionate (gluconeogenesis and satiety signaling regulation), and butyrate (a main energy source for human colonocytes) [32, 33]. Higher fiber intake was associated with the increase of SCFAs, and SCFA-producing bacteria, which regulate the immune system and metabolism and reduce the CRC risk [33]. According to the WCRF/AICR, there was limited evidence for the effect of fruit and non-starchy vegetable intake on CRC prevention [34]. According to pooled estimates from prospective cohort studies, per daily 100 g of fruit and vegetable intakes were associated with a decreased risk of CRC by 4% (relative risk (RR) 0.96, 95% CI = 0.93–0.99) and 2% (RR = 0.98, 95% CI = 0.96–0.99), respectively [35]. However, individual studies tended to show null associations. A previous case–control analysis of nine observational studies within the Genetics Epidemiology of Colorectal Cancer Consortium and the Colon Cancer Family Registry did not observe any significant associations between fruit (odds ratio (OR) 1.04, 95% CI = 0.93–1.15) and vegetable (OR = 0.92, 95% CI = 0.82–1.03) intakes with overall CRC risk [36]. Similarly, nonsignificant associations between fruit (HR = 1.00, 95% CI = 0.94–1.05) and vegetable (HR = 1.01, 95% CI = 0.93–1.11) intakes and CRC risks were recently reported in a prospective cohort analysis of the UK Biobank [19]. These inconsistent findings with our MR estimates may be partly due to differences in study design and analytical framework. In general, observational studies are more prone to residual confounding, reverse causation, and measurement error than MR analyses, which randomly assign the exposure of interest-related IVs among individuals [4, 26]. Such sources of bias may attenuate associations toward the null [4, 26]. Furthermore, while the MR estimates reflect the effect of lifelong perturbations in risk factors, observational results may reflect more acute effects, during the follow-up period since the enrolment time point of a cohort) [37]. Our present observational analysis with a longer follow-up period (12.4 vs. 5.7 years) suggested stronger favorable effects of fruits (HR = 0.99, 95% CI = 0.91–1.01) and vegetables (HR = 0.99, 95% CI = 0.98–1.00), thus supports the evidence of long-term beneficial effects [19].
Among dietary factors, the International Agency for Research on Cancer classified processed meat as a human carcinogen (Group 1) and red meat as a probable carcinogen (Group 2A) [38]. Carcinogenic effects of red meat and processed meat were introduced via several chemicals such as N-nitroso compounds, heterocyclic aromatic amines, and polycyclic aromatic hydrocarbons formed in red meat and when cooking meat at high temperatures [39]. The WCRF/AICR also reported probable to convincing evidence of red meat and processed meat intake in association with CRC risks [34]. However, our present study observed the association between red meat and processed meat with CRC risk in observational analyses and multivariable MR. Besides differences in study design and analytical framework, the explained variation of IVs for the exposure of interest may affect our estimates. Although the allele score IVs explained variations of dietary intake (F-statistics greater than 90), the number of SNPs used for the calculation of allele scores for red meat and processed meat was relatively small, which may not allow us to detect any significant associations. We further observed an inverse association between processed meat intake and rectal cancer risk. These findings disappeared in sex-specific subgroups and need to be interpreted cautiously, possibly due to the small proportion of rectal cancer cases among whole study participants.
To date, very few MR studies reported the effect of dietary factors on CRC risk. Most of them considered blood concentrations of nutrients (carotenoids, calcium, copper, fatty acids, folate, iron, magnesium, methionine, phosphorus, selenium, sodium, vitamin B6, vitamin B12, vitamin D, vitamin E, and zinc) as exposure of interest [8, 40,41,42,43,44]. Only the MR study conducted by Cornish et al. examined the causal estimate between diet consumption of coffee and CRC risk. Although we used much more SNPs in the allele score calculation, our study revealed a similar direction of the estimates (33 SNPs, HR = 1.16, 95% CI = 0.96–1.40 in the current study vs. 4 SNPs, OR = 1.17, 95% CI = 0.88–1.55 in the previous study) [8].
Furthermore, we found inconclusive evidence of the MR estimates of total fish, milk, cheese, coffee, tea, and alcohol consumption on CRC. Of these, pooled estimates from observational studies showed significantly or suggestively inverse associations of fish (RR = 0.89, 95% CI = 0.80–0.99), milk (RR = 0.94, 95% CI = 0.92–0.96), cheese (RR = 0.94, 95% CI = 0.87–1.02), coffee (RR = 1.00, 95% CI = 0.99–1.02), tea (RR = 0.99, 95% CI = 0.97–1.01), and alcohol (RR = 1.07, 95% = 1.05–1.08) intake with CRC risk [35]. Compared to observational analysis, estimates from MR may commonly have wider CIs and thus toward null findings [37].
This study has several strengths. Having large-scale individual-level data with much more genetic information of imputed SNPs compared to earlier GWAS, we applied the recent methodology to account for confounding effects of both population stratification and cryptic relatedness to identify loci associated with food intake. We also performed a comprehensive MR analysis to suggest evidence for the causal estimate of dietary intake and CRC risk. Genetic variants had adequate strengths; thus, bias due to small F-statistics or small sample size can be minimised. Undertaking sensitivity analyses to evaluate the plausibility of IV assumptions and robustness to pleiotropy and outliers, our findings from MR analyses may be less biased by residual confounding and reverse causation than observational results. Additionally, combining many SNPs into a single allele score may increase the power of the analysis and reduce the risk of bias from possible weak instruments [26]. Furthermore, available data for one-sample MR analysis allowed us to consider the effect estimate in several subgroups, such as sex and CRC subsites.
Despite providing new evidence about the causal effect of dietary intake on CRC risk, this study has some limitations that need to be addressed. One limitation of the study is the fact that we analysed CRC risk only using the dietary information measured at a single time point, which may not reflect the lifelong dietary intake, thus, our findings were based on the assumption that such dietary habits might not change or be equally changed during follow-up. The effect of dietary factors might be underestimated due to random measurement errors [45]. Previous study investigated the reproducibility of the touchscreen questionnaire of average diet over the previous 12 months used in the current study with the 24-h dietary assessment [45]. Overall, the intra-correlation of food groups was reported to range between 0.38 to 0.63, which was comparable with the overall reproducibility of FFQs in nutritional epidemiology studies (macronutrients: 0.44–0.79; micronutrients: 0.51–0.74) [46]. However, among all participants completed the touchscreen questionnaire, only approximately 42% study participants provided the 24-h dietary assessment [45]. Nevertheless, our findings were limited for 24-h dietary data. Besides, given that disparities in dietary intake according to different ethnic groups may exist due to cultural knowledge and food-related skills [47, 48], analyses for individuals from ethnic backgrounds other than White British require additional investigations. Furthermore, we derived SNPs and weights for IVs in all participants after quality control and performed the two-stage least square analysis in participants without any cancer at baseline. There could still be a winner’s curse on our estimate due to the overlap between the dataset in which genetic variants were selected and the dataset in which genetically predicted associations were determined [49]. However, the winner’s curse bias in our study can be mitigated by selecting more stringent SNPs based on not only significant threshold but also linkage disequilibrium among variants. Moreover, to obtain GWAS-identified variants for the MR analysis, our study assumed linear associations between dietary intake and risk of developing CRC.
Conclusion
In summary, the present study comprehensively assessed the influence of genetic variants and their functional mechanisms on the dietary behaviors of participants in the UK Biobank. By cautiously accounting for population stratification and cryptic relatedness in this large-scale of recently released imputation data, we identified several loci for food consumption. These genetic variants associated were used as IVs in the MR framework to address the relationship between dietary intake and CRC risk. Our findings supported a relationship between fruit intake and a decreased risk of CRC and suggested an effective strategy of consuming fruits in the primary prevention of CRC. Further studies in individuals from ethnic backgrounds other than White British are needed to validate our findings.
Availability of data and materials
The UK Biobank is an open access resource, available at https://www.ukbiobank.ac.uk/researchers/. Data used in this project can be obtained from the UK Biobank by submitting a data request proposal.
Abbreviations
- GWAS:
-
Genome-wide association study
- MR:
-
Mendelian randomisation
- IV:
-
Instrumental variable
- WCRF:
-
World Cancer Research Fund
- AICR:
-
American Institute for Cancer Research
- FFQ:
-
Food frequency questionnaire
- SNP:
-
Single nucleotide polymorphism
- POLMM:
-
proportional odds logistic mixed model
- HR:
-
Hazard ratio
- RR:
-
Relative risk
- CI:
-
Confidence interval
- PC:
-
Principal component
References
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–49.
Clinton SK, Giovannucci EL, Hursting SD. The World Cancer Research Fund/American Institute for Cancer Research third expert report on diet, nutrition, physical activity, and cancer: impact and future directions. J Nutr. 2020;150(4):663–71.
Benn M, Nordestgaard BG. From genome-wide association studies to Mendelian randomization: novel opportunities for understanding cardiovascular disease causality, pathogenesis, prevention, and treatment. Cardiovasc Res. 2018;114(9):1192–208.
Wade KH, Yarmolinsky J, Giovannucci E, Lewis SJ, Millwood IY, Munafo MR, Meddens F, Burrows K, Bell JA, Davies NM, et al. Applying Mendelian randomization to appraise causality in relationships between nutrition and cancer. Cancer Causes Control. 2022;33(5):631–52.
Markozannes G, Kanellopoulou A, Dimopoulou O, Kosmidis D, Zhang X, Wang L, Theodoratou E, Gill D, Burgess S, Tsilidis KK. Systematic review of Mendelian randomization studies on risk of cancer. BMC Med. 2022;20(1):41.
Jung SY, Papp JC, Sobel EM, Zhang ZF. Mendelian randomization study: the association between metabolic pathways and colorectal cancer risk. Front Oncol. 2020;10: 1005.
Mao Y, Yan C, Lu Q, Zhu M, Yu F, Wang C, Dai J, Ma H, Hu Z, Shen H, et al. Genetically predicted high body mass index is associated with increased gastric cancer risk. Eur J Hum Genet. 2017;25(9):1061–6.
Cornish AJ, Law PJ, Timofeeva M, Palin K, Farrington SM, Palles C, Jenkins MA, Casey G, Brenner H, Chang-Claude J, et al. Modifiable pathways for colorectal cancer: a Mendelian randomisation analysis. Lancet Gastroenterol Hepatol. 2020;5(1):55–62.
Smith AD, Fildes A, Cooke L, Herle M, Shakeshaft N, Plomin R, Llewellyn C. Genetic and environmental influences on food preferences in adolescence. Am J Clin Nutr. 2016;104(2):446–53.
Boesveldt S, de Graaf K. The differential role of smell and taste for eating behavior. Perception. 2017;46(3–4):307–19.
Vesnina A, Prosekov A, Kozlova O, Atuchin V. Genes and eating preferences, their roles in personalized nutrition. Genes (Basel). 2020;11(4):357.
Cole JB, Florez JC, Hirschhorn JN. Comprehensive genomic analysis of dietary habits in UK Biobank identifies hundreds of genetic associations. Nat Commun. 2020;11(1):1467.
Canela-Xandri O, Rawlik K, Tenesa A. An atlas of genetic associations in UK Biobank. Nat Genet. 2018;50(11):1593–9.
Bycroft C, Freeman C, Petkova D, Band G, Elliott LT, Sharp K, Motyer A, Vukcevic D, Delaneau O, O’Connell J, et al. The UK Biobank resource with deep phenotyping and genomic data. Nature. 2018;562(7726):203–9.
Sudlow C, Gallacher J, Allen N, Beral V, Burton P, Danesh J, Downey P, Elliott P, Green J, Landray M, et al. UK Biobank: an open access resource for identifying the causes of a wide range of complex diseases of middle and old age. PLoS Med. 2015;12(3): e1001779.
Orliac EJ, Trejo Banos D, Ojavee SE, Lall K, Magi R, Visscher PM, Robinson MR. Improving GWAS discovery and genomic prediction accuracy in biobank data. Proc Natl Acad Sci U S A. 2022;119(31):e2121279119.
Bradbury KE, Young HJ, Guo W, Key TJ. Dietary assessment in UK Biobank: an evaluation of the performance of the touchscreen dietary questionnaire. J Nutr Sci. 2018;7: e6.
Data field 113241: Touchscreen questionnaire ordering, validation and dependencies. https://biobank.ndph.ox.ac.uk/ukb/refer.cgi?id=113241.
Bradbury KE, Murphy N, Key TJ. Diet and colorectal cancer in UK Biobank: a prospective study. Int J Epidemiol. 2020;49(1):246–58.
Jiang L, Zheng Z, Qi T, Kemper KE, Wray NR, Visscher PM, Yang J. A resource-efficient tool for mixed model association analysis of large-scale data. Nat Genet. 2019;51(12):1749–55.
Thomson R, McWhirter R. Adjusting for familial relatedness in the analysis of GWAS data. Methods Mol Biol. 2017;1526:175–90.
Yang J, Benyamin B, McEvoy BP, Gordon S, Henders AK, Nyholt DR, Madden PA, Heath AC, Martin NG, Montgomery GW, et al. Common SNPs explain a large proportion of the heritability for human height. Nat Genet. 2010;42(7):565–9.
Watanabe K, Taskesen E, van Bochoven A, Posthuma D. Functional mapping and annotation of genetic associations with FUMA. Nat Commun. 2017;8(1):1826.
Stock JH, Wright JH, Yogo M. A survey of weak instruments and weak identification in generalized method of moments. J Bus Econ. 2002;20(4):518–29.
Burgess S, Small DS, Thompson SG. A review of instrumental variable estimators for Mendelian randomization. Stat Methods Med Res. 2017;26(5):2333–55.
Sanderson E, Glymour MM, Holmes MV, Kang H, Morrison J, Munafo MR, Palmer T, Schooling CM, Wallace C, Zhao Q, et al. Mendelian randomization. Nat Rev Methods Primers. 2022;2:6.
Verbanck M, Chen CY, Neale B, Do R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat Genet. 2018;50(5):693–8.
Bowden J, Davey Smith G, Burgess S. Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression. Int J Epidemiol. 2015;44(2):512–25.
May-Wilson S, Matoba N, Wade KH, Hottenga JJ, Concas MP, Mangino M, Grzeszkowiak EJ, Menni C, Gasparini P, Timpson NJ, et al. Large-scale GWAS of food liking reveals genetic determinants and genetic correlations with distinct neurophysiological traits. Nat Commun. 2022;13(1):2743.
Bi W, Zhou W, Dey R, Mukherjee B, Sampson JN, Lee S. Efficient mixed model approach for large-scale genome-wide association studies of ordinal categorical phenotypes. Am J Hum Genet. 2021;108(5):825–39.
Song M, Garrett WS, Chan AT. Nutrients, foods, and colorectal cancer prevention. Gastroenterology. 2015;148(6):1244-1260 e1216.
Valdes AM, Walter J, Segal E, Spector TD. Role of the gut microbiota in nutrition and health. BMJ. 2018;361: k2179.
Song M, Chan AT, Sun J. Influence of the gut microbiome, diet, and environment on risk of colorectal cancer. Gastroenterology. 2020;158(2):322–40.
Continuous update project expert report 2018. Diet, nutrition, physical activity and colorectal cancer. https://www.wcrf.org/diet-activity-and-cancer/.
Papadimitriou N, Markozannes G, Kanellopoulou A, Critselis E, Alhardan S, Karafousia V, Kasimis JC, Katsaraki C, Papadopoulou A, Zografou M, et al. An umbrella review of the evidence associating diet and cancer risk at 11 anatomical sites. Nat Commun. 2021;12(1):4579.
Hidaka A, Harrison TA, Cao Y, Sakoda LC, Barfield R, Giannakis M, Song M, Phipps AI, Figueiredo JC, Zaidi SH, et al. Intake of dietary fruit, vegetables, and fiber and risk of colorectal cancer according to molecular subtypes: a pooled analysis of 9 studies. Cancer Res. 2020;80(20):4578–90.
Davies NM, Holmes MV, Davey Smith G. Reading Mendelian randomisation studies: a guide, glossary, and checklist for clinicians. BMJ. 2018;362: k601.
Agents classified by the IARC monographs, volumes 1–133. https://monographs.iarc.who.int/agents-classified-by-the-iarc/.
Turesky RJ. Mechanistic evidence for red meat and processed meat intake and cancer risk: a follow-up on the international agency for research on cancer evaluation of 2015. Chimia (Aarau). 2018;72(10):718–24.
Lu Y, Li D, Wang L, Zhang H, Jiang F, Zhang R, Xu L, Yang N, Dai S, Xu X, et al. Comprehensive investigation on associations between dietary intake and blood levels of fatty acids and colorectal cancer risk. Nutrients. 2023;15(3):730.
Feng Q, Wong SH, Zheng J, Yang Q, Sung JJ, Tsoi KK. Intake of processed meat, but not sodium, is associated with risk of colorectal cancer: evidence from a large prospective cohort and two-sample Mendelian randomization. Clin Nutr. 2021;40(7):4551–9.
Tsilidis KK, Papadimitriou N, Dimou N, Gill D, Lewis SJ, Martin RM, Murphy N, Markozannes G, Zuber V, Cross AJ, et al. Genetically predicted circulating concentrations of micronutrients and risk of colorectal cancer among individuals of European descent: a Mendelian randomization study. Am J Clin Nutr. 2021;113(6):1490–502.
Ong JS, Gharahkhani P, An J, Law MH, Whiteman DC, Neale RE, MacGregor S. Vitamin D and overall cancer risk and cancer mortality: a Mendelian randomization study. Hum Mol Genet. 2018;27(24):4315–22.
Dimitrakopoulou VI, Tsilidis KK, Haycock PC, Dimou NL, Al-Dabhani K, Martin RM, Lewis SJ, Gunter MJ, Mondul A, Shui IM, et al. Circulating vitamin D concentration and risk of seven cancers: Mendelian randomisation study. BMJ. 2017;359: j4761.
Carter JL, Lewington S, Piernas C, Bradbury K, Key TJ, Jebb SA, Arnold M, Bennett D, Clarke R. Reproducibility of dietary intakes of macronutrients, specific food groups, and dietary patterns in 211 050 adults in the UK Biobank study. J Nutr Sci. 2019;8: e34.
Cui Q, Xia Y, Wu Q, Chang Q, Niu K, Zhao Y. A meta-analysis of the reproducibility of food frequency questionnaires in nutritional epidemiological studies. Int J Behav Nutr Phys Act. 2021;18(1):12.
Mackenbach JD, Dijkstra SC, Beulens JWJ, Seidell JC, Snijder MB, Stronks K, Monsivais P, Nicolaou M. Socioeconomic and ethnic differences in the relation between dietary costs and dietary quality: the HELIUS study. Nutr J. 2019;18(1):21.
Wang Y, Chen X. How much of racial/ethnic disparities in dietary intakes, exercise, and weight status can be explained by nutrition- and health-related psychosocial factors and socioeconomic status among US adults? J Am Diet Assoc. 2011;111(12):1904–11.
Jiang T, Gill D, Butterworth AS, Burgess S: An empirical investigation into the impact of winner's curse on estimates from Mendelian randomization. Int J Epidemiol. 2023;52(4):1209–14.
Acknowledgements
We thank Professor Seunggeun Lee (Shawn), from Seoul National University Graduate School of Data Science, for his useful comments on this study.
Funding
This work was supported by the grant from the National Research Foundation of Korea (NRF) (No: 2022R1A2C1004608).
Author information
Authors and Affiliations
Contributions
TH made contributions to study conceptualization, data analysis, interpretation the results, and was a major contributor in writing the manuscript. AS and SC made contributions to study conceptualization and design, data interpretation, and revising the manuscript critically for intellectual content. JC and DK contributed to involved in revising the manuscript critically for intellectual content. All authors critically reviewed this manuscript and approved the final version to be published.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This research was conducted using the UK Biobank Resource (Application Number: 94695). The study protocol was approved by the Institutional Review Board of Seoul National University (No. 2101–153-1191). The current analysis was approved under UKB application #94695.
Consent for publication
Not applicable.
Competing interests
The corresponding author, Aesun Shin, is an Editorial Board Member (Associate Editor) of BMC Cancer. The other authors have no competing interests to declare.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hoang, T., Cho, S., Choi, JY. et al. Genetically predicted dietary intake and risks of colorectal cancer: a Mendelian randomisation study. BMC Cancer 24, 1153 (2024). https://doi.org/10.1186/s12885-024-12923-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12885-024-12923-1