
Abstract
Background
Peste des petits ruminants (PPR) disease is a cross-species infectious disease that severely affects small ruminants and causes great losses to livestock industries in various countries. Distinguishing vaccine-immunized animals from naturally infected animals is an important prerequisite for the eradication of PPR. At present PPRV are classified into lineages I through IV, and only one vaccination strain, Nigeria/75/1, belongs to lineage II, but all of the epidemic strains in China at present are from lineage IV.
Results
To achieve this goal, we developed an SYBR Green I real-time qRT-PCR method for rapid detection and identification of PPRV lineages II and IV by analyzing different melting curve analyses. The negative amplification of other commonly circulating viruses such as orf virus, goat poxvirus, and foot-and-mouth disease virus demonstrated that primers targeting the L gene of PPRV were extremely specific. The sensitivity of the assay was assessed based on plasmid DNA and the detection limit achieved was 100 copies of PPRV lineages II and IV.
Conclusion
Since the method has high sensitivity, specificity, and reproducibility, it will be effectively differentiated PPRV lineages II from PPRV lineages IV in PPRV infected animals.
Similar content being viewed by others


Background
Peste des petits ruminants (PPR) is a highly contagious, cross-species disease caused by the Peste des petits ruminants virus (PPRV) [1, 2]. It can infect not only small ruminants such as goats and sheep, but also large ruminants such as camels, buffalo, and wild animals [3, 4]. More than 68% of sheep and goats and 2.5 billion small ruminants reside in countries affected with PPRV, according to FAO data from 2018 [5]. The widespread distribution and prevalence of PPR have an important impact on economic trade and biodiversity.
PPRV belongs to the genus Morbillivirus of the family Paramyxoviridae [6]. According to the sequences of the N gene and F gene, PPRV can be classified into four lineages, I, II, III, and IV [7]. Lineages I and II are predominantly endemic in West Africa [8, 9], while lineage III is mainly found in the Arab region, and has also been reported in East Africa [10, 11], Lineage IV is mainly prevalent in the Middle East and Asia, and has also been reported in Africa [81213]. PPR was first reported in the Tibet region in 2007 [14], then re-emerged in Xinjiang Uyghur Autonomous Region in 2013, and then spread to 23 provinces, autonomous regions, and municipalities of China [1516]. China has adopted strict compulsory immunization and culling policies, which have effectively controlled the occurrence of the PPR. Currently, the live vaccine named the Nigeria/75/1 is the only vaccine widely used for the prevention of PPR in China. It can provide strong immune protection in goats and sheep, as it can proliferate continuously after injection. A major obstacle to eliminating PPR is the inability to distinguish vaccine groups from PPRV antibodies or antigen-positive animals in clinical settings. So, for the prevention and elimination of PPR, there is a great need for a simple assay that can detect and distinguish the PPRV vaccinated and infected groups at the same time.
Since the Nigeria/75/1 strain PPRV belongs to lineage II, while the PPRV prevalent in China all belong to lineage IV [15], and SYBR Green I real-time qRT-PCR assay was developed by analyzing all the published PPRV sequences of lineages II and IV. We use only one pair of primers that can detect all the lineages II and IV PPRV, and by analyzing different melting curve analyses (MCA), it can differentiate them conveniently and quickly. The parameters were optimized, and the sensitivity, specificity and reproducibility of the assay were assessed and compared with the traditional methods. The diagnostic application of the assay was carefully evaluated based on standard plasmid which cloned from the L gene of PPRV genome.
Results
Standard curve for the real-time qRT-PCR
Two linear standard curves were constructed using tenfold dilutions of PPRV lineages II and IV standard plasmids, using a dilution concentration of 1.0 × 108-1.0 × 102 copies/µL. All samples were tested in triplicate and independently. The equations for the PPRV lineages II and IV standard curves were y = -3.1734x + 31.044 and y = -3.1358x + 32.956, respectively. Standard curves analysis demonstrated that when the copy numbers were in the range of 108 and 102 copies/µL, the amplification efficiency of PPRV lineages II and IV was 106% and 108%, respectively. The correlation coefficient R2 value for the linear regression equation of PPRV lineages II and IV was 0.9981 and 0.9984 (Fig. 1A and B).
In addition, the melting curves for both lineages II and IV were single peaks and melting temperatures (Tm) were 83.39℃ and 81.92℃, respectively (Fig. 2).

Amplification curves and standard curves of the real-time qRT-PCR assay to measure viral copy number of PPRV lineages II and IV. The tenfold diluted standard plasmid of PPRV lineages II and IV was tested with the same pair of primers. Regression lines between the Ct values and the input concentrations of PPRV lineage II (A) and PPRV lineage IV (B) plasmid DNA using SYBR Green I real-time qRT-PCR

Melting curves of the real-time qRT-PCR assay to detect PPRV lineages II and IV. Amplification melting curve and melting temperature values (Tm) after SYBR Green I real-time qRT-PCR followed by melting curve analysis of PPRV lineages II and IV
Specificity, sensitivity and reproducibility of real-time qRT-PCR assay
To verify the specificity of the assay, lineage II PPRV, ORFV, GTPV, and FMDV genomic cDNA or DNA, and lineage IV PPRV plasmids were used as templates and amplified using the conditions described above. Only PPRV (lineages II and IV) was amplified in these samples and no specific peaks appeared in the other samples (Fig. 3), which indicated that the established real-time qRT-PCR assay was specific for PPRV and does not cross-react with other pathogens. The detection limits for PPRV lineages II and IV were 100 copies/µL.

Specificity of the real-time qRT-PCR assay. The specific fluorescent signals were detected from cDNA of PPRV lineage II and PPRV lineage IV plasmids, and the dissociation curves showed that there were two specific product peaks for PPRV lineages II and IV, respectively, but no specific amplification for negative control (ORFV, GTPV, and FMDV)
Standard plasmids (PPRV lineages II and IV) were diluted (107,105,103 copies/µL) to assess intra- and inter-assay reproducibility. And the CV of CT values were all less than 5%. Specifically, for PPRV lineage II plasmids, the CV of the inter- and intra- assays ranged from 0.06% to 0.49% and 0.07% to 6.09%, and for PPRV lineage IV plasmids, the CVs ranged from 0.06% to 0.30% and 1.03% to 6.49%, respectively (Table 1). These results indicate that the real-time qRT-PCR assay had good reproducibility.
Detection and identification of PPRV in test samples
To evaluate the assay’s potential performance in the clinical setting, we calculate the melting temperature in the real-time qRT-PCR target region of all the PPRV lineage II and IV strains (9 lineage II strains, 25 lineage IV strains isolated from foreign countries, and 37 lineage IV strains from China) (Table 2). The average, median, and standard deviation melting temperature of lineage II (IV) amplified area are 82.34℃ (80.97℃), 82.30 ℃ (81 ℃), and 0.19℃ (0.14℃), respectively. Furthermore, there was no significant difference between lineage IV strains isolated from China and foreign countries. The DNA denaturation temperature diversity between the two lineages is about 1.3℃ which can be easily reflected after MCA, it indicates this assay can efficiently detect and differentiate Nigeria/75/1 and PPRV lineage IV isolates when used in the future.
To evaluate the application of the method, total RNA from 16 transfected cell samples was extracted and reverse transcribed after digested with DNase I (Fig. 4A), and then detected by the established assay. The results showed that the method was able to distinguish the PPRV lineages II and IV genomic RNA clearly (Fig. 4B). Since the detection results of standard plasmids and RNA samples are highly consistent, it is suggested that this method can effectively amplify and distinguish clinical samples containing PPRV lineages II and IV.

Detection and identification of PPRV in test samples. A Total RNA of PPRV-L gene transfected cell samples was digested with DNase I. HEK-293T cells were transfected with lineages II and IV PPRV-L gene plasmids. Cell samples were collected at 12, 24, 36 and 48 h after transfection. Total RNA was extracted and digested with DNase I. B PPRV lineages II and IV were detected and differentiated in cell samples by the real-time qRT-PCR assay. After the RNA of PPRV-L gene transfected cell samples was completely digested by DNase I, it was reversed transcribed and detected by the established real-time qRT-PCR assay
Discussion
Since the emergence of PPRV in 1942, it has been widespread around the world, causing huge economic losses to the world’s animal husbandry. After the first report of PPRV in China in 2007, the outbreak of PPR still occurred, even though China adopted strict compulsory immunization and culling policies. At present, the vaccine strain is lineage II PPRV, but the dominant circulating lineage in China is IV. It is particularly important to detect and distinguish vaccine strain from wild strains.
At present, there are many detection methods for PPRV, which are mainly designed to detect all PPRV strains and can not distinguish between field PPRV strains and vaccine strains [17–19]. The real-time RT-PCR method designed by Li et al. [20] could specifically detect lineage IV PPRV, while the other three lineages could not be detected which limits of detection were found to be 100 copies/reaction. Yang established two detection methods for PPRV, real-time and lateral flow strip RT-RPA. The sensitivity of both was as low as 100 copies/reaction at 40℃ and as low as 150 copies/reaction at 39 ℃, respectively [21]. Several quantitative methods have been used to detect PPRV [22, 23], all of these methods can detect PPRV rapidly and effectively, but cannot distinguish between the PPRV vaccine strain and the prevalent strain in China.
The stability of the DNA duplexes can be affected by the GC content and length of the PCR products. Thus, the fluorescence signal of the SYBR Green I was different and could be used to specifically identify PCR products by Melting curve analysis (MCA). MCA in conjunction with real-time PCR, including dsDNA dye SYBR Green I and TaqMan probe-based types, has been widely used in previous studies including and single-nucleotide polymorphism(SNP)analysis [24–26]. The advantage of probe-based MCA is its ability for multiplex detection using a probe with specific melting temperatures. Compared with TaqMan probe-based MCA, SYBR Green I is a simple, rapid, and inexpensive method for testing. In our laboratory, PCR coupled with MCA has also been developed to detect and genotype duck hepatitis A virus types 1 and 3 [27]. In this study, we describe for the first time that only uses one pair of primers to distinguish lineage II and IV of PPRV based on the MCA and without reducing sensitivity. The sensitivity of the method is 100 copies/reaction. In addition, it does not cross-react with other viruses such as ORFV, GTPV and FMDV.
In order to evaluate the potential performance of the method in clinical application, first, the qRT-PCR target melting temperature of all PPRV lineages II and IV strains were calculated, and found the mean, median and standard deviation of the melting temperature of PPRV lineages II and IV amplified regions were significant differences, which were 82.34 °C (80.97 °C), 82.30 °C (81 °C) and 0.19 °C (0.14 °C), respectively. The denaturation temperature difference between PPRV lineages II and IV is about 1.3 °C, which can be easily distinguished by the melting curve. Then cell samples transfected with PPRV lineages II and IV plasmids were detected by the method and found that this method can effectively distinguish the two lineages of PPRV. In summary, the method can effectively detect and differentiate IV isolates of Nigeria/75/1 and PPRV families and has clinical application value.
Conclusion
In this study, we developed an MCA-based SYBR Green I RT-PCR assay to effectively detect and differentiate PPRV lineages II and IV. The detection method is accurate, rapid, sensitive, reproducible, and easy to perform. Therefore, this detection method can be used for subsequent epidemiological investigations and PPR eradication.
Methods
Virus strains
Lineage II PPRV strain PPRV Nigeria/75/1 (GenBank accession no. HQ197753), which was a vaccine strain and propagated in Vero-SLAM cells. Lineage IV PPRV strain PPRV-FY (GenBank accession no. KX354359) was isolated and preserved in our lab. Orf virus (ORFV) strain AH-FD/2016/China, which was preserved in Shanghai Veterinary Research Institute (SHVRI), Chinese academy of agricultural science. The goat poxvirus (GTPV) strain AV41 and the foot-and-mouth disease virus (FMDV) strain O/Mya98/BY/2010 both were vaccine strains.
Primer design
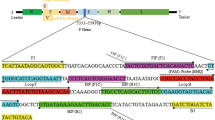
71 published whole-genome sequences (9 form lineage II PPRV and 62 form lineage IV PPRV, including 37 were Chinese strains and 25 were foreign strains) were retrieved from the GenBank database (Table 1). DNAstar software was used for comparison, and the L gene was used to design primers to detect and differentiate lineages II and IV PPRV, based on the principle that GC content difference in the inner region of two lineages PPRV gene sequences can produce different melting curves. The optimized forward primer F was 5’-ACAGGTTCGACAACATTCAAGCCA-3’ and reverse primer R was 5’-GCGAAGGTAGGTCAGAGAGCA-3’, and an expected 221 bp amplification product was generated.
Standard plasmids template preparation
Lineage II PPRV genomic RNA was extracted from the viral suspension with RNeasy (Qiagen, Germany), and used immediately for cDNA synthesis. cDNA synthesis was performed with SuperScript II reverse transcriptase (RT) (Invitrogen, USA) and specific RT primers (R). The cDNA fragment of lineage II PPRV and synthesized plasmid of lineage IV PPRV were amplified with primer F and R. The 221 bp products were cloned into the pMD19-T vector (TaKaRa) and sequenced by Sangon (Shanghai, China).
SYBR Green I-based quantitative real-time RT-PCR assay
The SYBR Green I-based real-time qRT-PCR was performed using Novozymes Ultra SYBR Mixture (Vazyme). Amplification was performed in a 20 µL reaction mixture containing 10 µL SYBR qPCR master mix (2×), 0.4 µL of each forward and reverse primers (10𝜇M), 2 𝜇L plasmid, and 7.2 µL H2O. The amplification conditions were 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 30 s. Two standard curves were generated with 10-fold serially diluted plasmids standards of 102-108 copies/µL. Each process was performed three times.
Specificity, sensitivity and reproducibility of the real-time qRT-PCR assay
To determine the specificity of the method, PPRV lineage II, ORFV, GTPV and FMDV, genomic cDNA or DNA, and PPRV lineage IV plasmids were used as templates in real-time qRT-PCR according to the aforementioned reaction conditions.
To determine the sensitivity of the method, ten 10-fold serial dilutions of recombinant plasmids with concentrations from 108 to 102 copies/µL were used as templates. Each experiment was performed three times.
Standard plasmids were chosen to evaluate the reproducibility of the method. Every dilution was evaluated for intra-assay and inter-assay variation. Each process was tested three times in parallel under the same amplification conditions. Real-time qRT-PCR was used to detect the coefficient of variation (CV) and to determine the repeatability of the inter-batch detection.
Evaluation of the real-time qRT-PCR assay
To evaluate the application of the method, lineages II and IV PPRV-L gene eukaryotic expression plasmids were constructed to simulate viral infection. HEK-293T cells were transfected with the lineages II and IV PPRV-L gene plasmids, then were collected at 12, 24, 36, and 48 h after transfection. Nucleic acids were extracted from these samples and tested by the real-time qRT-PCR.
Availability of data and materials
The data presented in this manuscript are available through the corresponding author upon reasonable request. The datasets analyzed in the study are available in the NCBI GenBank database, [lineage II PPRV Nigeria/75/1 (https://www.ncbi.nlm.nih.gov/nuccore/HQ197753), lineage IV PPRV-FY (https://www.ncbi.nlm.nih.gov/nuccore/KX354359)].
Abbreviations
- PPR:
-
Peste des petits ruminants
- PPRV:
-
Peste des petits ruminants virus
- qRT-PCR:
-
Quantitative reverse transcription polymerase chain reaction
- FAO:
-
Food and Agriculture Organization
- ORFV:
-
Orf virus
- GTPV:
-
Goat poxvirus
- FMDV:
-
Foot-and-mouth disease virus
- MCA:
-
Melting curve analyses
- Ct:
-
Cycle threshold
- CV:
-
Coefficients of variation
- Tm:
-
Melting temperatures
- HRM:
-
High-resolution melting
References
Abubakar M, Mahapatra M, Muniraju M, Arshed MJ, Khan EH, Banyard AC, Ali Q, Parida S. Serological detection of antibodies to Peste des Petits Ruminants Virus in large ruminants. Transbound Emerg Dis. 2017;64(2):513–9.
Njeumi F, Bailey D, Soula JJ, Diop B, Tekola BG. Eradicating the Scourge of Peste Des Petits Ruminants from the World. Viruses. 2020;12(3):313.
Lembo T, Oura C, Parida S, Hoare R, Frost L, Fyumagwa R, Kivaria F, Chubwa C, Kock R, Cleaveland S, et al. Peste des petits ruminants infection among cattle and wildlife in northern Tanzania. Emerg Infect Dis. 2013;19(12):2037–40.
Zakian A, Nouri M, Kahroba H, Mohammadian B, Mokhber-Dezfouli MR. The first report of peste des petits ruminants (PPR) in camels (Camelus dromedarius) in Iran. Trop Anim Health Prod. 2016;48(6):1215–9.
Zhao H, Njeumi F, Parida S, Benfield CTO. Progress towards Eradication of Peste des Petits Ruminants through Vaccination. Viruses 2021;13(1):59.
Gibbs EP, Taylor WP, Lawman MJ, Bryant J. Classification of peste des petits ruminants virus as the fourth member of the genus Morbillivirus. Intervirology. 1979;11(5):268–74.
Balamurugan V, Sen A, Venkatesan G, Yadav V, Bhanot V, Riyesh T, Bhanuprakash V, Singh RK. Sequence and phylogenetic analyses of the structural genes of virulent isolates and vaccine strains of peste des petits ruminants virus from India. Transbound Emerg Dis. 2010;57(5):352–64.
Tounkara K, Bataille A, Adombi CM, Maikano I, Djibo G, Settypalli TBK, Loitsch A, Diallo A, Libeau G. First genetic characterization of Peste des Petits Ruminants from Niger: on the advancing front of the Asian virus lineage. Transbound Emerg Dis. 2018;65(5):1145–51.
Muniraju M, Munir M, Parthiban AR, Banyard AC, Bao J, Wang Z, Ayebazibwe C, Ayelet G, El Harrak M, Mahapatra M, et al. Molecular evolution of peste des petits ruminants virus. Emerg Infect Dis. 2014;20(12):2023–33.
Muniraju M, Munir M, Banyard AC, Ayebazibwe C, Wensman J, Zohari S, et al. Complete Genome Sequences of Lineage III Peste des Petits Ruminants Viruses from the Middle East and East Africa. Genome Announc. 2014;2(5):e01023–14.
Rume VN, Dundon WG, Belay G, Baziki JD, Diakite A, Paul A, Tessema YD, Nwankpa N, Gizaw D, Cattoli G, et al. Molecular epidemiological update of Peste des Petits Ruminants virus (PPRV) in Ethiopia. Vet Microbiol. 2019;235:229–33.
Woma TY, Adombi CM, Yu D, Qasim AM, Sabi AA, Maurice NA, Olaiya OD, Loitsch A, Bailey D, Shamaki D, et al. Co-circulation of Peste-des-Petits-Ruminants Virus Asian lineage IV with lineage II in Nigeria. Transbound Emerg Dis. 2016;63(3):235–42.
Liu F, Li J, Li L, Liu Y, Wu X, Wang Z. Peste des petits ruminants in China since its first outbreak in 2007: a 10-year review. Transbound Emerg Dis. 2018;65(3):638–48.
Wang Z, Bao J, Wu X, Liu Y, Li L, Liu C, Suo L, Xie Z, Zhao W, Zhang W, et al. Peste des petits ruminants virus in Tibet, China. Emerg Infect Dis. 2009;15(2):299–301.
Bao J, Wang Q, Li L, Liu C, Zhang Z, Li J, Wang S, Wu X, Wang Z. Evolutionary dynamics of recent peste des petits ruminants virus epidemic in China during 2013–2014. Virology. 2017;510:156–64.
Li J, Li L, Wu X, Liu F, Zou Y, Wang Q, Liu C, Bao J, Wang W, Ma W, et al. Diagnosis of Peste des Petits Ruminants in Wild and domestic animals in Xinjiang, China, 2013–2016. Transbound Emerg Dis. 2017;64(6):e43–7.
Mahapatra M, Howson E, Fowler V, Batten C, Flannery J, Selvaraj M, Parida S. Rapid Detection of Peste des Petits Ruminants Virus (PPRV) Nucleic Acid Using a Novel Low-Cost Reverse Transcription Loop-Mediated Isothermal Amplification (RT-LAMP) Assay for Future Use in Nascent PPR Eradication Programme. Viruses. 2019;11(8):699.
Polci A, Cosseddu GM, Ancora M, Pinoni C, El Harrak M, Sebhatu TT, Ghebremeskel E, Sghaier S, Lelli R, Monaco F. Development and preliminary evaluation of a New Real-Time RT-PCR assay for detection of Peste des petits Ruminants Virus Genome. Transbound Emerg Dis. 2015;62(3):332–8.
Lucas J, Holder D, Dodd K, Wei J. A versatile dual-use RT-PCR control for use in assays for the detection of peste des petits ruminants virus. J Virol Methods. 2020;277:113799.
Li L, Wu X, Liu F, Wang Z, Liu C, Wang Q, Bao J. Rapid detection of lineage IV peste des petits ruminants virus by real-time RT-PCR. J Virol Methods. 2016;235:131–3.
Yang Y, Qin X, Song Y, Zhang W, Hu G, Dou Y, Li Y, Zhang Z. Development of real-time and lateral flow strip reverse transcription recombinase polymerase amplification assays for rapid detection of peste des petits ruminants virus. Virol J. 2017;14(1):24.
Balamurugan V, Sen A, Venkatesan G, Yadav V, Bhanot V, Bhanuprakash V, Singh RK. A rapid and sensitive one step-SYBR green based semi quantitative real time RT-PCR for the detection of peste des petits ruminants virus in the clinical samples. Virol Sin. 2012;27(1):1–9.
Flannery J, Rajko-Nenow P, Arnold H, van Weezep E, van Rijn PA, Ngeleja C, Batten C. Improved PCR diagnostics using up-to-date in silico validation: an F-gene RT-qPCR assay for the detection of all four lineages of peste des petits ruminants virus. J Virol Methods. 2019;274:113735.
Onozuka N, Ohki T, Oka N, Maoka T. One-step real-time multiplex reverse transcription-polymerase chain reaction assay with melt curve analysis for detection of potato leafroll virus, potato virus S, potato virus X, and potato virus Y. Virol J. 2021;18(1):131.
Chen KY, Xu JX, Wang MM, Hu D, Xie F, Huang D, Chen J, Yang T, Zhang J, Song F, et al. Single probe PCR melting curve analysis MTHFR C677T SNP sites. Anal Biochem. 2021;619:114102.
Liu Z, Bingga G, Zhang C, Shao J, Shen H, Sun J, Zhang J. Application of duplex fluorescence melting curve analysis (FMCA) to identify Canine Parvovirus Type 2 variants. Front Microbiol. 2019;10:419.
Meng C, Huang Y, Rehman ZU, Hu W, Li C, Liang R, Chen Z, Song K, Wei T, Liu G. Development of an MCA-Based real Time RT-qPCR assay for the simultaneous detection and differentiation of duck Hepatitis A Virus types 1 and 3. Virol Sin. 2020;35(5):666–9.
Acknowledgements
We thank International Science Editing for its linguistic assistance during the preparation of this manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (No. 32172832, No. 32000109), Shanghai Sailing Program (20YF1457700), the China Postdoctoral Science Foundation (No. 2019M660885, No. 2021T140718), the Central Public-interest Scientific Institution Basal Research Fund (2021JB08).
Author information
Authors and Affiliations
Contributions
GQL, CCM, and JYT conceived this project. JYT and HYD performed the experiments. HYD, NNJ, AXT, and JZ analyzed the data. JYT and CCM wrote the original draft. CFL, CCM, and GQL reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Tang, J., Du, H., Tang, A. et al. Simultaneous detection and identification of Peste des petits ruminants Virus Lineages II and IV by MCA-Based real-time quantitative RT-PCR assay within single reaction. BMC Vet Res 19, 11 (2023). https://doi.org/10.1186/s12917-023-03568-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12917-023-03568-6