
Abstract
Interleukin-2 (IL-2) and its receptor (IL-2R) are essential in orchestrating immune responses. Their function and expression in the tumor microenvironment make them attractive targets for immunotherapy, leading to the development of IL-2/IL-2R-targeted therapeutic strategies. However, the dynamic interplay between IL-2/IL-2R and various immune cells and their dual roles in promoting immune activation and tolerance presents a complex landscape for clinical exploitation. This review discusses the pivotal roles of IL-2 and IL-2R in tumorigenesis, shedding light on their potential as diagnostic and prognostic markers and their therapeutic manipulation in cancer. It underlines the necessity to balance the anti-tumor activity with regulatory T-cell expansion and evaluates strategies such as dose optimization and selective targeting for enhanced therapeutic effectiveness. The article explores recent advancements in the field, including developing genetically engineered IL-2 variants, combining IL-2/IL-2R-targeted therapies with other cancer treatments, and the potential benefits of a multidimensional approach integrating molecular profiling, immunological analyses, and clinical data. The review concludes that a deeper understanding of IL-2/IL-2R interactions within the tumor microenvironment is crucial for realizing the full potential of IL-2-based therapies, heralding the promise of improved outcomes for cancer patients.
Similar content being viewed by others

Introduction
Interleukin-2 (IL-2) is a cytokine produced by certain immune cells, such as T and B cells, which plays a crucial role in regulating the immune system by stimulating the proliferation and activation of these cells [1]. The Interleukin-2 receptor (IL-2R), a transmembrane glycoprotein receptor, is prominently located on the surface of T and B cells, among other immune system cells. This receptor has a crucial role in the immune response, as it binds to IL-2, instigating a cascade of events that culminate in the activation and proliferation of these cells. This process underscores the fundamental role of IL-2R in the modulation and functioning of the immune system [2]. The cooperative action of IL-2 and IL-2R is necessary for the proper functioning of the immune system, as it helps to maintain the balance between the activation and suppression of the immune response to pathogens [3].
Despite the beneficial role of IL-2 in instigating immune responses to attack tumor cells, its ability to expand regulatory T cells (Tregs), potentially dampening anti-tumor immunity, presents a nuanced and paradoxical situation. The intriguing complexity of the IL-2 and IL-2R signaling pathways extends beyond routine immune regulation, with compelling evidence illuminating their roles within the tumor microenvironment (TME). In addition, IL-2 and IL-2R have also been found to play a significant role in the development and progression of cancer [4]. Moreover, IL-2 has been shown to promote the growth and survival of specific tumor cells, and studies have demonstrated that IL-2R is involved in the angiogenesis or growth of new blood vessels in tumors [5,6,7,8]. Furthermore, IL-2 and IL-2R have been implicated in the metastatic process, whereby cancer cells migrate from the primary tumor to distant body parts [8, 9].
Consequently, therapies targeting IL-2 and IL-2R are currently under development and rigorous testing as potential oncological treatments, demonstrating encouraging outcomes in clinical trials [5, 10, 11]. These innovative therapeutic strategies aim to invigorate the body’s immune system, enabling it to identify and subsequently eradicate cancer cells. The pivotal role of IL-2 and IL-2R in this immunological response underscores their significance in the advancement of cancer treatment modalities. IL-2 has been shown to activate T cells, which can then target and kill cancer cells, while IL-2R is expressed on the surface of certain immune cells, such as natural killer (NK) cells, and can help to stimulate these cells to fight cancer [12, 13].
Although IL-2 has been utilized in immunotherapy, the success of these strategies is variably influenced by the differential expression of IL-2 and IL-2R across diverse cancer types. For instance, IL-2-based immunotherapy has demonstrated increased efficacy in malignancies such as melanoma and renal cell carcinoma (RCC), attributed to their higher expression of IL-2R ([7, 8]). In addition, IL-2 and IL-2R are involved in several cancer-related pathways. For example, IL-2 has been shown to regulate cell proliferation, apoptosis, and angiogenesis, while IL-2R has been implicated in tumor growth, metastasis, and immune evasion [5, 14,15,16]. In targeting IL-2 and IL-2R-mediated pathways, there is a promising approach for treating cancer, and various IL-2 and IL-2R-targeted therapies are currently being developed and tested in clinical trials [12, 17, 18].
However, there are still critical challenges in the therapeutic targeting of IL-2 and IL-2R in cancer, such as the lack of specificity of these molecules and the potential for side effects. For instance, IL-2 is known to regulate several immune pathways, and targeting it could lead to an over-activation of the immune system, resulting in adverse effects [19,20,21,22]. Therefore, this review article aims to delineate the dichotomy of IL-2 and IL-2R’s functions within the tumor milieu and its implications for cancer immunotherapy.
Our understanding of the IL-2 and IL-2R pathways within the context of cancer has significantly evolved, opening avenues for improved and safer therapeutic interventions. Nevertheless, there is still much to discern. This review seeks to address the key questions: How can we balance the dual roles of IL-2 to optimize its anti-tumor effects? Furthermore, how do the IL-2 and IL-2R expression variations among different cancer types influence the therapeutic response? Moreover, lastly, what is the potential of IL-2 and IL-2R as diagnostic, prognostic, or predictive markers for cancer? By addressing these questions, we aim to provide a comprehensive overview of IL-2 and IL-2R’s functions in the TME, catalyzing further research toward developing more effective cancer immunotherapies.
Deciphering IL-2 and IL-2R: biology, signaling, and regulation
Mechanisms of IL-2 and IL-2R signaling
IL-2 is a critical cytokine essential in the adaptive immune system’s regulation [23]. Predominantly produced by T helper cells and NK cells upon antigen recognition, IL-2 originates from other immune cells, such as dendritic cells [23, 24]. Its primary function is to stimulate the proliferation and differentiation of T and B lymphocytes [23, 24]. IL-2R is constituted by three unique subunits: CD25, also denoted as the alpha chain or IL-2Rα; CD122, identified as the beta chain or IL-2Rβ; and CD132, referred to as the gamma chain or IL-2Rγ [24, 25]. These chains contribute uniquely to the IL-2R’s functionality. IL-2 binding initiates via CD25, enhancing the receptor’s affinity for the cytokine [24, 25]. CD122, predominantly expressed in NK cells, is also found in other immune cells, playing a crucial role in cytokine binding and signal transduction1. CD132 assists in assembling the receptor complex and participates in signaling, contributing to the overall immune response [24, 25].
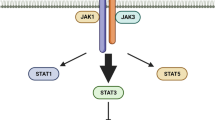
The IL-2R/JAK/STAT signaling pathway is instrumental in T and B lymphocyte development and functional maturation. Upon activation, this pathway orchestrates the nuclear translocation of specific transcription factors, thereby regulating the expression of multiple target genes, which notably include IL-2 itself. This results in a self-perpetuating positive feedback loop [23, 26]. In addition to the JAK/STAT pathway, IL-2R activation also instigates two other critical signaling cascades: the Mitogen-Activated Protein Kinase (MAPK) pathway [26, 27], and the Phosphatidylinositol 3-Kinase (PI3K) pathway [28]. While the MAPK pathway is crucial for regulating cell proliferation and differentiation processes, the PI3K pathway is instrumental in ensuring cell survival and promoting lymphocyte proliferation and differentiation [28]. (Figure.1).

This diagram provides a comprehensive illustration of the interleukin-2 (IL-2) and interleukin-2 receptor (IL-2R) pathway within the tumor microenvironment, detailing its intrinsic role in tumor immunology. IL-2R, encompassing three distinct chains—IL-2Rα (CD25), IL-2Rβ (CD122), and γc (CD132)—comes in three unique forms formed by different combinations of these chains. The depicted schematic demonstrates how the IL-2/IL-2R pathway activates Janus kinase 1 (JAK1) and Janus kinase 3 (JAK3) following their phosphorylation, thereby prompting the downstream signaling across multiple specialized pathways. Each of these pathways displays unique modulation within regulatory T cells (Treg cells), differing distinctly from effector T cells (Teff) or conventional T cells (Tconv). Notable components that demonstrate increased activity within Treg cells, in comparison to Teff or Tconv cells, include the signal transducer and activator of transcription 5 (STAT5) and phosphatase and tensin homolog (PTEN). On the other hand, the phosphatidylinositol 3-kinase (PI3K)/Akt and mitogen-activated protein kinase (MAPK)/extracellular signal–related kinase (Erk) pathways show enhanced activity within Teff or Tconv cells. Additionally, other significant transcription factors and regulators like IL2-inducible T-cell kinase (ITK), inhibitors of nuclear factor-κB (IκB), Nuclear factor of activated T-cells (NFAT), Activator protein-1 (AP-1), the C-JUN protein—a signal-transducing transcription factor of the AP-1 family, and Protein c-Fos—a proto-oncogene that is the human homolog of the retroviral oncogene v-fos, are also engaged in the IL-2 and IL-2R signaling pathway. These components further modulate the intricate signaling dynamics within the Treg, Teff, and Tconv cells. The Suppressors of Cytokine Signaling1 (SOCS1), a critical regulator of cytokine signaling, is also implicated in this pathway and contributes to the complex interplay of signals within the tumor microenvironment. This detailed insight into the IL-2/IL-2R signaling pathway, along with the roles of the associated proteins, aims to facilitate an enhanced understanding of tumor immunology and could possibly pave the way for novel therapeutic strategies. The figure was created using the online tool https://biorender.com/
IL-2, a potent lymphocyte growth factor, utilizes these signaling pathways to govern various aspects of the adaptive immune response. For instance, it fosters the proliferation and differentiation of CD4 + T cells into Th1 and Th2 cells [29, 30], while it aids CD8 + T cells in their development into cytotoxic T lymphocytes (CTLs) [31]. Additionally, IL-2 influences the B cell lineage by promoting differentiation into plasma cells and contributes to the development of Tregs, a subset critical for maintaining peripheral tolerance [32].
Besides T and B cells, IL-2 also profoundly impacts NK cells. By triggering the JAK/STAT pathway, IL-2 instigates the production of Interferon-gamma (IFN-γ), a cytokine essential for the activation and proliferation of NK cells [33]. Furthermore, IL-2 aids in the differentiation of NK cells into effector and memory phenotypes, further expanding its influence over the immune response [34] (Fig. 2).

The schematic diagram illustrates the multifaceted role of Interleukin-2 (IL-2) in the homeostasis and functional dynamics of Natural Killer (NK) cells, which are characterized as CD56+CD3− large granular lymphocytes integral to the human innate immune response. IL-2 is instrumental in the differentiation process, facilitating the transformation of NK precursors into mature NK cells. It also maintains the homeostasis of mature NK cells by regulating their survival, proliferation, and apoptosis. The diagram further highlights the influence of IL-2 on the functional aspects of mature NK cells. It enhances the cytolytic activity of both CD56bright and CD56dim NK cells and significantly amplifies cytokine secretion, particularly within the CD56bright human NK subset. In secondary lymphoid tissues such as lymph nodes, IL-2 promotes the acquisition of Killer Cell Immunoglobulin-like Receptors (KIR; also known as CD158) and CD16 expression by CD56bright NK cells. The diagram also showcases the characteristics of cytokine-induced memory-like (CIML) NK cells, which include increased expression of CD25 (IL-2Ra), decreased expression of KIRs, and Transforming Growth Factor-beta (TGFβ) receptors. These changes potentially alleviate inhibitory mechanisms in CIML NK cells. The enhanced production of Interferon-gamma (IFNγ) may augment anti-tumor responses. The schematic also highlights the metabolic changes in CIML NK cells. Metabolic alterations, including the roles of glucose transporter and transferrin receptor, are emphasized for their critical contribution to the long-term persistence and recall functions of CIML NK cells. Abbreviations: fms-like tyrosine kinase 3 ligand (FL), kit ligand (KL).The figure was created using the online tool https://biorender.com/
Regulation of IL-2 and IL-2R by transcription factors and regulators
The regulation of IL-2 and IL-2R, crucial in governing the immune response, is modulated by several transcription factors [35,36,37]. Positive regulators such as the Nuclear Factor of Activated T-cells (NFAT) and Activator Protein-1 (AP-1) bind to the promoter and enhancer regions of IL-2, respectively [35, 36]. JAKs, STAT5, and STAT3 also augment IL-2 receptor signaling [37]. Conversely, negative regulators include Suppressor of Cytokine Signaling-1 (SOCS1), which inhibits JAK/STAT signaling [38], and Forkhead box protein 3 (FoxP3), a transcription factorsuppressing IL-2 production and IL-2 receptor expression in regulatory T cells [39, 40] (Fig. 1; Table 1)([41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65]). Balancing these positive and negative regulators of IL-2 and its receptor is crucial for appropriate immune function. Dysregulation can lead to autoimmune disorders and cancer, emphasizing the importance of understanding the biology, signaling pathways, transcription factors, and regulators of IL-2 and IL-2R [35,36,37,38,39,40].
IL-2/IL-2R in the TME
TME is a complex, dynamic ecosystem comprising tumor cells, immune cells, stromal cells, and extracellular matrix components that significantly influence tumor growth, invasion, and metastasis [17]. IL-2 and IL-2R have garnered attention within this milieu due to their intricate role in modulating immune responses to tumors [17]. Evidence suggests that dysregulated IL-2/IL-2R signaling within the TME can profoundly impact tumor growth and anti-tumor immune responses. For instance, IL-2/IL-2R signaling may promote the expansion of Tregs, which could potentially inhibit anti-tumor immune responses, thereby facilitating tumor growth [17]. Conversely, other research indicates that IL-2/IL-2R signaling may promote CTL and NK cell infiltration into the TME, enhancing anti-tumor immune responses and impeding tumor growth [66]. It becomes apparent that the role of IL-2/IL-2R signaling in the TME is multifaceted and complex, and a complete understanding of these mechanisms remains a research priority. Promising therapeutic strategies are emerging from these insights. Recent preclinical models demonstrate that IL-2/IL-2R agonists could enhance anti-tumor immune responses and reduce tumor growth [18]. Moreover, combined targeting of IL-2/IL-2R signaling with other immunotherapies may exhibit synergistic effects on tumor growth inhibition [17, 67], (Fig. 3; Table 2) [68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96].

Interplay of IL-2/IL-2R Signaling in the Tumor Microenvironment (TME): This figure elucidates the multifaceted roles of Interleukin-2 (IL-2) and the Interleukin-2 Receptor (IL-2R) within the TME, illustrating the varying influence of IL-2 based on its concentration. Lower levels of IL-2 can foster a regulatory microenvironment which potentially augments tumor growth, while higher levels act as stimulants for immune cells, fostering tumor elimination. Additionally, the figure underscores the heterogeneous impacts of dysregulated IL-2/IL-2R signaling. On one hand, it can stimulate regulatory T cells (Tregs), which may suppress anti-tumor responses, on the other, it can induce infiltration of cytotoxic T cells (CTLs) and NK cells, thus fortifying anti-tumor immune responses. It further emphasizes how IL-2 connects the adaptive and innate immune systems, highlighting the role of cells such as T cells, B cells, NK cells, and dendritic cells, both as IL-2 producers and responders. The varying sizes of arrows in the figure represent the differential intensity of IL-2 production and its effects on different cell types within the TME. Despite its intricate nature, modulating IL-2/IL-2R signaling is underscored as a promising therapeutic avenue, particularly when synergized with other immunotherapies. The figure was created using the online tool https://biorender.com/
IL-2/IL-2R: T-cell regulation within the TME
Within the TME, T-cell function is a critical determinant of tumor growth and progression [97, 98]. Dysregulation of T-cell function, including impaired T-cell receptor signaling, reduced expression of co-stimulatory molecules, and upregulated expression of inhibitory receptors (such as PD-1, CTLA-4, and TIM-3), is a common immune evasion tactic employed by tumors [99,100,101,102,103,104]. Furthermore, tumors can induce the accumulation and differentiation of Tregs in the TME through the secretion of various cytokines, chemokines, and growth factors [100].
IL-2/IL-2R signaling, an essential player in T-cell regulation, could hold therapeutic potential in rebalancing the TME. Activated T-cells produce IL-2, a cytokine crucial for the proliferation, survival, and function of effector T-cells [29, 105]. IL-2R, expressed on various immune cells, mediates IL-2 signaling and is pivotal in determining the balance between effector T-cells and Tregs within the TME [29, 105]. Specifically, IL-2 signaling can bolster the proliferation and function of effector T-cells while restraining Treg proliferation and function [106]. In addition, IL-2 signaling also promotes the reprogramming of Tregs into effector T-cells, shifting the TME from a pro-tumor to an anti-tumor state [107, 108] (Fig. 3).
Evidence links dysregulated IL-2/IL-2R signaling to poor prognosis in various types of cancer, including melanoma, RCC, and breast cancer [109,110,111,112]. Emerging IL-2-based therapies show promise in enhancing the anti-tumor immune response and improving the efficacy of immunotherapy in cancer patients [113].
IL-2/IL-2R regulatory roles on NK cells, macrophages, neutrophils, and B cells in the TME
The TME is a dynamic network that integrates various cells, including immune and cancer cells, which are instrumental in cancer progression and metastasis [114]. IL-2/IL-2R signaling plays a pivotal role in modulating immune cells, and its dysregulation can contribute to tumor evasion [115]. The following sections delve into the function of this pathway on specific immune cell types in the TME.
Regulation of NK cells by IL-2/IL-2R
NK cells possess an innate capability to recognize and eliminate malignant cells, and their function is critically modulated by IL-2 [13] (Fig. 2). Zhang et al.‘s study illustrated that IL-2 signaling amplifies the cytotoxic potential of NK cells, thereby inhibiting tumor growth and metastasis. Notably, NK cells recognize and eliminate tumor cells in an MHC-independent manner, distinguishing them from T-cell mediated tumor recognition [116]. Furthermore, this research found that IL-2 treatment in a mouse model of breast cancer reduced tumor growth and metastasis, underscoring the potential for NK cell-based immunotherapies [33, 116].
IL-2/IL-2R influence on macrophages and neutrophils
Similarly, IL-2 impacts macrophages’ functionality, promoting their differentiation into the M1 phenotype, known for their anti-tumor and potent tumoricidal activities [117,118,119,120,121,122,123,124]. Furthermore, IL-2/IL-2R signaling plays a critical role in neutrophil activity within the TME, modulating their recruitment, activation, and lifespan, thereby influencing tumor progression [118]. A deeper understanding of these mechanisms can pave the way for novel strategies targeting IL-2/IL-2R signaling to enhance neutrophils’ anti-tumor activities or minimize their pro-tumor activities [119,120,121,122,123,124,125,126,127].
The role of IL-2/IL-2R signaling in B cells
In comparison to other immune cells, B cells are often overlooked in the TME context. However, recent research has demonstrated the importance of IL-2/IL-2R signaling in regulating B cells within the TME [128]. IL-2 can directly stimulate B cells, enhancing their proliferation, differentiation into antibody-secreting plasma cells, and co-stimulatory molecule expression, such as CD80 and CD86 [128].
However, IL-2/IL-2R signaling can also promote Treg expansion, potentially inhibiting the anti-tumor immune response [129]. The interplay between Tregs, B-cells, and other cytokines and immune cells within the TME adds to the complexity of the immune response [130,131,132,133,134,135,136,137]. The role of IL-2/IL-2R signaling in B-cell responses within the TME requires further exploration to fully understand these mechanisms and develop targeted immunotherapeutic strategies [138].
Interactions and roles of IL-2/IL-2R in dendritic cells (DCs) and myeloid-derived suppressor cells (MDSCs) within the TME
Critical to regulating anti-tumor immune responses within the TME are immune cells, such as DCs and MDSCs, which are influenced by IL-2 /IL-2R signaling [139,140,141,142]. DCs, as professional antigen-presenting cells, initiate and regulate immune responses. IL-2 is pivotal in augmenting their maturation and activation, enhancing antigen presentation capacity, and increasing T-cell activation [139] By upregulating co-stimulatory molecules, such as CD80 and CD86, IL-2 facilitates the activation of naïve T cells [140]. Additionally, IL-2 encourages DC migration to lymph nodes, fostering T cell activation and instigating anti-tumor immune responses [143].
Contrastingly, MDSCs, a diverse population of immature myeloid cells, accumulate within the TME and suppress the anti-tumor immune response. IL-2 exerts a dual role on MDSCs; it stimulates the expansion and accumulation of MDSCs, promoting an immunosuppressive microenvironment [141] while concurrently inducing MDSC differentiation into mature myeloid cells. This differentiation reduces suppressive functionality and potentially fosters anti-tumor immune responses [142]. The IL-2/IL-2R signaling pathway is central to the interplay between DCs and MDSCs within the TME. IL-2, by enhancing the maturation and activation of DCs, promotes antigen presentation and T-cell activation. Consequently, these activated T cells produce IL-2, which induces MDSC differentiation into mature myeloid cells, decreasing immunosuppression within the TME [29]. Nevertheless, IL-2 can simultaneously drive MDSC expansion and accumulation within the TME, creating an immunosuppressive environment [144]. In addition, MDSCs express high levels of IL-2Rα or CD25 [145], implying a possible role for IL-2 signaling in their functionality. Indeed, studies suggest that IL-2 promotes MDSC expansion and activation [29], while blockade of the IL-2/IL-2R pathway may reduce their suppressive function within the TME [144]. Recent studies highlight the intricate relationship between DCs and MDSCs in the TME, regulated by the IL-2/IL-2R pathway. Specifically, IL-2 produced by activated T cells can stimulate MDSC proliferation and differentiation, which may inhibit the maturation and antigen-presenting function of DCs [146, 147]. This reciprocal regulation is thought to contribute to the TME’s immunosuppressive nature, where MDSCs hinder the activation and proliferation of effector T cells. At the same time, DCs struggle to initiate a robust immune response against cancer cells [148].
Contemporary research has probed the role of IL-2 signaling in MDSC differentiation and functionality [149]. It was found that IL-2 signaling is critical for MDSC differentiation and promotion of their immunosuppressive function. This signaling also activates the STAT5 pathway, driving the upregulation of crucial genes involved in MDSC differentiation and functionality [150]. Moreover, IL-2 signaling blockade enhanced the anti-tumor immune response and inhibited tumor growth in murine models [74]. Another study investigated the DC-MDSC interplay within the TME, focusing on the IL-2/IL-2R pathway [151]. They discovered that MDSCs could directly inhibit the function of DCs via the IL-2/IL-2R pathway. Intriguingly, MDSCs express high levels of IL-2R, which competes with DCs for IL-2 binding. This competitive interaction reduces IL-2 signaling in DCs, impairing their function and inhibiting the anti-tumor immune response. However, blocking IL-2R signaling in MDSCs was observed to reverse this effect and enhance DC function [151].
IL-2 and IL-2R in Cancer Therapy: a balance between immunostimulation and immunosuppression
IL-2 and IL-2R play pivotal roles in the immunological response against cancer, wielding influence over immune regulation and, thus, cancer therapy. Their function extends to the activation of immune cells, such as T cells, B cells, and NK cells, augmenting immune responses against cancer cells. However, their role is dualistic, wielding effects of both immunostimulation and immunosuppression. This duality can present both benefits and challenges in cancer therapy [152,153,154].
Immunostimulatory effects of IL-2/IL-2R
IL-2 is instrumental in stimulating the proliferation and activation of T cells and NK cells. This activity enhances the anti-tumor immune response, mainly by IL-2 binding to the high-affinity IL-2R (IL-2Rα/β/γ) expressed on activated T cells [152]. This interaction results in the expansion and activation of effector T cells, responsible for the direct or indirect killing of cancer cells, primarily through cytokine production such as Interferon gamma (IFN-γ) and tumor necrosis factor alpha (TNF-α) [153]. In addition, IL-2 can further induce the expansion of NK cells, which increases tumor-specific cytotoxicity [154]. The efficacy of IL-2 as a cancer immunotherapy has been evaluated in various clinical trials. High-dose IL-2 demonstrates potential, inducing objective responses in up to 15% of patients with metastatic melanoma and RCC, with some patients achieving durable complete responses [7, 10, 91, 155, 156].
Immunosuppressive role of IL-2/IL-2R
IL-2/IL-2R signaling plays a pivotal role in the activation and proliferation of TILs, immune cells that infiltrate tumors and mediate anti-tumor immune responses. However, the influence of IL-2 on TILs is multifaceted and largely determined by the TME, the presence of Tregs, and the activation state of TILs. These elements can promote the expansion of Tregs and MDSCs that suppress effector T cell activity, thereby paradoxically fostering tumor growth and creating an immunosuppressive effect. Despite the potential of IL-2, the use of high-dose IL-2 is limited due to severe associated toxicities, such as hypotension, pulmonary edema, and renal failure, which restrict its broader clinical use [157,158,159,160,161,162].
Strategies for IL-2/IL-2R therapeutic targeting
The intricacies of IL-2 and IL-2R signaling necessitate a delicate balancing act in their therapeutic targeting. Key to this balance is the differential expression of IL-2R subunits on various immune cells [5]. The ratio of IL-2R subunit expression on different cell types can influence the overall effects of IL-2 signaling within the immune system. For instance, targeting IL-2R selectively on effector T cells could enhance the anti-tumor immune response [163]. Conversely, targeting IL-2R on Tregs could lead to an undesirable promotion of immunosuppression and tumor growth [164]. Therefore, IL-2 has been used in combination with other immunotherapies, such as checkpoint inhibitors, to optimize anti-tumor immune responses. For example, high-dose IL-2, combined with ipilimumab (an anti-CTLA-4 antibody), has shown promise by inducing durable responses in patients with metastatic melanoma [165]. However, combination immunotherapy could lead to an escalation in toxicity and the induction of autoimmune reactions.
Consequently, the most practical combination of immunotherapies requires meticulous evaluation in clinical trials. Another promising avenue is using IL-2 as an adjuvant in cancer vaccines to bolster the activation and proliferation of tumor-specific T cells [166,167,168]. The GVAX vaccine, which consists of irradiated tumor cells genetically modified to secrete GM-CSF, is one such example that has been paired with IL-2 to enhance the anti-tumor immune response [169,170,171]. Although preclinical models have shown promise, the efficacy of IL-2-based cancer vaccines remains to be verified in clinical trials.
Challenges in therapeutic targeting of IL-2 and IL-2R in cancer
Despite their immense potential, therapeutic targeting of IL-2 and IL-2R in cancer poses several challenges. A significant challenge is the complexity of IL-2 signaling pathways. IL-2 can activate multiple signaling pathways, including the JAK-STAT, PI3K-AKT, and MAPK pathways, exerting pro- and anti-tumor effects [172, 173]. In addition, the heterogeneity of IL-2R expression on different immune cell populations and the severe side effects associated with IL-2 treatment, such as life-threatening cytokine release syndrome, further complicates the development of effective and safe treatments [174]. Moreover, the issues of delivery and dosing present additional challenges for IL-2-based immunotherapies. As a large protein, IL-2 requires intravenous administration and exhibits a short half-life in vivo, making it challenging to achieve sustained therapeutic levels [13]. In addition, high-dose IL-2, although linked to higher response rates in metastatic melanoma, is associated with increased toxicity [175]. Furthermore, the efficacy of IL-2 and IL-2R-targeted therapies is limited in particular cancer types, such as breast or lung cancer [176].
Soluble IL-2R (sIL-2R) and IL-2Rα: emerging biomarkers and therapeutic targets in cancer
The IL-2R complex, composed of IL-2Rα, IL-2Rβ, and IL-2Rγ, plays critical roles in the immune system [177, 178]. Each subunit has specific locations and functions within the immune system (Fig. 1; Table 3) [179,180,181,182,183,184,185,186,187,188,189,190,191,192,193,194,195,196,197,198]. Importantly, IL-2Rα, primarily expressed in activated T cells, regulatory T cells, and activated B cells, initiates downstream signaling cascades upon binding to IL-2 [23, 199]. The soluble form of the receptor, sIL-2R, produced by proteolytic cleavage of the membrane-bound IL-2R complex, has been identified in the serum of patients with Hodgkin’s lymphoma and some non-lymphoid cancers [182, 183, 185]. In contrast to its membrane-bound counterpart IL-2Rα, this form presents a longer half-life and different modes of interaction with IL-2 [186, 187]. Both sIL-2R and IL-2Rα have been implicated in numerous immune-mediated diseases, signifying their potential as markers of disease activity and progression [186, 188, 200].
sIL-2R and IL-2Rα as cancer biomarkers
sIL-2R and IL-2Rα have emerged as promising biomarkers for cancer diagnosis and prognosis. They are found on the surface of cells stimulated by IL-2, with IL-2Rα being a low-affinity receptor whose expression is upregulated in malignant cells [181, 201,202,203]. Increased levels of sIL-2R and IL-2Rα are observed in several cancers, including colorectal, prostate, breast, and lung cancer [204,205,206,207], often indicating a poor prognosis. Moreover, they serve as potential diagnostic markers as elevated levels of sIL-2R and IL-2Rα have been detected in patients with colorectal cancer and those at increased risk of prostate cancer [189, 208]. Further, these markers have potential utility in monitoring treatment efficacy. For example, decreased levels of sIL-2R and IL-2Rα in response to chemotherapy and radiation therapy signify a favorable prognosis in patients with prostate and colorectal cancers [204, 209,210,211].
Therapeutic potential of sIL-2R and IL-2Rα
Beyond their roles as biomarkers, sIL-2R and IL-2Rα are under investigation as therapeutic targets for cancer [184, 186, 209, 212, 213]. The ability of sIL-2R to modulate the immune system and tumor microenvironment underlines its potential as cancer therapeutics [10, 188]. It has been proposed that sIL-2R may induce tumor cell apoptosis, inhibit tumor angiogenesis, and interfere with oncogenic pathways [214]. Similar anticancer effects have been attributed to IL-2Rα, including the induction of apoptosis in multiple myeloma cells and the inhibition of oncogenic pathways, such as the JAK-STAT pathway [215,216,217,218].
Preliminary clinical studies underscore the therapeutic potential of sIL-2R and IL-2Rα. A phase I trial demonstrated the safety and tolerability of sIL-2R in patients with advanced solid tumors and lymphomas [219,220,221]. A subsequent phase I/II trial reported a significant reduction in tumor burden in patients with advanced metastatic RCC treated with sIL-2R [222]. Moreover, a phase II trial revealed a significant survival benefit in patients with advanced non-small cell lung cancer (NSCLC) following IL-2Rα treatment [223].
IL-2 and engineered IL-2 for immunotherapy of autoimmunity and cancer
IL-2 and engineered IL-2 for immunotherapy of autoimmune disorders
IL-2, a critical cytokine, promotes the growth and activation of T cells, especially Tregs [153]. Tregs are indispensable in controlling immune responses and preventing autoimmune diseases [153]. However, systemic administration of IL-2 is associated with adverse effects due to its pleiotropic effects on various immune cells [71]. A novel strategy developed to mitigate this problem involves receptor-gated IL-2 delivery through an anti-human IL-2 antibody [71]. The receptor-gated IL-2 delivery is a targeted approach where the engineered anti-human IL-2 antibody binds to IL-2 receptors on Tregs, effectively delivering IL-2 specifically to these cells [71]. This enhances Treg activation and their suppressive activity, selectively impacting these cells without disturbing other immune cells [71]. This approach has demonstrated potential in activating Tregs across various species, such as mice, monkeys, and humans [224]. Studies using mouse models of autoimmune diseases like type 1 diabetes, multiple sclerosis, and graft-versus-host disease have illustrated decreased disease severity and improved survival with this approach [71]. Recent studies have also shown that IL-2 therapy can be effective in treating systemic lupus erythematosus by expanding Tregs and reducing disease activity [225].
Recognizing the therapeutic potential of IL-2, researchers are focused on engineering IL-2 for improved efficacy and safety in treating cancer and autoimmune diseases. Modifying IL-2 structure and function can increase its therapeutic potential [226]. Several strategies include designing IL-2 variants to selectively target Tregs [227], modifying the IL-2 receptor to increase Treg selectivity, or creating IL-2 variants with reduced binding affinity to non-specific immune cells [109, 228]. Among the engineered IL-2 variants, “aldesleukin” or “recombinant human IL-2” (rIL-2) has been extensively researched. It has shown promise in enhancing Treg activity and suppressing autoimmune responses, although associated with potential toxicities like vascular leak syndrome (VLS), limiting its clinical application [109, 226, 229, 230].
IL-2 and engineered IL-2 for cancer immunotherapy
The therapeutic potential of IL-2 has also been recognized in the field of cancer immunotherapy. IL-2 can enhance the anti-tumor immune response and reduce tumor growth, as demonstrated in mouse cancer models [231]. However, the systemic administration of IL-2 is associated with adverse effects, limiting its clinical application [71]. To overcome these limitations, researchers are engineering IL-2 for improved efficacy and safety in cancer treatment.
Further advancements in IL-2 engineering introduced IL-2 “superkines“(discussed beloe), such as “NARA1”. These molecules exhibit enhanced IL-2 receptor binding and improved signaling properties, providing increased potency and selectivity for Tregs compared to rIL-2 [179, 232,233,234]. However, these superkines may also have the potential to induce autoimmune responses due to their increased activity [234].
Several clinical trials have evaluated the safety and efficacy of these engineered IL-2 molecules for cancer and autoimmune disorders. For example, despite its limitations, Aldesleukin has been FDA-approved for treating metastatic melanoma and RCC [153, 235, 236]. Other engineered IL-2 molecules, like NKTR-214 and AMG 592, have demonstrated selectivity for activating Tregs and NK cells, showing potential in enhancing immune responses against tumors, and are under clinical evaluation [68, 237, 238]. ALT-803, another engineered IL-2 variant, is under clinical trial for treating various cancers, showing promise in preclinical models of multiple myeloma [ClinicalTrials.gov Identifier: NCT02099539]. Moreover, the IL-2 variant ‘tebentafusp’ has shown promise in clinical trials for treating uveal melanoma [239]. It selectively binds to T cells that recognize a specific tumor antigen, leading to their activation and expansion. However, due to its activity on non-tumor-specific T cells, it may also induce autoimmune responses.
Recent studies have also shown that engineered IL-2 molecules can enhance the efficacy of immune checkpoint inhibitors in cancer treatment by promoting the activation and expansion of tumor-specific T cells [240].
While these engineered IL-2 molecules show promise, they also present significant challenges. Potential toxicities, including VLS, can limit their clinical use [109, 239, 241,242,243]. Other limitations include the potential for inducing autoimmune responses [243], their high cost, and complex manufacturing processes [243, 244].
The burgeoning field of engineered IL-2 for cancer immunotherapy, though in its infancy, carries immense potential, yet it is not devoid of substantial hurdles [243]. A key obstacle lies in the creation of IL-2 variants that can selectively stimulate Tregs without triggering other immune cells [225, 240, 245]. Furthermore, fine-tuning the dosage and administration of engineered IL-2 to strike a balance between therapeutic effectiveness and toxicity presents another significant challenge [243, 246, 247].
Current efforts are directed towards designing IL-2 therapeutics with enhanced in vivo half-lives, targeting specific IL-2 receptor conformations to stimulate specific T cell subsets, or delivering localized therapies to target tissues [241]. A recent trend in the engineering of IL-2 for the therapy of cancer and autoimmunity is the development of PD-1-IL-2R agonists. Deak and colleagues (2022) effectively highlighted the potential of PD-1-IL-2R agonists in eliciting robust and selective immune responses by promoting the expansion of effector T cells without the simultaneous expansion of Tregs. They also underscored the synergistic benefits of PD-1-IL-2R agonists with conventional anti-PD-1 therapy, representing a novel, promising avenue for cancer immunotherapy [248]. Moreover, the study by Zhang et al. (2021) heralded the creation of orthogonal IL-2 systems, serving as a powerful platform for delineating the complex network of cytokine-receptor interactions and their role in immune modulation. By developing murine and human IL-2 orthogonal systems, they demonstrated a controlled activation of IL-2-dependent signaling pathways, providing insights into potential therapeutic strategies for both autoimmune diseases and cancer [249]. Furthermore, Evans et al.‘s work in 1999 on IL-2R-based chimeric molecules paved the way for a better understanding of how IL-2 and its receptor interact, contributing significantly to the development of more effective therapies [250]. These chimeric molecules can harness the power of the immune system more selectively and potently, addressing issues such as toxicity and lack of specificity seen with the conventional IL-2 therapy.
The primary goal of these advancements is to augment therapeutic efficacy while minimizing associated toxicity. Despite these hurdles, engineered IL-2 therapies are poised to become a significant addition to our current arsenal of cancer treatments [241, 251], provided the obstacles of optimizing effectiveness and minimizing potential drawbacks are carefully navigated.
Future outlooks and perspectives on IL-2-based therapies for immunotherapy: balancing promise and challenges
IL-2, a cytokine critical in T-cell activation and proliferation, serves as a vital component in cancer immunotherapy [252]. While FDA-approved for metastatic RCC and metastatic melanoma, its clinical application remains limited due to associated toxicity [252, 253]. This section delves into recent advancements and future directions in improving the efficacy of IL-2 therapy. The effectiveness of IL-2 therapy can be enhanced by implementing combination therapies. For instance, a phase I trial involving low-dose IL-2 and anti-PD-1 antibody nivolumab in patients with advanced solid tumors yielded a 30% overall response rate [253]. Notably, the toxicity experienced was manageable. Similarly, a combination of IL-2 and a cancer vaccine was evaluated in a phase I trial for patients with metastatic melanoma, which produced a response rate of 44% without significant toxicity [91].
As we continue to delve deeper into the molecular mechanisms and the vast cellular influences of IL-2, our understanding of its therapeutic potential in treating autoimmune diseases and cancer continues to expand. This section presents a comprehensive overview of future outlooks and perspectives, anchored in our current understanding of IL-2 and its applications in immunotherapy.
The regulation of IL-2 and IL-2R, being crucial in governing the immune response, is modulated by several transcription factors [35,36,37]. Hence, balancing these positive and negative regulators of IL-2 and its receptor is crucial for appropriate immune function. Subsequently, it follows that dysregulation can lead to autoimmune disorders and cancer, emphasizing the importance of understanding the biology, signaling pathways, transcription factors, and regulators of IL-2 and IL-2R [35,36,37,38,39,40]. In this regard, the IL-2/IL-2R signaling axis, with its profound impact on T-cell function and the overall immune response within the TME, represents a promising target for therapeutic intervention. In this context, it stands to reason that by strategically modulating this signaling pathway, there is potential to enhance the anti-tumor immune response and curtail tumor growth. Given this, understanding and manipulating the role of IL-2 and IL-2R in the regulation of T-cells within the TME may provide potent tools for bolstering the body’s natural defenses against cancer. Furthermore, IL-2 and IL-2R also play regulatory roles on various immune cells, including NK Cells, macrophages, neutrophils, and B Cells in the TME. They also interact significantly with DCs and MDSCs within the TME. Thus, it follows that future investigations should continue to probe the intricate roles and relationships of IL-2/IL-2R, with immune cells, DCs, and MDSCs in the TME. It is conceivable that this exploration may bolster the development of cancer immunotherapies. However, these strategies hinge on a complex and context-dependent relationship between IL-2/IL-2R signaling, DCs, and MDSCs in the TME. Hence, achieving a more profound understanding of this interplay and the mechanisms underpinning the effects of IL-2/IL-2R signaling on DCs and MDSCs is essential [148].
Notably, there is therapeutic potential in sIL-2R and IL-2Rα. Future advancements in cancer therapeutics involving these proteins are expected to encompass the development of novel formulations with improved efficacy and safety. Additionally, ongoing research efforts aim to identify reliable biomarkers of response to sIL-2R and IL-2Rα therapies [14, 23, 177,178,179,180,181,182,183,184,185,186,187,188,189,190,191,192,193,194,195,196,197,198, 200]. It is anticipated that such biomarkers could help predict which patients would benefit most from these therapies [201,202,203,204,205,206,207,208,209,210,211,212,213,214,215,216,217,218,219,220,221,222,223, 254, 255]. The burgeoning understanding of IL-2’s molecular mechanisms and cellular influences not only enhances its therapeutic potential in treating autoimmune diseases and cancer but also sets a new paradigm in immunotherapy [68, 71, 153, 226,227,228]. However, the challenge lies in fine-tuning the properties of engineered IL-2 to stimulate targeted immune responses without provoking adverse effects [109, 225, 240, 243, 245]. Accordingly, key developments encompass the engineering of IL-2 molecules that selectively bind to high-affinity IL-2 receptors on effector T cells and avoid low-affinity receptors on Tregs [68, 241].
In this regard, techniques involving receptor-targeted delivery systems, like anti-human IL-2 antibodies, mark a promising strategy in improving therapeutic potential and reducing systemic administration drawbacks [71, 224]. In turn, the evolution of IL-2 “superkines,“(discussed below) such as “NARA1,“ represents an achievement in IL-2 engineering, featuring enhanced IL-2 receptor binding and signaling properties [179, 232,233,234]. These innovative strategies aim to enhance therapeutic efficacy while minimizing toxicity, thus underscoring the potential of IL-2-based immunotherapies. However, potential autoimmune responses due to increased activity highlight the need for further investigation [234]. Significantly, the clinical success of engineered IL-2 molecules such as Aldesleukin, despite its limitations, reinforces the potential of IL-2 based therapies [153, 235, 236]. Other IL-2 molecules like NKTR-214 and AMG 592 show promise in preclinical trials, yet challenges of potential toxicities, manufacturing costs, and complex processes persist [68, 237, 243, 244]. Therefore, the integration of IL-2 therapy with other treatments, such as chimeric antigen receptor (CAR) T-cell therapy (discussed below) and anti-PD-1 antibodies, presents an intriguing landscape for innovative therapeutic strategies. These combined treatments in preliminary trials reveal promising outcomes, potentially addressing standalone IL-2 therapy limitations [253, 256, 257]. Consequently, to navigate the therapeutic potential and drawbacks of IL-2 molecules, research is ongoing in areas such as designing IL-2 therapeutics with enhanced in vivo half-lives, specific IL-2 receptor conformation targeting to stimulate particular T cell subsets, and localized therapies [241]. This concerted effort could lead to the evolution of more precise and effective IL-2-based therapies.
Interestingly, another burgeoning area of research is IL-2 therapy’s integration with other targeted therapies, such as tyrosine kinase inhibitors (TKIs) [10, 258, 259]. A preclinical study demonstrated improved anti-tumor activity when IL-2 was combined with a TKI targeting the mesenchymal-epithelial transition factor (MET) receptor in a mouse model of RCC [260]. Clinical trials are now underway to assess the combination of IL-2 and TKIs in patients with metastatic RCC [261, 262]. While IL-2-based therapies present certain challenges, they remain a key frontier in immunotherapy. Unquestionably, continuous research and clinical trials are expected to expand their scope and improve their therapeutic efficacy and safety [241, 251]. Despite the complexities, IL-2 therapies promise to significantly contribute to managing a broader range of cancer types and autoimmune diseases in the future..
Novel approaches and future perspectives
Receptor-gated IL-2 delivery via anti-human IL-2 antibody for regulatory T-cell activation
Receptor-gated IL-2 delivery is a unique therapeutic strategy that uses anti-human IL-2 antibodies to activate Tregs. In this technique, IL-2R functions as the ‘gate’, and the anti-human IL-2 antibody serves as the ‘key’ to unlock this gate. This selective delivery system ensures that IL-2 is selectively delivered to Tregs, promoting their activation and immunomodulatory functions [263].
The potential of receptor-gated IL-2 delivery to mitigate the side effects associated with conventional IL-2 administration, such as the unintended activation of various immune cells, makes it a promising therapeutic technique [263]. Furthermore, it has potential applications in situations where immune tolerance is compromised, like autoimmune diseases or transplantation [264]. A study that warrants particular attention utilized a cell-based and dynamic IL-2R platform to identify a distinct anti-human IL-2 antibody known as UFKA-20 [265]. UFKA-20 enabled selective and efficient stimulation of CD4 + Treg cells within freshly isolated human T cells ex vivo and in animal models in vivo [265]. However, it is crucial to calibrate this technique carefully to avoid excessive immune suppression that could potentially render the host vulnerable to infections or malignancies [71, 266].
Bispecific antibodies
Bispecific antibodies (bsAbs) represent a novel class of bioengineered molecules that can simultaneously engage two distinct antigenic epitopes. These uniquely dual-targeting agents demonstrate potential in revolutionizing cancer immunotherapy by concurrently interacting with IL-2R and tumor-associated antigens (TAAs), thereby bolstering T-cell mediated anti-tumor immunity [267,268,269,270]. Preclinical models have shown promising results with bsAbs enhancing the efficacy of receptor-gated IL-2 delivery and improving treatment outcomes in cancer [271,272,273]. For instance, in preclinical studies, a bsAb targeting PD-1 and LAG-3 exhibited enhanced T-cell activation and anti-tumor efficacy [272]. This dual-targeting approach of bsAbs provides a promising direction for cancer immunotherapy [274].
Multi-specific antibodies
Multi-specific antibodies (msAbs) can engage multiple antigens simultaneously, making them a valuable tool in cancer immunotherapy [268]. By targeting both IL-2 and co-stimulatory receptors like CD28 and 4-1BB, msAbs can enhance T-cell activation and proliferation, thereby improving the effectiveness of receptor-mediated IL-2 delivery and reinforcing anti-tumor immunity [152, 275]. Recent preclinical studies have shown promise in this area, demonstrating that msAbs can enhance anti-tumor immunity and the efficacy of receptor-gated IL-2 delivery [276, 277].
Fc receptor engineering
Fc receptor engineering involves modifications of the Fc region of anti-human IL-2 antibodies to augment antibody-dependent cell-mediated cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC). This can increase the efficacy of receptor-gated IL-2 delivery, potentially contributing to more effective therapeutic approaches in cancer management [278, 279]. Researchers continue investigating the potential of engineering the Fc region of anti-human IL-2 antibodies to improve ADCC and enhance receptor-gated IL-2 delivery [280,281,282,283].
Site-specific conjugation
Site-specific conjugation is attaching therapeutic molecules, such as drugs or toxins, to specific sites on an antibody molecule to create an antibody-drug conjugate (ADC). This technique can improve the selectivity and potency of cancer therapeutics by targeting specific antigens on tumor cells [41]. Site-specific conjugation has been used to develop several FDA-approved ADCs, such as ado-trastuzumab emtansine (Kadcyla) and brentuximab vedotin (Adcetris), for the treatment of HER2-positive breast cancer and CD30-positive lymphoma, respectively [42, 43].This method of attaching drugs or other molecules to specific sites on antibodies can improve their pharmacokinetic properties and reduce off-target effects. Recent studies have shown that site-specific conjugation can enhance the efficacy of receptor-gated IL-2 delivery and improve treatment outcomes in preclinical cancer models [44, 45].
Moreover, by enhancing the precision of IL-2 delivery through site-specific ADC conjugation, treatment outcomes can be improved significantly, as demonstrated by another recent preclinical cancer model [45, 46]. The aforementioned study underscores the potential of ADCs in cancer treatment, discussing the strategies for targeted drug delivery and how this precision may enhance the therapeutic potential of drugs [46]. These findings support the hypothesis that site-specific ADC conjugation can enhance IL-2 delivery, thereby improving treatment outcomes in cancer therapy. Further research in this domain is warranted to confirm these promising initial results and to explore possible applications in clinical settings.
CAR T cells and superkines
CAR T-cell therapy is a rapidly evolving therapeutic strategy that has been widely recognized for its role in harnessing the immune system to combat malignancies [90]. A prominent research interest lies in the modulation of CAR T-cell functions and their persistence in vivo through a combination therapy approach [284]. One such approach is integrating CAR T-cell therapy with IL-2 or IL-2R. This combined therapy has gained considerable attention in recent years due to its potential to enhance CAR T-cell performance and augment anti-tumor efficacy [285, 286]. In the context of CAR T-cell therapy, IL-2 can enhance the function of CAR T cells and improve their anti-tumor efficacy [287, 288]. Recent studies have shown that the co-administration of an anti-IL-2 antibody with CAR T cells improved CAR T cells’ persistence and function in a mouse glioblastoma model [289]. These advancements suggest that IL-2 or IL-2R-based therapies can impact the therapeutic success of CAR T-cell therapies.
In line with this, recent findings indicate that an orthogonal human IL-2 and IL-2Rβnsystem, termed Ortho-hIL-2, enhances CAR T cell expansion and antitumor activity in a murine model of leukemia [249]. This system not only boosts CAR T cells but also enhances their cytotoxicity and promotes their expansion, leading to leukemia regression [249]. Other orthogonal IL-2/IL-2R systems have been similarly found to enhance CAR T-cell therapy’s effectiveness, aiding in controlling CAR T-cell function, maximizing efficacy, and preventing acute graft-versus-host disease [290].
Given the growing body of evidence on IL-2/IL-2R’s role in enhancing CAR T-cell therapies, some researchers are exploring engineering CAR T cells to produce IL-2 within the tumor microenvironment [291]. This innovative approach could potentially improve the persistence and activation of CAR T cells, aiding in the clearance of bulky tumors [290, 291]. Further studies are necessary to validate this concept and determine its applicability in treating different cancer types.
However, it is important to consider that IL-2 therapies may also have limitations, such as promoting T cell exhaustion and influencing T cell differentiation [109, 290]. Therefore, fine-tuning the use of IL-2 in CAR T-cell therapies may be necessary to maximize benefits while minimizing adverse effects. On this note, targeted IL-2 variants have been shown to enhance CD8 + T-cell response, improve tumor control, and overcome resistance, suggesting that more personalized IL-2 therapies may be feasible [292,293,294].
Looking ahead, the integration of IL-2/IL-2R into CAR T-cell therapies holds substantial promise for cancer treatment. As research progresses, it will be important to continue exploring optimal combinations and doses, and develop strategies to mitigate potential side effects. This exciting frontier of cancer therapy stands to transform the treatment landscape for various types of cancer.
The field of cancer therapeutics has seen a paradigm shift with the introduction of superkines. Superkines are derived from diverse cytokine libraries and designed for enhanced biological potency, offering an optimistic avenue for cancer treatment [295]. A pivotal focus is the integration of IL-2 and IL-2R, which are promising therapeutic agents due to their critical role in immune cell function, cytotoxicity, and regulatory T cell expansion [109, 216]. However, designing IL-2 superkines has necessitated advanced computational methodologies to enhance affinity and stability [296]. An engineered IL-2 variant, MDNA109, was created to maximize antitumor effects while minimizing immune-related side effects [295]. This high-affinity, thermostable variant was shown to have an outward conformation that prearranges the IL-2Rβ binding site, optimizing its signaling properties [296]. Notably, the integration of IL-2 with its receptor, IL-2R, has been engineered for therapeutic proteins to bind more efficiently. This enhanced affinity for IL-2Rβois a key step in IL-2’s potency in immunotherapy [296]. IL-2’s interaction with IL-2R leads to signal cascades within the cell that promote proliferation and survival of effector T cells and NK cells, essential components in immune responses against tumors [109]. Further advancements include the development of fusion proteins and antibody complexes [109]. IL-2/anti-IL-2 antibody complexes, for instance, have been used to manipulate the immune system’s response to cancer by selectively expanding desired immune cell populations, such as CD4 regulatory T cells, and combating T cell exhaustion, a phenomenon in which T cells lose their functional capacities in chronic diseases like cancer [109]. Recent studies have demonstrated the potential for reshaping the TME using superkines, particularly with MDNA109. Delivered by an oncolytic adenovirus, MDNA109 shows superior anti-tumor responses in pancreatic cancer by enhancing immune cell activity and anti-tumor immune memory [297]. Interestingly, superkines like MDNA109 might be beneficial for treating immunologically “cold” tumors, known for their low mutation rates and limited immune cell infiltration [297]. Despite the promising prospects of superkines, certain issues need addressing. While high-dose IL-2 was previously used for melanoma and RCC treatment, its efficacy is questioned due to the rise of more targeted therapies [298]. Balancing the beneficial and detrimental effects of IL-2-based therapies continues to be a challenge in this field. Looking ahead, continuous research aims to optimize the potential of IL-2/IL-2R integration in superkines. Future perspectives include refining the therapeutic application of superkines through in silico affinity maturation and structure stabilization strategies, advancing clinical trials for cancer and autoimmune diseases, and targeting IL-2 to specific tissues for a more precise therapeutic approach [80, 296, 298]. The realization of these prospects may hold the key to transforming cancer therapeutics, making superkines a beacon of hope in the fight against this relentless disease.
Current limitations and challenges of IL-2-based therapies
IL-2 is a key cytokine in the gamma (c) family, with critical roles in the TME and various therapeutic applications. Gamma (c) cytokines, including IL-4, IL-7, IL-9, IL-15, and IL-21, are vital for immune regulation, with shared use of IL-2Rs across these cytokines contributing to their functional overlap and redundancy [299]. This receptor sharing could be a double-edged sword in therapeutic utilization, as it allows for broad immunomodulatory effects but could potentially lead to unintended off-target effects. IL-2’s primary role in the TME is to drive the proliferation and activation of cytotoxic T cells, effectively aiding in the elimination of tumor cells [300]. Moreover, IL-2 has been widely utilized in immunotherapy for its ability to promote the expansion and function of Tregs, which are instrumental in maintaining immune homeostasis and preventing autoimmunity [23]. However, the pleiotropic nature of IL-2 can also contribute to adverse effects such as vascular leak syndrome, presenting a challenge to its therapeutic use [91]. Comparative studies of gamma (c) cytokines in immunotherapeutic settings have demonstrated distinctive advantages and limitations. Markley and Sadelain (2010) highlighted how different gamma (c) cytokines, despite sharing IL-2Rs, can preferentially promote the expansion of distinct immune cell subsets [301]. For instance, IL-7 and IL-15, unlike IL-2, primarily support memory T cell survival and proliferation, offering potential benefits in long-term tumor control. However, the broad receptor sharing across these cytokines might also lead to the activation of unwanted cell populations, potentially aggravating immune-related adverse events. While IL-2 and other gamma (c) cytokines play pivotal roles in the TME and hold substantial promise in cancer immunotherapy, the therapeutic exploitation of these cytokines requires a delicate balance of maximizing antitumor efficacy while minimizing off-target effects and toxicity.
IL-2 and IL-2R-based therapies have emerged as significant breakthroughs in cancer immunotherapy, their role rooted in IL-2’s crucial functions in T cell biology and immune regulation [302]. Demonstrating marked efficacy in conditions like metastatic melanoma, renal cell carcinoma, and synovial sarcoma, they have further been enhanced by the advent of new IL-2 formulations such as Alb-IL2 and IL2-Fc, providing improved clinical outcomes [91, 93, 303]. However, despite these promising advancements, IL-2-based therapies are not devoid of limitations. IL-2’s inherent toxicity, notably its association with VLS, can impose restrictions on its therapeutic applications [10]. Factors such as angiopoietin 2 and endothelial nitric oxide synthase have been implicated in IL-2-induced VLS [93]. The influence of Tregs also critically determines the effectiveness of IL-2-based therapies. While IL-2 augments T cell responses, it concurrently promotes Tregs expansion, which can counteract anti-tumor immune responses [10, 303]. This expansion of Tregs could therefore detrimentally affect the efficacy of IL-2-based cancer immunotherapies. Moreover, the intricate administration protocols and restricted applicability of IL-2 therapy contribute to its limitations, as evidenced in the treatment of mRCC [303]. Another significant factor influencing the success of these therapies is the individual tumor immune microenvironment. Variability in response rates among melanoma patients undergoing immunotherapy may be attributable to differences in their specific tumor immune microenvironments [91]. Efforts are underway to improve the safety and efficacy of IL-2 based therapies by altering the molecule itself or by changing the way it’s administered. (Supplementary Table 1 [291, 304,305,306,307,308,309,310,311,312,313,314,315,316,317,318] summarizes the novel approaches for cancer immunotherapy).
Summary
IL-2 is a cytokine crucial for the activation and proliferation of immune cells, including T cells and natural killer cells. In the tumor microenvironment, IL-2 and its IL-2R have complex and sometimes opposing roles in tumor progression and immune response. While IL-2 can stimulate immune cells to attack tumors, it can also promote the expansion of regulatory T cells that suppress anti-tumor immunity. Hence, maintaining the balance between IL-2 and IL-2R signaling is critical for effective anti-tumor immunity. In addition, research has demonstrated that various cancer types exhibit different levels of IL-2 and IL-2R expression, which can impact their response to immunotherapy. For example, melanoma and renal cell carcinoma have been shown to have high levels of IL-2R expression, making them more responsive to IL-2-based immunotherapy.
Moreover, soluble IL-2R and IL-2 alpha have also been examined as potential biomarkers for cancer diagnosis, treatment, and prognosis. Engineered forms of IL-2 have been developed to improve its anti-tumor activity while reducing toxicity. For example, pegylated IL-2 has been demonstrated to possess a longer half-life and greater efficacy than native IL-2. Clinical trials have investigated the use of IL-2 alone or in combination with other agents, such as checkpoint inhibitors, in treating various types of cancer, including melanoma and renal cell carcinoma.
Although IL-2-based therapies have exhibited promise in some cancer types, the complex interplay between IL-2 and the tumor microenvironment necessitates further investigation. In addition, future research may focus on optimizing dosing and combination strategies to enhance the effectiveness of IL-2-based immunotherapy and identifying patient populations that are most likely to benefit from this approach. Overall, the study of IL-2 and its receptor in cancer holds excellent potential for developing new and effective treatments for a wide range of malignancies.
Conclusion
In conclusion, the role of IL-2 and IL-2R within the tumor microenvironment remains a fascinating and vital area of exploration. Their multifaceted interplay—mediating both immune activation and regulation—has profound implications on the immune response to various cancers, suggesting that their targeted manipulation holds promise for improving cancer immunotherapy outcomes. Emerging evidence supports diverse IL-2 and IL-2R expression levels across different cancer types, potentially shaping their responses to immunotherapy. This heterogeneity underscores the need for a personalized approach to IL-2-based immunotherapy, with future strategies potentially requiring tailoring based on individual patient characteristics and tumor profiles. The development and optimization of engineered forms of IL-2, such as pegylated IL-2, mark an exciting advancement in this field. These novel forms promise to enhance therapeutic efficacy while curbing systemic toxicity.
Furthermore, exploring combination therapies—IL-2/IL-2R-targeted therapies coupled with traditional cancer treatments like chemotherapy, radiotherapy, or other immunotherapies like checkpoint inhibitors—could optimize cancer treatment efficacy. As we continue to investigate the role of IL-2 and IL-2R in tumorigenesis, there is also growing interest in their potential as diagnostic, prognostic, and monitoring biomarkers in cancer. Their utility in this regard could provide invaluable insights to inform therapeutic selection and timing. While substantial progress has been made, much remains to be understood about the intricate dance between IL-2 and the tumor microenvironment. Future research should strive to elucidate this complexity and translate these insights into more effective, safe, and patient-specific therapies. As we navigate this challenging yet promising landscape, the full therapeutic potential of IL-2 and IL-2R-targeted treatments in cancer immunotherapy comes into sharper focus. Through this ongoing scientific exploration, we may ultimately improve cancer patient outcomes and transform the future of cancer treatment.
Data availability
All data generated or analyzed during this study are included in this published article and its additional information files.
Abbreviations
- ADCC:
-
Antibody-dependent cell-mediated cytotoxicity
- ADC:
-
Antibody-drug conjugate
- AMG 592:
-
An Engineered IL-2 Molecule
- bsAbs:
-
Bispecific antibodies
- CAR:
-
Chimeric Antigen Receptor
- CDC:
-
Complement-dependent cytotoxicity
- CD122:
-
Interleukin-2 receptor subunit beta
- CD132:
-
Interleukin-2 receptor subunit gamma
- CD25:
-
Interleukin-2 receptor subunit alpha
- CD28:
-
Cluster of Differentiation 28, a co-stimulatory protein
- CD4:
-
Cluster of Differentiation 4, a glycoprotein found on immune cells
- CTL:
-
Cytotoxic T Lymphocyte
- CTLA-4:
-
Cytotoxic T-Lymphocyte-Associated Protein 4
- FDA:
-
Food and Drug Administration
- Fc:
-
Fragment crystallizable region
- HER2:
-
Human epidermal growth factor receptor 2
- IFN-γ:
-
Interferon Gamma
- IL-2:
-
Interleukin-2
- IL-2R:
-
Interleukin-2 Receptor
- IL-2Rα:
-
Interleukin-2 Receptor Alpha
- IL-2Rβ:
-
Interleukin-2 Receptor Beta
- IL-2Rγ:
-
Interleukin-2 Receptor Gamma
- IL-2/UFKA-20 complex:
-
A complex formed between IL-2 and the anti-human IL-2 antibody UFKA-20
- JAK:
-
Janus Kinase
- LAG-3:
-
Lymphocyte-activation gene 3
- mAbs:
-
Monoclonal Antibodies
- msAbs:
-
Multi-specific antibodies
- NARA1:
-
Not an actual abbreviation, but a specific type of Superkine molecule
- NK:
-
Natural Killer (Cells)
- NKTR-214:
-
A type of Engineered IL-2 Molecule
- PD-1:
-
Programmed cell death protein 1
- rIL-2:
-
Recombinant Interleukin-2
- STAT:
-
Signal Transducer and Activator of Transcription
- TAA:
-
Tumor-associated antigen
- TCR:
-
T-Cell Receptor
- TGF-β:
-
Transforming Growth Factor Beta
- TIL:
-
Tumor Infiltrating Lymphocyte
- TME:
-
Tumor Microenvironment
- Tregs:
-
Regulatory T Cells
- VEGF:
-
Vascular Endothelial Growth Factor
- VLS:
-
Vascular Leak Syndrome
References
Bachmann MF, Oxenius A. Interleukin 2: from immunostimulation to immunoregulation and back again. EMBO Rep. 2007;8(12):1142–8.
Waldman T. The interleukin-2 receptor. J Biol Chem (Print). 1991;266(5):2681–4.
Waters RS, Perry JSA, Han S, Bielekova B, Gedeon T. The effects of interleukin-2 on immune response regulation. Math Med Biol. 2018;35(1):79–119.
Maeda M, Tanabe-Shibuya J, Miyazato P, Masutani H, Yasunaga JI, Usami K, et al. IL-2/IL-2 receptor pathway plays a crucial role in the Growth and Malignant Transformation of HTLV-1-Infected T cells to develop adult T-Cell leukemia. Front Microbiol. 2020;11:356.
Yu H, Lee H, Herrmann A, Buettner R, Jove R. Revisiting STAT3 signalling in cancer: new and unexpected biological functions. Nat Rev Cancer. 2014;14(11):736–46.
Atkins MB, Lotze MT, Dutcher JP, Fisher RI, Weiss G, Margolin K, et al. High-dose recombinant interleukin 2 therapy for patients with metastatic melanoma: analysis of 270 patients treated between 1985 and 1993. J Clin Oncol. 1999;17(7):2105–16.
Atkins MB, Kunkel L, Sznol M, Rosenberg SA. High-dose recombinant interleukin-2 therapy in patients with metastatic melanoma: long-term survival update. Cancer J Sci Am. 2000;6(Suppl 1):11–4.
Yang JC, Sherry RM, Steinberg SM, Topalian SL, Schwartzentruber DJ, Hwu P, et al. Randomized study of high-dose and low-dose interleukin-2 in patients with metastatic renal cancer. J Clin Oncol. 2003;21(16):3127–32.
Ahmadzadeh M, Rosenberg SA. IL-2 administration increases CD4 + CD25(hi) Foxp3 + regulatory T cells in cancer patients. Blood. 2006;107(6):2409–14.
Jiang T, Zhou C, Ren S. Role of IL-2 in cancer immunotherapy. Oncoimmunology. 2016;5(6):e1163462.
Considine B, Hurwitz ME. Current status and future directions of Immunotherapy in Renal Cell Carcinoma. Curr Oncol Rep. 2019;21(4):34.
Rosenberg SA, Lotze MT, Muul LM, Leitman S, Chang AE, Ettinghausen SE, et al. Observations on the systemic administration of autologous lymphokine-activated killer cells and recombinant interleukin-2 to patients with metastatic cancer. N Engl J Med. 1985;313(23):1485–92.
Waldmann TA. The biology of interleukin-2 and interleukin-15: implications for cancer therapy and vaccine design. Nat Rev Immunol. 2006;6(8):595–601.
Wang X, Rickert M, Garcia KC. Structure of the quaternary complex of interleukin-2 with its alpha, beta, and gammac receptors. Science. 2005;310(5751):1159–63.
Mondino A, Mueller DL. mTOR at the crossroads of T cell proliferation and tolerance. Semin Immunol. 2007;19(3):162–72.
Lee MS, Kim YJ. Signaling pathways downstream of pattern-recognition receptors and their cross talk. Annu Rev Biochem. 2007;76:447–80.
Charych DH, Hoch U, Langowski JL, Lee SR, Addepalli MK, Kirk PB, et al. NKTR-214, an Engineered Cytokine with biased IL2 receptor binding, increased tumor exposure, and marked efficacy in mouse Tumor Models. Clin Cancer Res. 2016;22(3):680–90.
Boyman O, Kovar M, Rubinstein MP, Surh CD, Sprent J. Selective stimulation of T cell subsets with antibody-cytokine immune complexes. Science. 2006;311(5769):1924–7.
Xin Yu J, Hubbard-Lucey VM, Tang J. The global pipeline of cell therapies for cancer. Nat Rev Drug Discov. 2019;18(11):821–2.
Choudhry H, Helmi N, Abdulaal WH, Zeyadi M, Zamzami MA, Wu W, et al. Prospects of IL-2 in Cancer Immunotherapy. Biomed Res Int. 2018;2018:9056173.
Kean LS, Turka LA, Blazar BR. Advances in targeting co-inhibitory and co-stimulatory pathways in transplantation settings: the Yin to the Yang of cancer immunotherapy. Immunol Rev. 2017;276(1):192–212.
Peterson C, Denlinger N, Yang Y. Recent advances and Challenges in Cancer Immunotherapy. Cancers (Basel). 2022;14(16).
Malek TR, Castro I. Interleukin-2 receptor signaling: at the interface between tolerance and immunity. Immunity. 2010;33(2):153–65.
Smith KA. Interleukin-2: inception, impact, and implications. Science. 1988;240(4856):1169–76.
Waldmann TA. The structure, function, and expression of interleukin-2 receptors on normal and malignant lymphocytes. Science. 1986;232(4751):727–32.
Leonard WJ, Lin JX, O’Shea JJ. The gamma(c) family of Cytokines: Basic Biology to therapeutic ramifications. Immunity. 2019;50(4):832–50.
Taniguchi T, Takaoka A. The interferon-alpha/beta system in antiviral responses: a multimodal machinery of gene regulation by the IRF family of transcription factors. Curr Opin Immunol. 2002;14(1):111–6.
Fruman DA, Cantley LC, Carpenter CL. Structural organization and alternative splicing of the murine phosphoinositide 3-kinase p85 alpha gene. Genomics. 1996;37(1):113–21.
Boyman O, Sprent J. The role of interleukin-2 during homeostasis and activation of the immune system. Nat Rev Immunol. 2012;12(3):180–90.
Abbas AK, Murphy KM, Sher A. Functional diversity of helper T lymphocytes. Nature. 1996;383(6603):787–93.
Lanzavecchia A, Sallusto F. Dynamics of T lymphocyte responses: intermediates, effectors, and memory cells. Science. 2000;290(5489):92–7.
Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J Immunol. 1995;155(3):1151–64.
Vivier E, Raulet DH, Moretta A, Caligiuri MA, Zitvogel L, Lanier LL, et al. Innate or adaptive immunity? The example of natural killer cells. Science. 2011;331(6013):44–9.
Cooper MA, Fehniger TA, Turner SC, Chen KS, Ghaheri BA, Ghayur T, et al. Human natural killer cells: a unique innate immunoregulatory role for the CD56(bright) subset. Blood. 2001;97(10):3146–51.
Macian F. NFAT proteins: key regulators of T-cell development and function. Nat Rev Immunol. 2005;5(6):472–84.
Zhu J, Paul WE. Peripheral CD4 + T-cell differentiation regulated by networks of cytokines and transcription factors. Immunol Rev. 2010;238(1):247–62.
Johnston JA, Bacon CM, Finbloom DS, Rees RC, Kaplan D, Shibuya K, et al. Tyrosine phosphorylation and activation of STAT5, STAT3, and Janus kinases by interleukins 2 and 15. Proc Natl Acad Sci U S A. 1995;92(19):8705–9.
Yasukawa H, Sasaki A, Yoshimura A. Negative regulation of cytokine signaling pathways. Annu Rev Immunol. 2000;18:143–64.
Sakaguchi S, Yamaguchi T, Nomura T, Ono M. Regulatory T cells and immune tolerance. Cell. 2008;133(5):775–87.
Ruan Q, Kameswaran V, Tone Y, Li L, Liou HC, Greene MI, et al. Development of Foxp3(+) regulatory t cells is driven by the c-Rel enhanceosome. Immunity. 2009;31(6):932–40.
Crabtree GR. Generic signals and specific outcomes: signaling through Ca2+, calcineurin, and NF-AT. Cell. 1999;96(5):611–4.
Beals CR, Sheridan CM, Turck CW, Gardner P, Crabtree GR. Nuclear export of NF-ATc enhanced by glycogen synthase kinase-3. Science. 1997;275(5308):1930–3.
Rincon M, Enslen H, Raingeaud J, Recht M, Zapton T, Su MS, et al. Interferon-γ expression by Th1 effector T cells mediated by the p38 MAP kinase signaling pathway. EMBO J. 1998;17(10):2817–29.
Katagiri T, Kameda H, Nakano H, Yamazaki S. Regulation of T cell differentiation by the AP-1 transcription factor JunB. Immunological Med. 2021;44(3):197–203.
Baldwin AS Jr. The NF-κB and IκB proteins: new discoveries and insights. Annu Rev Immunol. 1996;14(1):649–81.
Spolski R, Li P, Leonard WJ. Biology and regulation of IL-2: from molecular mechanisms to human therapy. Nat Rev Immunol. 2018;18(10):648–59.
Decker EL, Skerka C, Zipfel PF. The early growth response protein (EGR-1) regulates interleukin-2 transcription by synergistic interaction with the nuclear factor of activated T cells. J Biol Chem. 1998;273(41):26923–30.
Jadhav K, Zhang Y. Activating transcription factor 3 in immune response and metabolic regulation. Liver Res. 2017;1(2):96–102.
Leonard WJ, O’Shea JJ. Jaks and STATs: biological implications. Annu Rev Immunol. 1998;16(1):293–322.
Tsuji-Takayama K, Suzuki M, Yamamoto M, Harashima A, Okochi A, Otani T, et al. IL-2 activation of STAT5 enhances production of IL-10 from human cytotoxic regulatory T cells, HOZOT. Exp Hematol. 2008;36(2):181–92.
Rani A, Murphy JJ. STAT5 in cancer and immunity. J Interferon Cytokine Res. 2016;36(4):226–37.
Krebs DL, Hilton DJ. SOCS proteins: negative regulators of cytokine signaling. Stem Cells. 2001;19(5):378–87.
Linossi EM, Babon JJ, Hilton DJ, Nicholson SE. Suppression of cytokine signaling: the SOCS perspective. Cytokine Growth Factor Rev. 2013;24(3):241–8.
Krebs DL, Hilton DJ. SOCS: physiological suppressors of cytokine signaling. J Cell Sci. 2000;113(16):2813–9.
Zheng SG, Wang J, Wang P, Gray JD, Horwitz DA. IL-2 is essential for TGF-β to convert naive CD4 + CD25 – cells to CD25 + Foxp3 + regulatory T cells and for expansion of these cells. J Immunol. 2007;178(4):2018–27.
Wuest TY, Willette-Brown J, Durum SK, Hurwitz AA. The influence of IL-2 family cytokines on activation and function of naturally occurring regulatory T cells. J Leucocyte Biology. 2008;84(4):973–80.
Fontenot JD, Gavin MA, Rudensky AY. Foxp3 programs the development and function of CD4 + CD25 + regulatory T cells. Nat Immunol. 2003;4(4):330–6.
Pandiyan P, Zheng L, Ishihara S, Reed J, Lenardo MJ. CD4 + CD25 + Foxp3 + regulatory T cells induce cytokine deprivation–mediated apoptosis of effector CD4 + T cells. Nat Immunol. 2007;8(12):1353–62.
Mamontov P, Eberwine RA, Perrigoue J, Das A, Friedman JR, Mora JR. A negative role for the interleukin-2-inducible T-cell kinase (ITK) in human Foxp3 + TREG differentiation. PLoS ONE. 2019;14(4):e0215963.
Gomez-Rodriguez J, Wohlfert EA, Handon R, Meylan F, Wu JZ, Anderson SM, et al. Itk-mediated integration of T cell receptor and cytokine signaling regulates the balance between Th17 and regulatory T cells. J Exp Med. 2014;211(3):529–43.
Mammadli M, Harris R, Suo L, May A, Gentile T, Waickman AT, et al. Interleukin-2‐inducible T‐cell kinase (itk) signaling regulates potent noncanonical regulatory T cells. Clin Translational Med. 2021;11(12):e625.
Ghosh S, Bienemann K, Boztug K, Borkhardt A. Interleukin-2-inducible T-cell kinase (ITK) deficiency-clinical and molecular aspects. J Clin Immunol. 2014;34:892–9.
Hayashi K, Altman A. Protein kinase C theta (PKCθ): a key player in T cell life and death. Pharmacol Res. 2007;55(6):537–44.
Ozay EI, Shanthalingam S, Sherman HL, Torres JA, Osborne BA, Tew GN, et al. Cell-penetrating anti-protein kinase C theta antibodies act intracellularly to generate stable, highly suppressive regulatory T cells. Mol Ther. 2020;28(9):1987–2006.
Ghaffari-Tabrizi N, Bauer B, Villunger A, Baier‐Bitterlich G, Altman A, Utermann G, et al. Protein kinase Cθ, a selective upstream regulator of JNK/SAPK and IL‐2 promoter activation in Jurkat T cells. Eur J Immunol. 1999;29(1):132–42.
Sakaguchi S, Miyara M, Costantino CM, Hafler DA. FOXP3 + regulatory T cells in the human immune system. Nat Rev Immunol. 2010;10(7):490–500.
Muranski P, Boni A, Wrzesinski C, Citrin DE, Rosenberg SA, Childs R, et al. Increased intensity lymphodepletion and adoptive immunotherapy–how far can we go? Nat Clin Pract Oncol. 2006;3(12):668–81.
Raeber ME, Sahin D, Karakus U, Boyman O. A systematic review of interleukin-2-based immunotherapies in clinical trials for cancer and autoimmune diseases. EBioMedicine. 2023;90.
Graßhoff H, Comdühr S, Monne LR, Müller A, Lamprecht P, Riemekasten G, et al. Low-dose IL-2 therapy in autoimmune and rheumatic diseases. Front Immunol. 2021;12:648408.
Solomon I, Amann M, Goubier A, Arce Vargas F, Zervas D, Qing C, et al. CD25-Treg-depleting antibodies preserving IL-2 signaling on effector T cells enhance effector activation and antitumor immunity. Nat cancer. 2020;1(12):1153–66.
Karakus U, Sahin D, Mittl PR, Mooij P, Koopman G, Boyman O. Receptor-gated IL-2 delivery by an anti-human IL-2 antibody activates regulatory T cells in three different species. Sci Transl Med. 2020;12(574):eabb9283.
Wang D-R, Wu X-L, Sun Y-L. Therapeutic targets and biomarkers of tumor immunotherapy: response versus non-response. Signal Transduct Target Therapy. 2022;7(1):331.
Li J-Y, Chen Y-P, Li Y-Q, Liu N, Ma J. Chemotherapeutic and targeted agents can modulate the tumor microenvironment and increase the efficacy of immune checkpoint blockades. Mol Cancer. 2021;20:1–21.
Mortara L, Balza E, Bruno A, Poggi A, Orecchia P, Carnemolla B. Anti-cancer therapies employing IL-2 cytokine tumor targeting: contribution of innate, adaptive and immunosuppressive cells in the anti-tumor efficacy. Front Immunol. 2018;9:2905.
Morris JC, Waldmann TA. Advances in interleukin 2 receptor targeted treatment. Ann Rheum Dis. 2000;59(suppl 1):i109–i14.
Li Y, Xiang S, Pan W, Wang J, Zhan H, Liu S. Targeting tumor immunosuppressive microenvironment for pancreatic cancer immunotherapy: current research and future perspective. Front Oncol. 2023;13.
Zhou W-T, Jin W-L. B7-H3/CD276: an emerging cancer immunotherapy. Front Immunol. 2021;12:701006.
Zhong L, Li Y, Xiong L, Wang W, Wu M, Yuan T, et al. Small molecules in targeted cancer therapy: advances, challenges, and future perspectives. Signal Transduct Target therapy. 2021;6(1):201.
Liu GH, Chen T, Zhang X, Ma XL, Shi HS. Small molecule inhibitors targeting the cancers. MedComm. 2022;3(4):e181.
Merchant R, Galligan C, Munegowda MA, Pearce LB, Lloyd P, Smith P et al. Fine-tuned long-acting interleukin-2 superkine potentiates durable immune responses in mice and non-human primate. J Immunother Cancer. 2022;10(1).
Dutcher JP, Schwartzentruber DJ, Kaufman HL, Agarwala SS, Tarhini AA, Lowder JN, et al. High dose interleukin-2 (Aldesleukin)-expert consensus on best management practices-2014. J Immunother Cancer. 2014;2:1–23.
Abdou Y, Pandey M, Sarma M, Shah S, Baron J, Ernstoff MS. Mechanism-based treatment of cancer with immune checkpoint inhibitor therapies. Br J Clin Pharmacol. 2020;86(9):1690–702.
Milling L, Zhang Y, Irvine DJ. Delivering safer immunotherapies for cancer. Adv Drug Deliv Rev. 2017;114:79–101.
Tähtinen S, Kaikkonen S, Merisalo-Soikkeli M, Grönberg-Vähä-Koskela S, Kanerva A, Parviainen S, et al. Favorable alteration of tumor microenvironment by immunomodulatory cytokines for efficient T-cell therapy in solid tumors. PLoS ONE. 2015;10(6):e0131242.
Abdou P, Wang Z, Chen Q, Chan A, Zhou DR, Gunadhi V, et al. Advances in engineering local drug delivery systems for cancer immunotherapy. Wiley Interdisciplinary Reviews: Nanomedicine and Nanobiotechnology. 2020;12(5):e1632.
Parnsamut C, Brimson S. Effects of silver nanoparticles and gold nanoparticles on IL-2, IL-6, and TNF-α production via MAPK pathway in leukemic cell lines. Genet Mol Res. 2015;14(2):3650–68.
Pires IS, Hammond PT, Irvine DJ. Engineering strategies for immunomodulatory cytokine therapies: challenges and clinical progress. Adv Ther. 2021;4(8):2100035.
Bell M, Gottschalk S. Engineered cytokine signaling to improve CAR T cell effector function. Front Immunol. 2021;12:684642.
Labbé RP, Vessillier S, Rafiq QA. Lentiviral vectors for T cell engineering: clinical applications, bioprocessing and future perspectives. Viruses. 2021;13(8):1528.
Jogalekar MP, Rajendran RL, Khan F, Dmello C, Gangadaran P, Ahn B-C. CAR T-cell-based gene therapy for cancers: new perspectives, challenges, and clinical developments. Front Immunol. 2022;13.
Rosenberg SA. IL-2: the first effective immunotherapy for human cancer. J Immunol. 2014;192(12):5451–8.
Levine BL, Miskin J, Wonnacott K, Keir C. Global manufacturing of CAR T cell therapy. Mol Therapy-Methods Clin Dev. 2017;4:92–101.
MacDonald A, Wu T-C, Hung C-F. Interleukin 2-based fusion proteins for the treatment of cancer. J Immunol Res. 2021;2021:1–11.
Jing H, Hettich M, Gaedicke S, Firat E, Bartholomä M, Niedermann G. Combination treatment with hypofractionated radiotherapy plus IL-2/anti-IL-2 complexes and its theranostic evaluation. J Immunother Cancer. 2019;7:1–16.
Richards JM, Gale D, Mehta N, Lestingi T. Combination of chemotherapy with interleukin-2 and interferon alfa for the treatment of metastatic melanoma. J Clin Oncol. 1999;17(2):651.
Rosenberg SA, Yang JC, Schwartzentruber DJ, Hwu P, Marincola FM, Topalian SL, et al. Prospective randomized trial of the treatment of patients with metastatic melanoma using chemotherapy with cisplatin, dacarbazine, and tamoxifen alone or in combination with interleukin-2 and interferon alfa-2b. J Clin Oncol. 1999;17(3):968.
Chen DS, Mellman I. Oncology meets immunology: the cancer-immunity cycle. Immunity. 2013;39(1):1–10.
Fridman WH, Pages F, Sautes-Fridman C, Galon J. The immune contexture in human tumours: impact on clinical outcome. Nat Rev Cancer. 2012;12(4):298–306.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–74.
Zou W. Immunosuppressive networks in the tumour environment and their therapeutic relevance. Nat Rev Cancer. 2005;5(4):263–74.
Schreiber RD, Old LJ, Smyth MJ. Cancer immunoediting: integrating immunity’s roles in cancer suppression and promotion. Science. 2011;331(6024):1565–70.
Wherry EJ, Kurachi M. Molecular and cellular insights into T cell exhaustion. Nat Rev Immunol. 2015;15(8):486–99.
Pardoll DM. The blockade of immune checkpoints in cancer immunotherapy. Nat Rev Cancer. 2012;12(4):252–64.
Dong Y, Sun Q, Zhang X. PD-1 and its ligands are important immune checkpoints in cancer. Oncotarget. 2017;8(2):2171–86.
Wang W, Kryczek I, Dostal L, Lin H, Tan L, Zhao L, et al. Effector T cells abrogate stroma-mediated Chemoresistance in Ovarian Cancer. Cell. 2016;165(5):1092–105.
Lan RY, Selmi C, Gershwin ME. The regulatory, inflammatory, and T cell programming roles of interleukin-2 (IL-2). J Autoimmun. 2008;31(1):7–12.
Yurchenko E, Shio MT, Huang TC, Da Silva Martins M, Szyf M, Levings MK, et al. Inflammation-driven reprogramming of CD4 + Foxp3 + regulatory T cells into pathogenic Th1/Th17 T effectors is abrogated by mTOR inhibition in vivo. PLoS ONE. 2012;7(4):e35572.
Basu A, Ramamoorthi G, Albert G, Gallen C, Beyer A, Snyder C, et al. Differentiation and regulation of TH cells: a balancing act for cancer immunotherapy. Front Immunol. 2021;12:669474.
Hernandez R, Põder J, LaPorte KM, Malek TR. Engineering IL-2 for immunotherapy of autoimmunity and cancer. Nat Rev Immunol. 2022;22(10):614–28.
Lisiero DN, Soto H, Liau LM, Prins RM. Enhanced sensitivity to IL-2 signaling regulates the clinical responsiveness of IL-12–primed CD8 + T cells in a melanoma model. J Immunol. 2011;186(9):5068–77.
Kolenko V, Wang Q, Riedy M, O’Shea J, Ritz J, Cathcart MK et al. Tumor-induced suppression of T lymphocyte proliferation coincides with inhibition of Jak3 expression and IL-2 receptor signaling: role of soluble products from human renal cell carcinomas. Journal of immunology (Baltimore, Md: 1950). 1997;159(6):3057-67.
García-Tuñón I, Ricote M, Ruiz A, Fraile B, Paniagua R, Royuela M. Interleukin-2 and its receptor complex (α, β and γ chains) in in situand infiltrative human breast cancer: an immunohistochemical comparative study. Breast Cancer Res. 2003;6(1):1–7.
Liao W, Lin J-X, Leonard WJ. Interleukin-2 at the crossroads of effector responses, tolerance, and immunotherapy. Immunity. 2013;38(1):13–25.
Quail DF, Joyce JA. Microenvironmental regulation of tumor progression and metastasis. Nat Med. 2013;19(11):1423–37.
Nelson BH. IL-2, regulatory T cells, and tolerance. J Immunol. 2004;172(7):3983–8.
Huang Z, Kang SG, Li Y, Zak J, Shaabani N, Deng K, et al. IFNAR1 signaling in NK cells promotes persistent virus infection. Sci Adv. 2021;7(13):eabb8087.
Liao D, Luo Y, Markowitz D, Xiang R, Reisfeld RA. Cancer associated fibroblasts promote tumor growth and metastasis by modulating the tumor immune microenvironment in a 4T1 murine breast cancer model. PLoS ONE. 2009;4(11):e7965.
Mantovani A, Cassatella MA, Costantini C, Jaillon S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat Rev Immunol. 2011;11(8):519–31.
Pillay J, Kamp VM, Van Hoffen E, Visser T, Tak T, Lammers J-W, et al. A subset of neutrophils in human systemic inflammation inhibits T cell responses through Mac-1. J Clin Investig. 2012;122(1):327–36.
Braumüller H, Mauerer B, Andris J, Berlin C, Wieder T, Kesselring R. The Cytokine Network in Colorectal Cancer: implications for New Treatment Strategies. Cells. 2023;12(1):138.
Shen G, Krienke S, Schiller P, Nießen A, Neu S, Eckstein V, et al. Microvesicles released by apoptotic human neutrophils suppress proliferation and IL-2/IL‐2 receptor expression of resting T helper cells. Eur J Immunol. 2017;47(5):900–10.
Zhou Y, Cheng L, Liu L, Li X. NK cells are never alone: crosstalk and communication in tumour microenvironments. Mol Cancer. 2023;22(1):34.
Sharma R, Fu SM, Ju S-T. IL-2: a two-faced master regulator of autoimmunity. J Autoimmun. 2011;36(2):91–7.
Mantovani A, Sozzani S, Locati M, Allavena P, Sica A. Macrophage polarization: tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002;23(11):549–55.
Chinen T, Kannan AK, Levine AG, Fan X, Klein U, Zheng Y, et al. An essential role for the IL-2 receptor in Treg cell function. Nat Immunol. 2016;17(11):1322–33.
Levin AM, Bates DL, Ring AM, Krieg C, Lin JT, Su L, et al. Exploiting a natural conformational switch to engineer an interleukin-2 ‘superkine’. Nature. 2012;484(7395):529–33.
Sharma P, Allison JP. The future of immune checkpoint therapy. Science. 2015;348(6230):56–61.
Fife BT, Bluestone JA. Control of peripheral T-cell tolerance and autoimmunity via the CTLA‐4 and PD‐1 pathways. Immunol Rev. 2008;224(1):166–82.
Rouse M, Nagarkatti M, Nagarkatti PS. The role of IL-2 in the activation and expansion of regulatory T-cells and the development of experimental autoimmune encephalomyelitis. Immunobiology. 2013;218(4):674–82.
Wing JB, Tanaka A, Sakaguchi S. Human FOXP3 + regulatory T cell heterogeneity and function in autoimmunity and cancer. Immunity. 2019;50(2):302–16.
Zhang Y, Zhang Z. The history and advances in cancer immunotherapy: understanding the characteristics of tumor-infiltrating immune cells and their therapeutic implications. Cell Mol Immunol. 2020;17(8):807–21.
Quezada SA, Peggs KS, Simpson TR, Allison JP. Shifting the equilibrium in cancer immunoediting: from tumor tolerance to eradication. Immunol Rev. 2011;241(1):104–18.
Flynn NJ, Somasundaram R, Arnold KM, Sims-Mourtada J. The multifaceted roles of B cells in solid tumors: emerging treatment opportunities. Target Oncol. 2017;12:139–52.
Tsou P, Katayama H, Ostrin EJ, Hanash SM. The emerging role of B cells in Tumor ImmunityB cells, Cancer Immunity, and autoantibodies. Cancer Res. 2016;76(19):5597–601.
Vakil DS, Autocrine. IL-2 Signaling And The Memory CD8 + T Cell Response. 2012.
Liu Y, Lv Y, Zhang T, Huang T, Lang Y, Sheng Q et al. T cells and their products in diabetic kidney disease. Front Immunol. 2023;14.
Hu X-X, Wu Y-j, Zhang J, Wei W. T-cells interact with B cells, dendritic cells, and fibroblast-like synoviocytes as hub-like key cells in rheumatoid arthritis. Int Immunopharmacol. 2019;70:428–34.
Jianyi D, Haili G, Bo Y, Meiqin Y, Baoyou H, Haoran H, et al. Myeloid-derived suppressor cells cross-talk with B10 cells by BAFF/BAFF-R pathway to promote immunosuppression in cervical cancer. Cancer Immunol Immunother. 2023;72(1):73–85.
Banchereau J, Steinman R. Dendritic cells and the control of immunity. Nat [Internet]. 1998;392(6673):245–52.
Muthuswamy R, Urban J, Lee J-J, Reinhart TA, Bartlett D, Kalinski P. Ability of mature dendritic cells to interact with regulatory T cells is imprinted during maturation. Cancer Res. 2008;68(14):5972–8.
Lei X, Lei Y, Li J-K, Du W-X, Li R-G, Yang J, et al. Immune cells within the tumor microenvironment: Biological functions and roles in cancer immunotherapy. Cancer Lett. 2020;470:126–33.
Marigo I, Dolcetti L, Serafini P, Zanovello P, Bronte V. Tumor-induced tolerance and immune suppression by myeloid derived suppressor cells. Immunol Rev. 2008;222(1):162–79.
Nishikawa H, Sakaguchi S. Regulatory T cells in cancer immunotherapy. Curr Opin Immunol. 2014;27:1–7.
Challagundla N, Shah D, Yadav S, Agrawal-Rajput R. Saga of monokines in shaping tumour-immune microenvironment: origin to execution. Cytokine. 2022;157:155948.
Zhang B, Sun J, Yuan Y, Ji D, Sun Y, Liu Y, et al. Proximity-enabled covalent binding of IL-2 to IL-2Rα selectively activates regulatory T cells and suppresses autoimmunity. Signal Transduct Target Therapy. 2023;8(1):28.
Murdaca G, Allegra A, Paladin F, Calapai F, Musolino C, Gangemi S. Involvement of alarmins in the pathogenesis and progression of multiple myeloma. Int J Mol Sci. 2021;22(16):9039.
Jitschin R. Crosstalk of human mesenchymal stromal cells with the cellular components of the immune system. Karolinska Institutet (Sweden); 2013.
Upadhyay S, Sharma N, Gupta KB, Dhiman M. Role of immune system in tumor progression and carcinogenesis. J Cell Biochem. 2018;119(7):5028–42.
Dysthe M, Parihar R. Myeloid-derived suppressor cells in the tumor microenvironment. Tumor Microenvironment: Hematopoietic Cells–Part A. 2020:117 – 40.
Rébé C, Végran F, Berger H, Ghiringhelli F. STAT3 activation: a key factor in tumor immunoescape. Jak-stat. 2013;2(1):e23010.
Hao Z, Li R, Wang Y, Li S, Hong Z, Han Z. Landscape of myeloid-derived suppressor cell in tumor immunotherapy. Biomark Res. 2021;9(1):1–28.
Malek TR. The biology of interleukin-2. Annu Rev Immunol. 2008;26:453–79.
Ye C, Brand D, Zheng SG, Targeting. IL-2: an unexpected effect in treating immunological diseases. Signal Transduct Target therapy. 2018;3(1):2.
Lotze MT, Matory Y, Ettinghausen S, Rayner A, Sharrow S, Seipp C et al. In vivo administration of purified human interleukin 2. II. Half life, immunologic effects, and expansion of peripheral lymphoid cells in vivo with recombinant IL 2. Journal of immunology (Baltimore, Md: 1950). 1985;135(4):2865-75.
Bulgarelli J, Piccinini C, Petracci E, Pancisi E, Granato AM, de Rosa F, et al. Radiotherapy and High-Dose Interleukin-2: clinical and immunological results of a proof of Principle Study in Metastatic Melanoma and Renal Cell Carcinoma. Front Immunol. 2021;12:778459.
Buchbinder EI, Dutcher JP, Daniels GA, Curti BD, Patel SP, Holtan SG, et al. Therapy with high-dose Interleukin-2 (HD IL-2) in metastatic melanoma and renal cell carcinoma following PD1 or PDL1 inhibition. J Immunother Cancer. 2019;7(1):1–7.
Shah NR, Declouette B, Ansari-Gilani K, Alhomoud MS, Hoimes C, Ramaiya NH, et al. High-dose interleukin-2 therapy related adverse events and implications on imaging. Diagn Interventional Radiol. 2021;27(5):684.
Goff SL, Dudley ME, Citrin DE, Somerville RP, Wunderlich JR, Danforth DN, et al. Randomized, prospective evaluation comparing intensity of lymphodepletion before adoptive transfer of tumor-infiltrating lymphocytes for patients with metastatic melanoma. J Clin Oncol. 2016;34(20):2389.
Setoguchi R, Hori S, Takahashi T, Sakaguchi S. Homeostatic maintenance of natural Foxp3 + CD25 + CD4 + regulatory T cells by interleukin (IL)-2 and induction of autoimmune disease by IL-2 neutralization. J Exp Med. 2005;201(5):723–35.
Condamine T, Gabrilovich DI. Molecular mechanisms regulating myeloid-derived suppressor cell differentiation and function. Trends Immunol. 2011;32(1):19–25.
Kobayashi M, Fitz L, Ryan M, Hewick RM, Clark SC, Chan S, et al. Identification and purification of natural killer cell stimulatory factor (NKSF), a cytokine with multiple biologic effects on human lymphocytes. J Exp Med. 1989;170(3):827–45.
Anderson KG, Stromnes IM, Greenberg PD. Obstacles posed by the tumor microenvironment to T cell activity: a case for synergistic therapies. Cancer Cell. 2017;31(3):311–25.
Gattinoni L, Finkelstein SE, Klebanoff CA, Antony PA, Palmer DC, Spiess PJ, et al. Removal of homeostatic cytokine sinks by lymphodepletion enhances the efficacy of adoptively transferred tumor-specific CD8 + T cells. J Exp Med. 2005;202(7):907–12.
Klatzmann D, Abbas AK. The promise of low-dose interleukin-2 therapy for autoimmune and inflammatory diseases. Nat Rev Immunol. 2015;15(5):283–94.
Hodi FS, O’day SJ, McDermott DF, Weber RW, Sosman JA, Haanen JB, et al. Improved survival with ipilimumab in patients with metastatic melanoma. N Engl J Med. 2010;363(8):711–23.
Sulzmaier FJ, Kern N, Ahn SJ, Polovina A, Ho J, Hussain A et al. INBRX-120, a CD8α-targeted detuned IL-2 that selectively expands and activates tumoricidal effector cells for safe and durable in vivo responses. Journal for Immunotherapy of Cancer. 2023;11(1).
Ye F, Huang Z, Cheng L, Huang J, Chen S, Hua M, et al. 1104 a safe and highly potent PD-1-IL-2 fusion (AWT020) that decouples the efficacy and toxicity of IL-2 therapy. BMJ Specialist Journals; 2022.
Rosen DB, Kvarnhammar AM, Laufer B, Knappe T, Karlsson JJ, Hong E et al. TransCon IL-2 β/γ: a novel long-acting prodrug with sustained release of an IL-2Rβ/γ-selective IL-2 variant with improved pharmacokinetics and potent activation of cytotoxic immune cells for the treatment of cancer. Journal for Immunotherapy of Cancer. 2022;10(7).
Czerniecki BJ, Bertholet S, Klinman D, McArdle SE, Cuzzubbo S, Mangsbo S, et al. Cancer Vaccines: Adjuvant Potency, Importance of Age, Lifestyle, and treatments. Cancer Vaccines: Time to Think Differently!; 2021.
Emens LA, Asquith JM, Leatherman JM, Kobrin BJ, Petrik S, Laiko M, et al. Timed sequential treatment with cyclophosphamide, doxorubicin, and an allogeneic granulocyte-macrophage colony-stimulating factor–secreting breast tumor vaccine: a chemotherapy dose-ranging factorial study of safety and immune activation. J Clin Oncol. 2009;27(35):5911.
Cuzzubbo S, Mangsbo S, Nagarajan D, Habra K, Pockley AG, McArdle SE. Cancer vaccines: adjuvant potency, importance of age, lifestyle, and treatments. Front Immunol. 2021:3850.
Tang Q, Bluestone JA. The Foxp3 + regulatory T cell: a jack of all trades, master of regulation. Nat Immunol. 2008;9(3):239–44.
Nelson BH, Willerford DM. Biology of the interleukin-2 receptor. Adv Immunol. 1998;70:1–81.
Maude SL, Barrett D, Teachey DT, Grupp SA. Managing cytokine release syndrome associated with novel T cell-engaging therapies. Cancer J (Sudbury Mass). 2014;20(2):119.
Rosenberg SA, Yang JC, White DE, Steinberg SM. Durability of complete responses in patients with metastatic cancer treated with high-dose interleukin-2: identification of the antigens mediating response. Ann Surg. 1998;228(3):307.
Mocellin S, Pasquali S, Rossi CR, Nitti D. Interferon alpha adjuvant therapy in patients with high-risk melanoma: a systematic review and meta-analysis. J Natl Cancer Inst. 2010;102(7):493–501.
Waldmann TA. The shared and contrasting roles of IL2 and IL15 in the life and death of normal and neoplastic lymphocytes: implications for cancer therapy. Cancer Immunol Res. 2015;3(3):219–27.
Leonard WJ, Lin J-X. Cytokine receptor signaling pathways. J Allergy Clin Immunol. 2000;105(5):877–88.
Zhou Y, Quan G, Liu Y, Shi N, Wu Y, Zhang R et al. The application of Interleukin-2 family cytokines in tumor immunotherapy research. Front Immunol. 2023;14.
Wrangle JM, Patterson A, Johnson CB, Neitzke DJ, Mehrotra S, Denlinger CE, et al. IL-2 and beyond in cancer immunotherapy. J Interferon Cytokine Res. 2018;38(2):45–68.
Yang Y, Lundqvist A. Immunomodulatory effects of IL-2 and IL-15; implications for cancer immunotherapy. Cancers. 2020;12(12):3586.
Rubin LA, Nelson DL. The soluble interleukin-2 receptor: biology, function, and clinical application. Ann Intern Med. 1990;113(8):619–27.
Kirschke S, Ogunsulire I, Selvakumar B, Schumacher N, Sezin T, Rose-John S et al. The metalloprotease ADAM10 generates soluble interleukin-2 receptor alpha (sCD25) in vivo. J Biol Chem. 2022;298(6).
Vanmaris RM, Rijkers GT, Sarcoidosis. Vasculitis and Diffuse Lung Diseases. 2017;34(2):122.
Bien E, Balcerska A. Serum soluble interleukin 2 receptor α in human cancer of adults and children: a review. Biomarkers. 2008;13(1):1–26.
Damoiseaux J. The IL-2–IL-2 receptor pathway in health and disease: the role of the soluble IL-2 receptor. Clin Immunol. 2020;218:108515.
Minami Y, Kono T, Miyazaki T, Taniguchi T. The IL-2 receptor complex: its structure, function, and target genes. Annu Rev Immunol. 1993;11(1):245–68.
Li Y, Li X, Geng X, Zhao H. The IL-2A receptor pathway and its role in lymphocyte differentiation and function. Cytokine & Growth Factor Reviews; 2022.
Kuhn DJ, Dou QP. The role of interleukin-2 receptor alpha in cancer. Front Bioscience-Landmark. 2005;10(2):1462–74.
Bendickova K, Fric J. Roles of IL-2 in bridging adaptive and innate immunity, and as a tool for cellular immunotherapy. J Leucocyte Biology. 2020;108(1):427–37.
Gao H, Wang L, Ren J, Liu Y, Liang S, Zhang B, et al. Interleukin 2 receptor subunit beta as a novel hub gene plays a potential role in the immune microenvironment of abdominal aortic aneurysms. Gene. 2022;827:146472.
Zhou X, Xing J, Tang X, Sheng X, Chi H, Zhan W. Interleukin-2 (IL-2) interacts with IL-2 receptor beta (IL-2Rβ): its potential to enhance the proliferation of CD4 + T lymphocytes in flounder (Paralichthys olivaceus). Front Immunol. 2020;11:531785.
Zhang Z, Gothe F, Pennamen P, James JR, McDonald D, Mata CP, et al. Human interleukin-2 receptor β mutations associated with defects in immunity and peripheral tolerance. J Exp Med. 2019;216(6):1311–27.
Zhou X, Xing J, Tang X, Sheng X, Zhan W. Immunological characteristics of Interleukin-2 receptor subunit beta (IL-2Rβ) in flounder (Paralichtlys olivaceus): implication for IL-2R function. Fish Shellfish Immunol. 2019;93:641–51.
Li G, Wang Y, Cheng Y. IL2RB Is a Prognostic Biomarker Associated with Immune Infiltrates in Pan-Cancer. Journal of Oncology. 2022;2022.
Rochman Y, Spolski R, Leonard WJ. New insights into the regulation of T cells by γc family cytokines. Nat Rev Immunol. 2009;9(7):480–90.
Belaid B, Lamara Mahammed L, Mohand Oussaid A, Migaud M, Khadri Y, Casanova JL, et al. Case Report: Interleukin-2 receptor common Gamma Chain defect presented as a Hyper-IgE syndrome. Front Immunol. 2021;12:696350.
Mahdavi FS, Keramatipour M, Ansari S, Sharafian S, Karamzade A, Tavakol M. X-linked SCID with a rare mutation. Allergy Asthma & Clinical Immunology. 2021;17:1–9.
Wang X, Rickert M, Garcia KC. Structure of the Quaternary Complex of Interleukin-2 with its α, ß, and γc receptors. Science. 2005;310(5751):1159–63.
Kucukguven A, Khalil A. Matrix metalloproteinases as potential targets in the venous dilation associated with varicose veins. Curr Drug Targets. 2013;14(3):287–324.
Machowicz R, Janka G, Wiktor-Jedrzejczak W. Similar but not the same: differential diagnosis of HLH and sepsis. Crit Rev Oncol/Hematol. 2017;114:1–12.
Baughman RP, Valeyre D. Sarcoidosis: A Clinician’s Guide. Elsevier Health Sciences; 2018.
Chechlinska M, Duma A, Swierkowska K, Kaminska J, Steffen J. Sera of lung cancer patients affect the release of Th1, Th2 and monocyte-derived cytokines, and the expression of IL-2Ralpha by normal, stimulated mononuclear cells. Cell Mol Biology Lett. 2004;9(1):69–82.
Witkowska AM. On the role of sIL-2R measurements in rheumatoid arthritis and cancers. Mediat Inflamm. 2005;2005(3):121–30.
Xu X, Wang W, Sun T, Tian B, Du L, Jing J. The Predicting Role of Serum TSGF and sIL-2R for the Lymph Node Metastasis of Papillary Thyroid Carcinoma. Disease Markers. 2022;2022.
Siemiątkowska A, Bryl M, Kosicka-Noworzyń K, Tvrdoň J, Gołda-Gocka I, Barinow-Wojewódzki A, et al. Serum sCD25 protein as a predictor of lack of long-term benefits from Immunotherapy in Non-Small Cell Lung Cancer: a pilot study. Cancers. 2021;13(15):3702.
Gonda K, Horita S, Maejima Y, Takenoshita S, Shimomura K. Soluble interleukin-2 receptor as a predictive and prognostic marker for patients with familial breast cancer. Sci Prog. 2021;104(3):00368504211039590.
Pavlidis N, Nicolaides C, Bairaktari E, Kalef-Ezra J, Athanassiadis A, Seferiadis C, et al. Soluble interleukin-2 receptors in patients with advanced colorectal carcinoma. Int J Biol Mark. 1996;11(1):6–11.
Dik W, Heron M. Clinical significance of soluble interleukin-2 receptor measurement in immune-mediated diseases. Neth J Med. 2020;78(5):220–31.
Kharagjitsing H, Hendriksz T, Fouraux M, van Gelder T, van Bommel E. Clinical significance of soluble interleukin-2 receptor measurement in patients with idiopathic retroperitoneal fibrosis. Int Urol Nephrol. 2022;54(10):2663–71.
Setrerrahmane S, Xu H. Tumor-related interleukins: old validated targets for new anti-cancer drug development. Mol Cancer. 2017;16(1):1–17.
Yoshida N, Oda M, Kuroda Y, Katayama Y, Okikawa Y, Masunari T, et al. Clinical significance of sIL-2R levels in B-cell lymphomas. PLoS ONE. 2013;8(11):e78730.
Murakami S. Soluble interleukin-2 receptor in cancer. Front Bioscience-Landmark. 2004;9(5):3085–90.
Sugiura R, Satoh R, Takasaki T. ERK: a double-edged sword in cancer. ERK-dependent apoptosis as a potential therapeutic strategy for cancer. Cells. 2021;10(10):2509.
Kuhn DJ, Smith DM, Pross S, Whiteside TL, Dou QP. Overexpression of interleukin-2 receptor α in a human squamous cell carcinoma of the head and neck cell line is associated with increased proliferation, drug resistance, and transforming ability. J Cell Biochem. 2003;89(4):824–36.
Briukhovetska D, Dörr J, Endres S, Libby P, Dinarello CA, Kobold S. Interleukins in cancer: from biology to therapy. Nat Rev Cancer. 2021;21(8):481–99.
Zhang M, Mathews Griner LA, Ju W, Duveau DY, Guha R, Petrus MN et al. Selective targeting of JAK/STAT signaling is potentiated by Bcl-xL blockade in IL-2–dependent adult T-cell leukemia. Proceedings of the National Academy of Sciences. 2015;112(40):12480-5.
Hu X, Li J, Fu M, Zhao X, Wang W. The JAK/STAT signaling pathway: from bench to clinic. Signal Transduct Target therapy. 2021;6(1):402.
Rovelli F, Lissoni P, Crispino S, Barni S, Fumagalli G, Paolorossi F, et al. Increased level of soluble interleukin-2 receptor in advanced solid tumors: a preliminary study. Tumori J. 1988;74(6):633–7.
Viviani S, Camerini E, Bonfante V, Santoro A, Balzarotti M, Fornier M, et al. Soluble interleukin-2 receptors (sIL-2R) in Hodgkin’s disease: outcome and clinical implications. Br J Cancer. 1998;77(6):992–7.
Sella A, Kilbourn RG, Gray I, Finn L, Zukiwski AA, Ellerhorst J, et al. Phase I study of interleukin-2 combined with interferon-α and 5-fluorouracil in patients with metastatic renal cell cancer. Cancer Biother Radiopharm. 1994;9(2):103–11.
Atkins MB, Sparano J, Fisher RI, Weiss GR, Margolin KA, Fink KI, et al. Randomized phase II trial of high-dose interleukin-2 either alone or in combination with interferon alfa-2b in advanced renal cell carcinoma. J Clin Oncol. 1993;11(4):661–70.
Clamon G, Herndon J, Perry MC, Ozer H, Kreisman H, Maher T et al. Interleukin-2 activity in patients with extensive small-cell lung cancer: a phase II trial of Cancer and Leukemia Group B. JNCI: Journal of the National Cancer Institute. 1993;85(4):316 – 20.
Karakus U. A novel anti-interleukin-2 antibody mediates receptor-gated Interleukin-2 delivery to Activate Regulatory T cells in three species. University of Zurich; 2020.
Khoryati L, Pham MN, Sherve M, Kumari S, Cook K, Pearson J, et al. An IL-2 mutein engineered to promote expansion of regulatory T cells arrests ongoing autoimmunity in mice. Sci Immunol. 2020;5(50):eaba5264.
Yuan Y, Kolios AG, Liu Y, Zhang B, Li H, Tsokos GC et al. Therapeutic potential of interleukin-2 in autoimmune diseases. Trends Mol Med. 2022.
Besser M, Schallmach E, Oved K, Treves A, Markel G, Reiter Y, et al. Modifying interleukin-2 concentrations during culture improves function of T cells for adoptive immunotherapy. Cytotherapy. 2009;11(2):206–17.
Ghelani A, Bates D, Conner K, Wu M-Z, Lu J, Hu Y-L, et al. Defining the threshold IL-2 signal required for induction of selective Treg cell responses using engineered IL-2 muteins. Front Immunol. 2020;11:1106.
Harris F, Arroyo Berdugo Y, Tree T. IL-2-based approaches to Treg Enhancement. Clinical and Experimental Immunology; 2022.
Floros T, Tarhini AA, editors. Anticancer cytokines: biology and clinical effects of interferon-α2, interleukin (IL)-2, IL-15, IL-21, and IL-12. Seminars in oncology. Elsevier; 2015.
Moorman CD, Sohn SJ, Phee H. Emerging therapeutics for immune tolerance: tolerogenic vaccines, T cell therapy, and IL-2 therapy. Front Immunol. 2021;12:657768.
Arenas-Ramirez N, Zou C, Popp S, Zingg D, Brannetti B, Wirth E, et al. Improved cancer immunotherapy by a CD25-mimobody conferring selectivity to human interleukin-2. Sci Transl Med. 2016;8(367):367ra166–6.
Mitra S, Leonard WJ. Biology of IL-2 and its therapeutic modulation: mechanisms and strategies. J Leukoc Biol. 2018;103(4):643–55.
Arenas-Ramirez N. Immunotherapy using a Novel Agonistic Anti-IL-2 antibody combined with epigenetic modulation limits Tumor Immune escape and controls. Tumor Growth: University of Zurich; 2016.
Norell H, Poschke I, Charo J, Wei WZ, Erskine C, Piechocki MP, et al. Vaccination with a plasmid DNA encoding HER-2/neu together with low doses of GM-CSF and IL-2 in patients with metastatic breast carcinoma: a pilot clinical trial. J translational Med. 2010;8(1):1–11.
Mizui M. Natural and modified IL-2 for the treatment of cancer and autoimmune diseases. Clin Immunol. 2019;206:63–70.
Mullard A, Restoring. IL-2 to its cancer immunotherapy glory. Nat Rev Drug Discovery. 2021;20(3):163–6.
Clinical Trials for NKTR-214. Retrieved from https://clinicaltrials.gov/ct2/show/NCT03635983: United States National Library of Medicine; 2023 [.
Administration USFaD. FDA Approves Tebentafusp-tebn for the Treatment of Unresectable or Metastatic Uveal Melanoma. https://www.fda.gov/drugs/resources-information-approved-drugs/fda-approves-tebentafusp-tebn-unresectable-or-metastatic-uveal-melanoma 2022 [.
Hirai T, Ramos TL, Lin P-Y, Simonetta F, Su LL, Picton LK et al. Selective expansion of regulatory T cells using an orthogonal IL-2/IL-2 receptor system facilitates transplantation tolerance. J Clin Investig. 2021;131(8).
Overwijk WW, Tagliaferri MA, Zalevsky J. Engineering IL-2 to give new life to T cell immunotherapy. Annu Rev Med. 2021;72:281–311.
Bell CJ, Sun Y, Nowak UM, Clark J, Howlett S, Pekalski ML, et al. Sustained in vivo signaling by long-lived IL-2 induces prolonged increases of regulatory T cells. J Autoimmun. 2015;56:66–80.
Tang A, Harding F. The challenges and molecular approaches surrounding interleukin-2-based therapeutics in cancer. Cytokine: X. 2019;1(1):100001.
Lopes JE, Fisher JL, Flick HL, Wang C, Sun L, Ernstoff MS et al. ALKS 4230: a novel engineered IL-2 fusion protein with an improved cellular selectivity profile for cancer immunotherapy. J Immunother Cancer. 2020;8(1).
Ramos TL, Bolivar-Wagers S, Jin S, Thangavelu G, Simonetta F, Lin P-Y, et al. Prevention of acute GVHD using an orthogonal IL-2/IL-2Rβ system to selectively expand regulatory T cells in vivo. Blood The Journal of the American Society of Hematology. 2023;141(11):1337–52.
Sznol M, Rizvi N. Teaching an old dog new tricks: re-engineering IL-2 for immuno-oncology applications. BMJ Specialist Journals; 2023. p. e006346.
Quijano-Rubio A, Bhuiyan AM, Yang H, Leung I, Bello E, Ali LR, et al. A split, conditionally active mimetic of IL-2 reduces the toxicity of systemic cytokine therapy. Nat Biotechnol. 2023;41(4):532–40.
Codarri Deak L, Nicolini V, Hashimoto M, Karagianni M, Schwalie PC, Lauener L, et al. PD-1-cis IL-2R agonism yields better effectors from stem-like CD8 + T cells. Nature. 2022;610(7930):161–72.
Zhang Q, Hresko ME, Picton LK, Su L, Hollander MJ, Nunez-Cruz S, et al. A human orthogonal IL-2 and IL-2Rβ system enhances CAR T cell expansion and antitumor activity in a murine model of leukemia. Sci Transl Med. 2021;13(625):eabg6986.
Evans LS, Witte PR, Feldhaus AL, Nelson BH, Riddell SR, Greenberg PD, et al. Expression of chimeric granulocyte-macrophage colony-stimulating factor/interleukin 2 receptors in human cytotoxic T lymphocyte clones results in granulocyte-macrophage colony-stimulating factor-dependent growth. Hum Gene Ther. 1999;10(12):1941–51.
Dolgin E. IL-2 upgrades show promise at ASCO. Nat Biotechnol. 2022;40(7):986–8.
Silk AW, Curti B, Bryan J, Saunders T, Shih W, Kane MP et al. A phase ib study of interleukin-2 plus pembrolizumab for patients with advanced melanoma. Front Oncol. 2023;13.
Caram MV. Phase Ib/II trial of Interleukin-2 and PD-1 checkpoint inhibitor, Nivolumab in Metastatic Clear Cell Renal Cell Cancer: UMCC 2016.103.
Takai R, Funakoshi Y, Suto H, Nagatani Y, Imamura Y, Toyoda M, et al. Serum soluble interleukin-2 receptor as a potential biomarker for immune-related adverse events. Anticancer Res. 2021;41(2):1021–6.
Yan WF, Nie CF, Wu G, Zhang JC, Zhu YZ, Zhang W, et al. Soluble interleukin–2 receptor as a factor associated with angiogenesis in gastric cancer. Mol Med Rep. 2017;16(5):6916–9.
Al-Haideri M, Tondok SB, Safa SH, Rostami S, Jalil AT, Al-Gazally ME, et al. CAR-T cell combination therapy: the next revolution in cancer treatment. Cancer Cell Int. 2022;22(1):1–26.
Patricia AG, editor. CAR T cells with dual targeting of CD19 and CD22 in adult patients with recurrent or refractory B cell malignancies: a phase 1 trial. Traducciones 2019–2022; 2021.
Petrazzuolo A, Maiuri MC, Zitvogel L, Kroemer G, Kepp O. Trial Watch: combination of tyrosine kinase inhibitors (TKIs) and immunotherapy. Oncoimmunology. 2022;11(1):2077898.
Lam E, Wong M, Agarwal N, Redman B, Logan T, Flaig T, et al. Safety and efficacy of sequencing high-dose interleukin 2 (IL2) after tyrosine kinase inhibitor (TKI) therapy for metastatic renal cell carcinoma. J Clin Oncol. 2011;29(15suppl):e15079–e.
Dutcher JP, Flippot R, Fallah J, Escudier B. On the shoulders of giants: the evolution of renal cell carcinoma treatment—cytokines, targeted therapy, and immunotherapy. Am Soc Clin Oncol Educational Book. 2020;40:418–35.
Re GL, Santeufemia DA, Re FL, Bortolus R, Doretto P, Marus W, et al. Interleukin-2 chronotherapy for metastatic renal cell carcinoma: results of a phase I-II study. Cytokine. 2020;128:154984.
Clark JI, Wong MK, Kaufman HL, Daniels GA, Morse MA, McDermott DF, et al. Impact of sequencing targeted therapies with high-dose interleukin-2 immunotherapy: an analysis of outcome and survival of patients with metastatic renal cell carcinoma from an on-going observational IL-2 clinical trial: PROCLAIMSM. Clin Genitourin Cancer. 2017;15(1):31–41. e4.
Rosenzwajg M, Lorenzon R, Cacoub P, Pham HP, Pitoiset F, El Soufi K, et al. Immunological and clinical effects of low-dose interleukin-2 across 11 autoimmune diseases in a single, open clinical trial. Ann Rheum Dis. 2019;78(2):209–17.
Trzonkowski P, Bieniaszewska M, Juścińska J, Dobyszuk A, Krzystyniak A, Marek N, et al. First-in-man clinical results of the treatment of patients with graft versus host disease with human ex vivo expanded CD4 + CD25 + CD127 – T regulatory cells. Clin Immunol. 2009;133(1):22–6.
Trotta E, Bessette PH, Silveria SL, Ely LK, Jude KM, Le DT, et al. A human anti-IL-2 antibody that potentiates regulatory T cells by a structure-based mechanism. Nat Med. 2018;24(7):1005–14.
Wang CJ, Petersone L, Edner NM, Heuts F, Ovcinnikovs V, Ntavli E, et al. Costimulation blockade in combination with IL-2 permits regulatory T cell sparing immunomodulation that inhibits autoimmunity. Nat Commun. 2022;13(1):6757.
Moon D, Tae N, Park Y, Lee S-W, Kim DH. Development of bispecific antibody for cancer immunotherapy: focus on T cell engaging antibody. Immune Netw. 2022;22(1).
Labrijn AF, Janmaat ML, Reichert JM, Parren PW. Bispecific antibodies: a mechanistic review of the pipeline. Nat Rev Drug Discovery. 2019;18(8):585–608.
Huehls AM, Coupet TA, Sentman CL. Bispecific T-cell engagers for cancer immunotherapy. Immunol Cell Biol. 2015;93(3):290–6.
Seimetz D, Lindhofer H, Bokemeyer C. Development and approval of the trifunctional antibody catumaxomab (anti-EpCAM× anti-CD3) as a targeted cancer immunotherapy. Cancer Treat Rev. 2010;36(6):458–67.
You G, Won J, Lee Y, Moon D, Park Y, Lee SH, et al. Bispecific antibodies: a smart arsenal for cancer immunotherapies. Vaccines. 2021;9(7):724.
Zhang B, Li W, Fan D, Tian W, Zhou J, Ji Z, et al. Advances in the study of CD47-based bispecific antibody in cancer immunotherapy. Immunology. 2022;167(1):15–27.
Wei J, Yang Y, Wang G, Liu M. Current landscape and future directions of bispecific antibodies in cancer immunotherapy. Front Immunol. 2022;13:6573.
Kontermann RE, Brinkmann U. Bispecific antibodies. Drug Discovery Today. 2015;20(7):838–47.
Chen L, Flies DB. Molecular mechanisms of T cell co-stimulation and co-inhibition. Nat Rev Immunol. 2013;13(4):227–42.
Li J, Jie H-B, Lei Y, Gildener-Leapman N, Trivedi S, Green T, et al. PD-1/SHP-2 inhibits Tc1/Th1 phenotypic responses and the activation of T cells in the Tumor MicroenvironmentPD-1/SHP-2 inhibits type I response and TCR activation in TME. Cancer Res. 2015;75(3):508–18.
Choi BD, Yu X, Castano AP, Darr H, Henderson DB, Bouffard AA, et al. CRISPR-Cas9 disruption of PD-1 enhances activity of universal EGFRvIII CAR T cells in a preclinical model of human glioblastoma. J Immunother Cancer. 2019;7(1):1–8.
van der Horst HJ, Nijhof IS, Mutis T, Chamuleau ME. Fc-engineered antibodies with enhanced Fc-effector function for the treatment of B-cell malignancies. Cancers. 2020;12(10):3041.
Liu R, Oldham RJ, Teal E, Beers SA, Cragg MS. Fc-engineering for modulated effector functions—improving antibodies for cancer treatment. Antibodies. 2020;9(4):64.
Bruhns P. Properties of mouse and human IgG receptors and their contribution to disease models. Blood The Journal of the American Society of Hematology. 2012;119(24):5640–9.
Li F, Ravetch JV. Inhibitory Fcγ receptor engagement drives adjuvant and anti-tumor activities of agonistic CD40 antibodies. Science. 2011;333(6045):1030–4.
Yang Z, Hou Y, Grande G, Wang C, Shi Y, Zak J et al. Enhancing the anti-tumor efficacy of bispecific T cell engagers via cell surface glycocalyx editing. bioRxiv. 2022:2022.05. 22.492978.
Wilkinson I, Anderson S, Fry J, Julien LA, Neville D, Qureshi O, et al. Fc-engineered antibodies with immune effector functions completely abolished. PLoS ONE. 2021;16(12):e0260954.
López-Cantillo G, Urueña C, Camacho BA, Ramírez-Segura C. CAR-T Cell Performance: How to Improve Their Persistence? Frontiers in Immunology. 2022:1757.
Wagner DL, Fritsche E, Pulsipher MA, Ahmed N, Hamieh M, Hegde M, et al. Immunogenicity of CAR T cells in cancer therapy. Nat Reviews Clin Oncol. 2021;18(6):379–93.
Newick K, O’Brien S, Moon E, Albelda SM. CAR T cell therapy for solid tumors. Annu Rev Med. 2017;68:139–52.
Institute NC. CAR T Cells: Engineering Patients’ Immune Cells to Treat Their Cancers. 2022.
Marofi F, Motavalli R, Safonov VA, Thangavelu L, Yumashev AV, Alexander M, et al. CAR T cells in solid tumors: challenges and opportunities. Stem Cell Res Ther. 2021;12(1):1–16.
Agliardi G, Liuzzi AR, Hotblack A, De Feo D, Núñez N, Stowe CL, et al. Intratumoral IL-12 delivery empowers CAR-T cell immunotherapy in a pre-clinical model of glioblastoma. Nat Commun. 2021;12(1):444.
Aspuria P-J, Vivona S, Bauer M, Semana M, Ratti N, McCauley S, et al. An orthogonal IL-2 and IL-2Rβ system drives persistence and activation of CAR T cells and clearance of bulky lymphoma. Sci Transl Med. 2021;13(625):eabg7565.
Sun Z, Ren Z, Yang K, Liu Z, Cao S, Deng S, et al. A next-generation tumor-targeting IL-2 preferentially promotes tumor-infiltrating CD8 + T-cell response and effective tumor control. Nat Commun. 2019;10(1):3874.
Kim J-H, Lee K-J, Lee S-W. Cancer immunotherapy with T-cell targeting cytokines: IL-2 and IL-7. BMB Rep. 2021;54(1):21.
Rafiq S, Hackett CS, Brentjens RJ. Engineering strategies to overcome the current roadblocks in CAR T cell therapy. Nat reviews Clin Oncol. 2020;17(3):147–67.
Sterner RC, Sterner RM. CAR-T cell therapy: current limitations and potential strategies. Blood cancer journal. 2021;11(4):69.
Stat ES. Extraordinary Superkines purge tumors and their immunosuppressive ecosystem.
Ren J, Chu AE, Jude KM, Picton LK, Kare AJ, Su L et al. Interleukin-2 superkines by computational design. Proceedings of the National Academy of Sciences. 2022;119(12):e2117401119.
Zheng X, Wu Y, Bi J, Huang Y, Cheng Y, Li Y, et al. The use of supercytokines, immunocytokines, engager cytokines, and other synthetic cytokines in immunotherapy. Cell Mol Immunol. 2022;19(2):192–209.
Raeber ME, Sahin D, Boyman O. Interleukin-2–based therapies in cancer. Sci Transl Med. 2022;14(670):eabo5409.
Takeshita T, Asao H, Ohtani K, Ishii N, Kumaki S, Tanaka N, et al. Cloning of the γ chain of the human IL-2 receptor. Science. 1992;257(5068):379–82.
Liao W, Lin J-X, Leonard WJ. IL-2 family cytokines: new insights into the complex roles of IL-2 as a broad regulator of T helper cell differentiation. Curr Opin Immunol. 2011;23(5):598–604.
Markley JC, Sadelain M. IL-7 and IL-21 are superior to IL-2 and IL-15 in promoting human T cell–mediated rejection of systemic lymphoma in immunodeficient mice. Blood The Journal of the American Society of Hematology. 2010;115(17):3508–19.
Abbas AK, Trotta E, Simeonov R, Marson D, Bluestone A. Revisiting IL-2: Biology and therapeutic prospects. Sci Immunol. 2018;3(25):eaat1482.
Gupta S, Agarwal N. Cancer Immunotherapy with Interleukin-2—Current Status and Future Developments. Oncol Hematol Rev (US). 2016;12(02):82.
Gillies SD, Stein PA, Lo K-M. DNA encoding IL-2 fusion proteins with modulated selectivity. Google Patents; 2011.
Kim J, Kang S, Kim KW, Heo M-G, Park D-I, Lee J-H, et al. Nanoparticle delivery of recombinant IL-2 (BALLkine-2) achieves durable tumor control with less systemic adverse effects in cancer immunotherapy. Biomaterials. 2022;280:121257.
Wang X-S, Zheng Z-S, Zheng M-F, Wang D, Zhang H-L, Zhang Z-Q et al. IL-2-loaded polypeptide nanoparticles for enhanced anti-cancer immunotherapy. Chin J Polym Sci. 2023:1–10.
Panelli MC, White R, Foster M, Martin B, Wang E, Smith K, et al. Forecasting the cytokine storm following systemic interleukin (IL)-2 administration. J translational Med. 2004;2:1–14.
Skrombolas D, Frelinger JG. Challenges and developing solutions for increasing the benefits of IL-2 treatment in tumor therapy. Expert Rev Clin Immunol. 2014;10(2):207–17.
Carmenate T, Ortíz Y, Enamorado M, García-Martínez K, Avellanet J, Moreno E, et al. Blocking IL-2 signal in vivo with an IL-2 antagonist reduces tumor growth through the control of regulatory T cells. J Immunol. 2018;200(10):3475–84.
Liu DV, Maier LM, Hafler DA, Wittrup KD. Engineered interleukin-2 antagonists for the inhibition of regulatory T cells. Journal of immunotherapy (Hagerstown, Md: 1997). 2009;32(9):887.
Shi H, Wang W, Yin J, Ouyang Y, Pang L, Feng Y, et al. The inhibition of IL-2/IL-2R gives rise to CD8 + T cell and lymphocyte decrease through JAK1-STAT5 in critical patients with COVID-19 pneumonia. Cell Death Dis. 2020;11(6):429.
Uchibori R, Teruya T, Ninomiya S, Ido H, Tsukahara T, Ohmine K, et al. 225. Activation signals from CD19-CAR Permit NFAT-Controlled Inducible expression of transgenes in PBMCs. Mol Ther. 2015;23:89.
Goudy KS, Johnson MC, Garland A, Li C, Samulski RJ, Wang B, et al. Inducible adeno-associated virus-mediated IL-2 gene therapy prevents autoimmune diabetes. J Immunol. 2011;186(6):3779–86.
Xu Z, Zeng S, Gong Z, Yan Y. Exosome-based immunotherapy: a promising approach for cancer treatment. Mol Cancer. 2020;19:1–16.
Zhang H, Wang S, Sun M, Cui Y, Xing J, Teng L et al. Exosomes as smart drug delivery vehicles for cancer immunotherapy. Front Immunol. 2022;13.
Jung D, Shin S, Kang SM, Jung I, Ryu S, Noh S, et al. Reprogramming of T cell-derived small extracellular vesicles using IL2 surface engineering induces potent anti‐cancer effects through miRNA delivery. J Extracell Vesicles. 2022;11(12):12287.
Zhou Y, Zhang Y, Gong H, Luo S, Cui Y. The role of exosomes and their applications in cancer. Int J Mol Sci. 2021;22(22):12204.
Park S. Exosome Encapsulation of Curcumin Blocks TNF-α and IL-2 Dependent Cytokine Storm. 2022.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Key R&D Program of China (2020YFC2006400, 2020AAA0109500), the National Natural Science Foundation of China (82122053 to GY, 82188102 to JH), the CAMS Initiative for Innovative Medicine (2021-I2M-1-067 to GY), the Non-profit Central Research Institute Fund of Chinese Academy of Medical Sciences (2021-RC310-020 to GY), and the Key Area Research and Development Program of Guangdong Province (2021B0101420005 to GY).
Author information
Authors and Affiliations
Contributions
SM, FT, YH, GY, and HJ conceptualized and designed the study and reviewed and revised the manuscript. SM and FT, were involved in data acquisition and analysis. GY and HJ supervised the study. All of the authors have read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interest
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Shan Muhammad and Tao Fan contributed equally to this work and share first authorship.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Table.1
: A Comprehensive overview of IL-2 and IL-2R based novel approaches in cancer treatment
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Muhammad, S., Fan, T., Hai, Y. et al. Reigniting hope in cancer treatment: the promise and pitfalls of IL-2 and IL-2R targeting strategies. Mol Cancer 22, 121 (2023). https://doi.org/10.1186/s12943-023-01826-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12943-023-01826-7
Keywords
- Interleukin-2 (IL-2)
- IL-2 receptor (IL-2R)
- Tumor microenvironment
- IL-2-based immunotherapy
- Regulatory T cells
- Cancer biomarkers
- Engineered IL-2 variants
- Combination cancer therapy
- Checkpoint inhibitors
- Tumor immune response
- Dose optimization of IL-2
- Cancer prognosis and IL-2
- Immune activation and tolerance
- Tumorigenesis
- Personalized cancer treatment
- Cancer immunosurveillance