
Abstract
Cancer-associated fibroblasts (CAFs) are a diverse stromal cell population within the tumour microenvironment, where they play fundamental roles in cancer progression and patient prognosis. Multiple lines of evidence have identified that CAFs are critically involved in shaping the structure and function of the tumour microenvironment with numerous functions in regulating tumour behaviours, such as metastasis, invasion, and epithelial-mesenchymal transition (EMT). CAFs can interact extensively with cancer cells by producing extracellular vesicles (EVs), multiple secreted factors, and metabolites. Notably, CAF-derived EVs have been identified as critical mediators of cancer therapy resistance, and constitute novel therapy targets and biomarkers in cancer management. This review aimed to summarize the biological roles and detailed molecular mechanisms of CAF-derived EVs in mediating cancer resistance to chemotherapy, targeted therapy agents, radiotherapy, and immunotherapy. We also discussed the therapeutic potential of CAF-derived EVs as novel targets and clinical biomarkers in cancer clinical management, thereby providing a novel therapeutic strategy for enhancing cancer therapy efficacy and improving patient prognosis.
Similar content being viewed by others

Introduction
Cancer continues to be a serious public health challenge affecting people worldwide [1, 2]. Despite the substantial investment in cancer research and the extensive array of anti-cancer treatments available, therapy resistance remains the most serious factor impeding therapeutic efficacy [3, 4]. The intricate panorama of cancer therapy resistance entails multifaceted mechanisms such as altered drug uptake and efflux, inhibition of apoptosis signaling pathways, activation of cancer stem cells (CSCs), enhanced DNA repair capacities, disrupted cell cycle regulation, and physical obstacles impairing drug penetration and efficacy [5,6,7]. Nowadays, multiple ongoing studies have explored new drugs and combinations to address therapy resistance. For instance, bufalin, a compound found in secretions from the glands of toads can effectively inhibit cancer progression and reverse various drug resistance mechanisms, supporting combining bufalin with standard therapies as a promising therapeutic strategy [8,9,10,11]. However, the complex molecular mechanisms behind cancer therapy resistance remain largely elusive. Notably, complex biological interactions between the tumour and tumour microenvironment have been confirmed as an essential factor that leads to cancer treatment failure. Multiple non-tumour cell types within tumour microenvironment interact closely with tumour cells and either positively or negatively mediate cancer therapy resistance [12, 13].
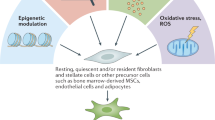
Cancer-associated fibroblasts (CAFs) are a major component of the stroma and are critically involved in the maintenance and reshaping of the extracellular matrix (ECM) [14, 15]. CAFs may originate from multiple cellular types, including quiescent tissue-resident fibroblasts, pancreatic or hepatic stellate cells, bone marrow-derived mesenchymal stem cells, endothelial cells, pericytes, and adipocytes [14, 16]. Multiple mechanisms have been confirmed to induce CAFs activation, including cancer cell-CAFs interacting signaling, ECM remodeling, oxidative stress, signals from other cells within the tumour microenvironment, cancer therapies, numerous tumor-inhibiting/promoting functions. Mechanistically, CAFs can regulate multiple tumour behaviours by secreting various cytokines, exosomes, and metabolites or by remodeling the structure and functions of the surrounding ECM [17, 18]. For instance, CAFs can produce TGFβ, fibroblast growth factor 5 (FGF5), and hepatocyte growth factor (HGF) to facilitate the invasion, proliferation, and therapy resistance of cancer cells [19]. On the contrary, CAFs can exert tumour-inhibiting functions by enhancing anticancer immunity, secreting pro-inflammatory cytokines, and producing certain ECM components as barriers to inhibit tumour cell invasion and dissemination [18] (Fig. 1).

Overview of CAFs. Potential cellular origins of CAFs include quiescent stellate cells, tissue-resident fibroblasts, mesenchymal stem cells (MSCs), endothelial cells, pericytes, and other cell types. CAFs can be activated by various mechanisms, including ECM remodeling, inflammation, oxidative stress and signalling interaction with cancer cells or other cells within tumour microenvironment. Activated CAFs can exert either tumour-restraining or tumour-promoting functions by secreting cytokines, exosomes, remodeling ECM and altering metabolism
Extracellular vesicles (EVs) are heterogeneous cell-derived membrane structures composed of exosomes and microvesicles that originate from the endosomal system or are shed from the plasma membrane, respectively [20, 21]. EVs harbor a rich cargo of molecules, including nucleic acids like DNA, mRNAs, and noncoding RNAs (ncRNAs), as well as lipids and diverse proteins. Functionally, EVs serve as vital mediators of intercellular communication, shuttling a plethora of molecules that regulate cellular processes in both normal and pathological contexts [22, 23]. Notably, multiple studies have highlighted the pivotal role of CAF-derived EVs in shaping the tumor microenvironment and influencing the development of various human cancers [24, 25]. For instance, CAF-derived EV-packaged PIAT can promote neural remodeling by enhancing the binding of YBX1 and PNI-associated mRNAs in a m5C-dependent manner in pancreatic cancer [26]. Intriguingly, CAF-derived EVs are implicated in mediating resistance to cancer therapies, including chemotherapy, targeted therapy, radiotherapy, and immunotherapy, indicating that CAFs and CAF-derived EVs can constitute a novel therapeutic target.
This review explores the crucial roles played by CAF-derived EVs in mediating resistance to cancer therapies and delves into the intricate mechanisms through which CAF-derived EVs bolster or undermine anticancer therapy resistance. Additionally, we discussed the prognostic value and therapeutic potential of CAF-derived EVs in cancer management.
CAF-derived EVs in cancer therapy resistance
Therapeutic resistance is a significant obstacle in enhancing cancer therapy efficacy and improving patient prognosis. Although multiple studies have proposed a conceptual framework of therapy resistance mechanisms, and novel therapeutic agents and combinational therapies have been developed to conquer the development of acquired therapy resistance, the complex biological mechanisms underlying cancer therapy resistance remain largely elusive. Hence, elucidating the detailed mechanisms of cancer therapy resistance and identifying novel therapy resistance-related biomarkers are critical for advancing personalized precision medicine and improving patient prognosis. Notably, ectopic alterations in CAFs and CAF-derived EVs have been found in the tumour microenvironment in various therapy-resistant cancers, indicating the role of CAF-derived EVs in cancer therapy resistance. The involvement of CAF-derived EVs in cancer resistance to chemotherapy, targeted therapy, radiotherapy and immunotherapy is summarized below (Table 1).
CAF-derived EVs in cancer chemotherapy resistance
As the first-line mode of anti-tumour treatment in multiple malignancies, multiple chemotherapeutic drugs with different properties and targets have exhibited prominent effectiveness. Nonetheless, both the inherent and acquired chemotherapy resistance take primary responsibility for treatment failure, adverse prognosis, and high recurrence rate. Multiple studies have demonstrated that CAF-derived EVs consisting of functional non-coding RNAs and proteins are critically involved in mediating cancer chemotherapy resistance through complex mechanisms and signaling pathways [27,28,29,30] (Fig. 2).

CAF-derived EVs in cancer chemotherapy resistance. The detailed biological mechanisms of CAF-derived EVs in mediating cancer chemoresistance are summarized. CAF-derived EVs can deliver miRNAs, lncRNAs, circRNAs and proteins to mediate cancer chemotherapy resistance by regulating cell cycle, DNA repair, oxidative stress, EMT, apoptosis, autophagy, cancer cell stemness, angiogenesis, immune cells infiltration and various signalling pathways including PI3K/AKT, Wnt/β-catenin, and PTEN/mTOR pathways
miRNAs
MicroRNAs (miRNAs) are about 21–23 nucleotides in length and function by binding to the 3’ untranslated region of target mRNA, leading to mRNA degradation or translational repression [31]. CAFs can deliver diverse miRNAs to targeted cancer cells through EVs, thus regulating cancer therapy resistance. For instance, CAF-derived exosomal miR-130a has been confirmed to promote cisplatin (DDP) resistance in NSCLC, and the packing and secretion of exosomal miR-130a were regulated by the PUM2 in CAFs [32]. Similarly, miR-20a was abnormally upregulated in NSCLC patient tissue samples and CAF-derived exosomes from NSCLC patients compared with normal tissues and NAFs from healthy paracancerous lung tissues, respectively. Functionally, CAF-derived exosomal miR-20a can promote tumour progression and DDP resistance by inhibiting PTEN and activating PI3K/AKT pathway in vivo and in vitro [33]. Monocytic myeloid-derived suppressor cells (M-MDSCs) have been found to correlate with DDP resistance in patients with ESCC, and CAFs can facilitate the differentiation of monocytes into M-MDSCs phenotypically and functionally in vitro. Mechanically, CAFs secreted IL-6 and exosomal miR-21 to promote M-MDSCs formation by stimulating the STAT3 signaling pathway, and CAFs-induced M-MDSCs promoted DDP resistance of ESCC cells [34]. In colon cancer, miR-24-3p transferred from CAFs to cancer cells through exosomes downregulated the expression of CDX2 and HEPN, inducing the methotrexate (MTX) resistance in both in vivo and in vitro experiments [35]. Taxanes are the first-line anti-tumor drugs for many malignant tumors via inhibiting microtubules, inducing inflammatory response and apoptosis, etc [36]. Both in vivo and in vitro experiments have revealed that CAF-derived exosomal miR-423-5p targeted 3’-UTR of GREM2 to stimulate the TGF-β signaling pathway, thereby resulting in the therapy resistance to taxane in prostate cancer [37]. Sun et.al have reported that miR-296-3p, which was upregulated in activated CAFs‐derived EVs from ovarian cancer, can promote ovarian cancer progression and paclitaxel resistance by regulating PTEN/AKT and SOCS6/STAT3 signaling pathways [38]. CAFs‐derived exosomal miR-21 can also induce ovarian cancer paclitaxel resistance, migration, and invasion by directly targeting apoptosis protease activating fator1(APAF1) [39].
Ferroptosis is a novel form of regulated cell death that involves iron-dependent lipid-ROS accumulation [40]. Ferroptosis has been identified as an important regulator of cancer therapy resistance, and inducing ferroptosis is a novel therapeutic strategy to reverse drug resistance [41]. Zhang et al. have isolated serum exosomes from gastric cancer patients and normal controls by ultra-centrifugation, and found that the expression level of miR-552 was abnormally elevated in both tumour tissues and serum exosomes of gastric cancer patients. Further investigation has reported that DDP and paclitaxel treatment can induce CAFs to secret exosomal miR-552 by activating the USP7/hnRNPA1 axis in gastric cancer. Meanwhile, CAF-derived exosomal miR-552 can impair chemosensitivity by inhibiting ferroptosis by suppressing ALOX15 [42]. Similarly, Qi et al. have obtained normal fibroblasts and CAFs from normal paracancerous pancreatic tissues and PDAC tumors respectively, and isolated normal fibroblast- and CAF-derived exosomes by ultracentrifugation. CAF-derived exosomes can inhibit ferroptosis in pancreatic cancer cells after gemcitabine (GEM) treatment in a non-contact co-culture system in vitro. Besides, miR-3173-5p was upregulated in co-induced CAF-derived exosomes and tumor tissues after GEM treatment. The mechanical investigation suggested that CAF-derived exosomal miR-3173-5p facilitated GEM resistance and suppressed ferroptosis by directly targeting ACSL4 in vivo and in vitro [43]. In prostate cancer, CAF-derived exosomal miR-432-5p reduces lipid peroxidation stress and inhibits ferroptosis by targeting CHAC1, thereby conferring docetaxel (DTX) resistance [44].
Evasion of apoptosis has been regarded as a major cause of cancer chemoresistance [45]. In esophageal squamous cell carcinoma (ESCC), CAF-derived exosomes decreased the chemotherapy sensitivity of DDP and inhibited ESCC cell apoptosis via the RIG-I/IFN-β signaling in vivo and in vitro [46]. Pancreatic fibroblasts have exhibited innately chemoresistant to GEM, and co-culturing PDAC cells with CAFs improved PDAC proliferation capability and reduced GEM-induced cell apoptosis by establishing a three-dimensional organoid-fibroblast co-culture system [47]. Simultaneously, miR-106b was upregulated in GEM-treated CAFs and CAF-derived exosomes, and exosomal miR-106b possessed the potential to promote acquired resistance to GEM via binding to Tumor protein p53-inducible nuclear protein 1 (TP53INP1) [29]. Korc et al. have isolated GEM-treated CAF-derived exosomes by ExoQuick-TC™ and microRNA-Seq was conducted to select differentially expressed miRNAs. The abnormal upregulation of a total of five miRNAs (miR-21, miR-181a, miR-221, miR-222, and miR-92a) was detected in GEM-treated CAF-derived exosomes, and these exosomal miRNAs may confer GEM resistance by directly targeting PTEN [48].
Epithelial-mesenchymal transition (EMT) plays multifaceted roles in cancer progression, metastasis, and therapy resistance, and targeting the EMT process and EMT transcription factors, e.g. ZEB, TWIST and SNAIL has become promising approaches to reverse therapy resistance [49]. GEM treatment can induce CAFs to secret more miR-146a and the specific promoter binding transcription factor of miR-146a, Snail, subsequently inducing EMT and chemoresistance to GEM in vitro [50]. CAF-derived exosomal miR-148b-3p has been demonstrated to induce the chemoresistance to PTX and DOX through inducing EMT by directly targeting PTEN and the Wnt/β-catenin pathway in bladder cancer, whereas the downregulation of miR-148b-3p and expression of PTEN could reverse the chemoresistance via the Wnt/β-catenin pathway [51]. 5-FU/L-OHP has a prominent chemotherapy effect on colorectal cancer (CRC). Hu et al. have isolated normal fibroblast- and CAF-derived exosomes by ultracentrifugation, and found that CAF-derived exosomes can facilitate invasion, metastasis, and chemotherapy resistance in CRC. miRNA microarray assay revealed that miR-92a-3p was significantly upregulated in CAFs and CAF-derived exosomes, and exosomal miR-92a-3p secreted by CAFs has been found to promote EMT, cell stemness, 5-FU/L-OHP resistance, and suppress mitochondrial apoptosis of CRC cells in vivo and in vitro. Mechanically, exosomal miR-92a-3p directly activated the Wnt/β-catenin signaling pathway and inhibited the expression of FBXW7 and MOAP [52].
Oxidative stress is an adverse condition for cancer cells survival, whereas the hypoxic microenvironment is one of the key factors that promote cancer progression and facilitate therapy resistance [53]. In pancreatic cancer, hypoxia-induced CAFs can secret miR-21 abundant extracellular vesicles to promote pancreatic cancer cell stemness and GEM resistance [54]. Mechanically, the hypoxic microenvironment induced miR-21 secretion through HIF-1α activation, and miR-21 conferred resistance to GEM by activating the RAS/ATK/ERK axis [54]. Aerobic glycolysis is a typical phenotype of cancer cells, and glycolysis is also a determinant of cancer chemoresistance [55, 56]. In malignant lymphoma, CAF-derived exosomes can support the survival of lymphoma cells by promoting glycolysis, and inhibition of exosome secretion by RAB27B suppression inhibits the survival of lymphoma cells. Besides, CAF-derived exosomal miR-4717-5p can induce anti-pyrimidine drug resistance by directly targeting ENT2 in lymphoma [57]. Cancer stem cells have been identified as a critical factor for tumor growth, chemoresistance, and recurrence, and tackling stemness-associated chemoresistance may be an effective therapy approach [58]. In urothelial bladder cancer (UBC), CAF-derived exosomal miR-146a-5p can facilitate cancer stem cell niche formation, UBC cell stemness, and resistance to GEM and DDP by directly targeting the 3’UTR of mRNAs of ARID1A and PRKAA2. Mechanically, the downregulation of ARID1A can suppress the expression of SOCS1 and activate the STAT3 signaling pathway, and downregulated PRKAA2 can activate the mTOR signaling [59]. In vivo study has demonstrated that CAF-derived exosomal miR-34c-5p reduced the DDP resistance and stemness of laryngeal squamous cell carcinoma (LSCC) [60].
LncRNAs
LncRNAs are ncRNAs that are longer than 200 nt without protein-coding potential [61]. Multiple studies have indicated that CAF-derived exosomal lncRNAs can interact with multiple miRNAs in cancer cells to mediate chemoresistance. For instance, exosomal LINC00355 secreted by CAFs promoted DDP resistance by sponging miR-34b-5p to upregulate ABCB1 in bladder cancer [62]. Intriguingly, the LINC00355/miR-34b-5p axis was also involved in facilitating EMT and the chemoresistance to DDP, OXA, and 5-FU in CRC [63]. Correspondingly in vulvar squamous cell carcinoma (VSCC) cells, exosomal lncRNA UCAI from CAFs can induce DDP resistance via the miR-103a-WWE1 axis [64]. Exosomal lncMEG3 was secreted and transferred from CAFs to SCLC cells and induced DDP chemoresistance by targeting the miR-15a-5p/CCNE1 axis [65]. Simultaneously, lncRNAs can alter the sensitivity of cancer cells to chemotherapeutic drugs depending on the no-miRNA binding pathways. Beclin1 was a crucial factor during OXA-mediated autophagic cell death in CRC cells. CAF-derived exosomal lncRNA FAL1 has been found to act as the scaffold that promoted the interaction between Beclin1 and TRIM3 and induced the TRIM3-dependent Beclin1 polyubiquitination, resulting in inhibition of autophagy and chemoresistance to OXA [66]. LncRNA CCAL in CAF-derived exosomes has been confirmed to confer OXA resistance by interacting directly with mRNA stabilizing protein HuR (human antigen R) to increase β-catenin mRNA and protein expression [67]. Downregulation of DACT3-AS1 was significantly with worse prognosis of GC patients. CAF-derived exosomal DACT3-AS1 elevated the oxidative stress level through miR-181a-5p/SIRT1-mediated ferroptosis, enhance the sensitivity to OXA of GC cells [68].
CircRNAs
In addition to miRNAs and lncRNAs, CAFs-secreted circular RNAs (circRNAs) also take a novel role in the development of both inherent and acquired chemoresistance. CircRNAs are a type of non-coding RNA that forms a covalently closed continuous loop structure [69]. For instance, circZFR was highly expressed in DDP-resistant HCC cell lines, CAFs, and CAF-derived exosomes. In vitro and in vivo studies further indicated that CAFs-secreted exosomal circZFR enhanced the DDP resistance of HCC cells by inhibiting the STAT3/NF-κB pathway [70]. As a third-generation platinum drug, OXA plays a prominent role in cancer therapy. CircN4BP2L2 secreted by CAFs could positively bind to EIF4A3 to stimulate the PI3K/AKT/mTOR axis, correspondingly inducing the resistance to OXA in CRC [71]. In PDAC, CAF-derived exosomal circFARP1 also contributed to PDAC cell stemness and GEM resistance via the LIF/STAT3 axis [72]. Researchers have isolated and purified EVs from the plasma of PDAC patients by EV isolation kit, and detected that circBIRC6 was upregulated in plasma EVs and primary tumor tissues of oxaliplatin-resistant patients. Mechanically, circBIRC6 from CAF-derived EVs can directly bound to XRCC4, which encoded DNA ligase IV and elevated the crosstalk between XRCC4 and Small Ubiquitin Like Modifier 1 (SUMO1) in PDAC. Consequently, non-homologous end joining (NHEJ) dependent DNA repair was activated, resulting in the resistance to oxaliplatin [73].
Proteins
Proteins are also an important part in CAFs-secreted EVs, and multiple CAFs-secreted exosomal proteins also serve a pivotal role in cancer progression and therapy resistance. For instance, FOSL1 was enriched in CAFs-secreted exosomes and was transmitted to CRC cells, thus conferring resistance to OXA of CRC cells by transcriptionally activating integrin β4 (ITGB4) [74]. Vascular Endothelial Growth Factor A (VEGFA) can induce the proliferation and migration of vascular endothelia cell and is involved in tumor angiogenesis [75]. Intriguingly, VEGFA was elevated in exosomes derived by CAFs isolated from DDP-resistant CRC tissues. Functionally, CRC cells exhibited enhanced viability and DDP resistance after CAF-derived exosomal VEGFA treatment [76] (Table 1).
CAF-derived EVs in cancer targeted therapy resistance
Targeted therapy involves corresponding therapeutic drugs for targeting and inhibiting driver mutations that are characteristic of certain types of cancer [77, 78]. Multiple targeted therapy drugs have been applied in the treatment of diverse cancers, such as claudiximab for gastric cancer, TKIs for lung cancer and colorectal cancer, and trastuzumab targeting HER-2 for breast cancer [79]. However, the greatest obstacle to targeted cancer therapy is the inevitable emergence of drug resistance, which contributes to therapeutic failure and ultimate patient demise in genotype-matched precision medicine [80]. Multiple studies have indicated that EMT is connected with cancer progression and targeted therapy resistance [81]. It has been identified that CAF-derived EVs were involved in EMT induction through several biological processes including ECM remodeling, metabolic reprogramming, and oncogenic pathways activating [82]. In HCC, CAF-derived protein SPP1 was able to induce TKIs (sorafenib and lenvatinib) resistance. Mechanically, SPP1 induced EMT by activating the phosphorylation of the RAF/MAPK axis and PI3K/AKT/mTOR axis through the PKCα signalling pathway [83]. Similarly, Gremlin-1 from CAF-derived exosomes was proved to have an active role in regulating the EMT process and reducing the sensitivity of HCC cells to sorafenib via Wnt/β‐catenin and BMP signaling pathways. Clinically, patients with HCC exhibited a higher exosomal gremlin-1 level in serum, and serum gremlin-1 level could predict the sorafenib response of HCC patients [84]. In addition, CAFs-secreted CXCL12 conferred sorafenib resistance in HCC cells by upregulating FOLR1 expression in the mRNA level, and the CXCL12/CXCR4/FOLR1 was involved in sorafenib resistance of HCC [85]. Moreover, the in vitro experiment has demonstrated that miR-1228-3p carried by CAF-derived exosomes conferred sorafenib resistance in HCC9724 cells by modulating the PLAC8/PI3K/AKT axis [86]. Gao et al. have identified that CD63 + CAFs, a new CAF subset, can promote ERα downregulation and tamoxifen resistance in breast cancer by single‐cell sequencing. Mechanically, CD63 + CAFs can secrete exosomal miR-22 to induce tamoxifen resistance by directly targeting ERα and PTEN. Furthermore, the packaging of miR-22 into CD63 + CAF-derived exosomes was regulated by SFRS1, and CD63 stimulated STAT3 to maintain the phenotype and function of CD63 + CAFs. Inspiringly, the pharmacological inhibition of CD63 + CAFs with a CD63-neutralizing antibody or cRGD-miR-22-sponge nanoparticles can effective reverse tamoxifen resistance in breast cancer [87].
CAF-derived EVs in cancer radiotherapy resistance
Radiotherapy is one of the mainstays of cancer treatment, and radiotherapy is widely used in the treatment of multiple malignancies, such as anaplastic thyroid cancer (ATC), and ESCC [88, 89]. Approximately half of cancer patients will receive radiotherapy either alone or in combination with other therapies during their treatment [90]. However, radioresistance is still a critical challenge resulting in radiotherapy failure, cancer relapses, and poor prognosis in clinical practice [91]. Recent work has indicated that radiation can mediate the contents and secretion of functional exosomes from cancer cells and other cell types within tumor microenvironment, resulting in radiotherapy resistance and radiation-induced bystander effects [92, 93]. For instance, exosomal miR-143-3p has been estimated to confer radiation resistance by inducing M2 macrophage polarization in locally advanced ESCC [94]. CSCs in tumours are mostly resistant to conventional therapies including ionizing radiation, and a high dose of radiation is required to eradicate CSCs. CAF-derived exosomes promoted radiotherapy resistance through enhancing CRC stemness by activating TGF-β signalling pathway [95]. Chen et al. have isolated and purified exosomes from the plasma of CRC patients by ExoQuick Exosome Precipitation Kit, and detected that miR-590-3p was upregulated in patients with radioresistance compared with patients with radiosensitivity. Besides, ectopic upregulation of miR-590-3p was detected in CAF-derived exosomes compared to normal fibroblast-derived exosomes, and CAF-derived exosomal miR-590-3p decreased CRC sensitivity to radiation. Mechanically, miR-590-3p inhibited the expression of CLCA4 and mediated the phosphorylation of PI3K and AKT in CRC cells [96]. Similarly, both in vitro and in vivo studies have demonstrated that miR93-5p was also transferred by exosomes from CAFs to CRCs, and CAF-derived exosomal miR-93-5p downregulated FOXA1 and reversed the suppression of FOXA1 on TGFB3, subsequently contributing to radiation resistance [97]. In lung cancer, in vitro experiment has revealed that CAFs derived exosomal miR-196a-5p was upregulated and induced radioresistance by sponging NFKBIA and activating the NF-κB signalling pathway [98]. FAP-positive CAFs, a specific CAF subpopulation in ESCC have been found to transfer exosomal lncRNA AFAP1-AS1 to ESCC cells, and AFAP-AS1 promoted the repairment of damaged DNA, conferring radiotherapy resistance of ESCC cells [99] (Table 2).
CAF-derived EVs in cancer immunotherapy resistance
Immunotherapy has ushered in a new era of cancer treatment and altered the therapeutic landscape for multiple human malignancies [100, 101]. The clinical goal of cancer immunotherapy is to prime the host immune system to provide passive or active immunity against malignant tumors [102, 103]. Rather than merely enhancing overall immunity, immunotherapy is also involved in the improvement of tumour immune microenvironment, potentially providing prolonged survival for cancer patients [104]. Various immunotherapies, including immune checkpoint inhibitors (ICIs), such as anti-PD-L1 drugs nivolumab and pembrolizumab, chimeric antigen receptor (CAR) T cells, tumor vaccines, and adoptive cell transfer (ACT), have shown promising curative effect on multiple cancers [105,106,107,108]. Despite tumour immunotherapy being approved for clinical use and achieving unprecedented results, resistance remains an obstacle to the application of immunotherapies [109]. Multiple studies have identified that CAFs were involved in mediating tumour immunotherapy resistance through immunosuppressive microenvironment generation, immune cell exclusion, and phenotype alternation [18]. For instance, signal-cell analysis revealed different CAF clusters, among which the cluster 0 CAFs (CAFs associated with ECM remodeling, collagen formation, and cell adhesion) upregulated the expression of PD-1 and cytotoxic T-lymphocyte associated protein 4 (CTLA4) on Treg cells, which, in turn, inhibited T cells function and induced immune resistance [110]. In breast cancer, CAFs inhibited ICIs efficiency by suppressing CD8 + T cell infiltration [111]. In HCC, prominent CD36 + CAFs exhibited high-level lipid metabolism and mediated macrophage migration inhibitory factor (MIF) expression by activating lipid peroxidation/p38/CEBPs axis. CD36 + CAFs also recruited CD33 + MDSCs, altering the tumour immune microenvironment [112]. CAF-derived EVs also play a critical role in regulating cancer immunotherapy resistance. For instance, CAF-derived exosomal miR-92 can significantly promote T cell apoptosis and confer immunotherapy resistance of breast cancer cells. Mechanically, miRNA-92 specifically bound to LATS2, which further interacted with YAP1, and chromatin immunoprecipitation confirmed that YAP1 could bind to the enhancer region of PD-L1 after nuclear translocation, promoting transcriptional activity [113]. CAFs can influence the efficacy of immunotherapy by interfering with the cytotoxic effects of T cells. In melanoma, studies have found that hypoxia can induce the secretion of immunosuppression factors such as TGF-β, VEGF, and PD-L1 from CAFs, which exerted an inhibitory effect on T cell-mediated cytotoxicity [114]. In CRC, the hypoxic microenvironment induced the production and excretion of circEIF3K-containing exosomes from CAFs compared with normoxic CAFs. CircEIF3 can reduce miR-214 to upregulate the expression of PD-L1, thereby inducing CRC cell proliferation, migration, metastasis, and immune escape [115]. In epithelial ovarian cancer (EOC), CAF-derived protein FMO2 facilitated the infiltration of lymphocytes. Higher level FMO2 was associated with worse prognosis in patients with similar levels of immune checkpoints such as PD-1 and PD-L1 suggesting that FMO2 could be a biomarker for predicting immunotherapy sensitivity [116]. In addition, it has been demonstrated that the CAF-derived EVs could mediate the immune escape of UBC by promoting PD-L1/PD-1 expression. Simultaneously, these EVs inhibited apoptosis in UBC cells, enhanced the invasion capability of T24 cells, and reduced the secretion of IFN-γ, IL-2, and TNF-α from CD8 + T cells, implying their potential role in inducing immunotherapy resistance of UBC [117].
Therapeutic potential of CAF-derived EVs
In the previous sections, we summarized the biological functions and detailed mechanisms of CAF-derived EVs in mediating cancer therapy resistance, providing evidence for the promising therapeutic potential of CAF-derived EVs in cancer management (Fig. 3). In this section, we will summarize and discuss the promising potential of CAF-derived EVs as clinical biomarkers and novel therapeutic targets for enhancing therapeutic efficacy.

CAF-derived EVs mediate cancer therapy resistance through complex mechanisms. CAF-derived EVs can deliver multiple cargoes to tumour cells, including miRNAs, lncRNAs, circRNAs and proteins. They can mediate the resistance to chemotherapy, targeted therapy, radiotherapy, and immunotherapy through multiple biological mechanisms, such as regulation of autophagy, EMT, and cancer stemness, metabolism, angiogenesis, DNA repair, immune infiltration, cell cycle, transcription and oncogenic signalling pathways
CAF-derived EVs as clinical biomarkers
Cancer is a highly heterogeneous disease involving diverse components, hence, developing effective cancer biomarkers is a critical way to represent cancer status and monitor cancer progression [118]. Liquid biopsy is a novel approach that collects biofluid specimens and provides more opportunities for early cancer diagnosis, prognosis prediction, and treatment efficacy evaluation [118, 119]. The large amounts of circulating EVs in biofluids produced by different cell types have become a critical component in liquid biopsies. For instance, Hoshino et al. have conducted a large-scale and comprehensive proteomic analysis of exosomes and exomeres from 426 human samples to develop novel diagnostic biomarkers [120]. Nowadays, a large number of clinical trials have been conducted to evaluate the therapeutic potential of EVs as diagnostic and prognostic biomarkers for diverse cancers. For instance, blood plasma samples from 420 lung cancer patients and 150 healthy controls were obtained to purify the exosomes in a recent clinical trial (ClinicalTrials.gov Identifier: NCT04529915). Deep-learning analysis of exosomes is performed to screen exosomal proteins for lung cancer early diagnosis and distinguish lung cancer patients in different clinical stages to improve the prognosis. In 2016, the first exosome-based liquid biopsy ExoDx™ Lung (ALK), was developed for the isolation and analysis of exosomal RNA from blood samples, which can effectively detect EML4-ALK mutations in NSCLC patients with 88% sensitivity and 100% specificity, thereby providing a more direct and sensitive approach to detect gene muutations than cfDNA [119]. The ExoDx Prostate IntelliScore (EPI), approved by the FDA, can effectively predict the risk of higher-grade prostate cancer for patients with PSA from 2 to 10 ng/mL by analyzing ERG, PCA3, and SPDEF RNA in exosomes [121]. Three independent, prospective, and multicenter clinical trials have reported that EPI can outperform the standard of care and serve as a novel biomarker for the early diagnosis of prostate cancer [122]. Another multicenter study has confirmed that GATA2 mRNA concentration in EVs in urine can effectively improve the detection of high-risk prostate cancer and may avoid unnecessary prostate biopsies [123]. In addition, MedOncAlyzer 170, a newly developed liquid biopsy system, has been developed for detecting exosomal RNA and ctDNA in a single trial. It can identify significant and functional mutations in multiple cancer types in a small volume (0.5 ml) of patient blood or plasma. Due to the unique way exosomes and ctDNA are formed, MedOncAlyzer 170 is accurate and highly sensitive to detect mutations at all stages of cancer progression and treatment [119].
Multiple experimental studies have evaluated the ectopic expression of diverse cargoes in CAF-derived EVs during cancer progression, and these cargoes are significantly correlated with clinical pathological features of cancer patients, thus highlighting the promising potential of CAF-derived EVs as biomarkers in the diagnosis and prognosis of various cancers. For instance, CAF-derived exosomal miR-196a is critically involved in HNC progression and chemoresistance, and plasma exosomal miR-196a levels were higher in HNC patients compared with healthy controls. Exosomal miR-196a levels were significantly higher in the plasma of patients with poor chemotherapeutic responses, larger tumor size, lymph node metastasis, and advanced tumor stage. Kaplan-Meier analysis and Cox proportional hazards regression analysis further confirmed that plasma exosomal miR-196a is a critical prognostic biomarker for HNC patients [124]. Similarly, miR-3188 is significantly downregulated in CAF-derived exosomes from HNC patients, and plasma miR-3188 levels were also downregulated in HNC patients and upregulated after tumour excision. A receiver operating characteristic (ROC) curve identified that plasma miR-3188 levels could serve as a diagnostic biomarker for HNC patients and low plasma miR-3188 expression was significantly correlated with larger tumor sizes and advanced TNM stage [125]. In HCC, miR-150-3p was significantly downregulated in CAF-derived exosomes and patients with low miR-150-3p levels in plasma exosomes had a worse prognosis [126]. Notably, several ongoing studies have been conducted to evaluate the potential of EVs as predictive biomarkers for cancer therapeutic efficacy. For instance, a previous study based on 18 melanoma and 8 NSCLC patients under anti PD-1 therapy has revealed that downregulation of PD-L1 mRNA in EVs was detected in patients with partial or complete immunotherapy response compared with those with progressive disease [127]. A recent clinical trial is designed to evaluate the response to immunotherapy (pablolizumab and nafulizumab) by measuring the PD-L1 and miRNA expression in plasma exosomes in pre- and post-treated NSCLC patients and correlates it with the treatment outcome (NCT04427475). Another ongoing clinical trial in Limoges (NCT03985696) is conducted to explore the expression of therapeutic targets (as CD20, PDL-1) in exosomes and immunotherapy resistance in patients with aggressive Non-Hodgkin B-cell lymphomas (B-NHL). Despite most of the current clinical trials being in Phase 0 or the initiation of recruitment, we believe that EVs will become promising and reliable clinical biomarkers and will eventually go to the clinic. In summary, diverse CAF-derived exosomal cargoes in body fluids across pan-cancers may function as non-invasive biomarkers in liquid biopsy for early diagnosis and clinical prediction.
CAF-derived EVs as therapeutic targets
Multiple lines of evidence have demonstrated that CAF-derived EVs actively participate in cancer tumorigenesis and progression. Hence, targeting CAFs and CAF-derived EVs has opened new avenues for cancer treatment. For instance, CAFs can promote intrahepatic cholangiocarcinoma progression by delivering miR-493-5p in EVs, highlighting that inhibiting miR-493-5p in CAFs may be a novel therapeutic strategy for intrahepatic cholangiocarcinoma treatment [128]. The CXCR4 antagonist motixafortide (BL-8040) has been found to inhibit the immunosuppressive CXCL12-CXCR4 axis mediated by FAP + CAFs and has been applied in combination with pembrolizumab and/or chemotherapy for the treatment of pancreatic cancer patients in phase II clinical trial (NCT02826486). CAF-derived ANXA6 EVs carry the ANXA6/LRP1/TSP1 complex to facilitate pancreatic cancer aggressiveness and depletion of ANXA6 in CAFs impaired complex formation and inhibited pancreatic cancer metastasis occurrence [129]. Inhibiting the biosynthesis and secretion of tumour-promoting EVs and blocking the endocytosis of tumour-promoting EVs by cancer cells have been investigated in multiple cancers recently. Several agents, such as tipifarnib and manumycin A have been reported to selectively inhibit tumour cell-released exosomes but exert no effects on normal cells [130]. Xi et al. have found that can stimulate ATM activation and facilitate autophagy-associated exosome release from CAFs to induce breast cancer cell invasion, and inhibition of oxidized ATM kinase by KU60019 (a small-molecule inhibitor of activated ATM) or shRNA effectively inhibits the exosome secretion from hypoxia-induced CAFs, subsequently inhibiting breast cancer progression [24]. Hence, specific inhibition of tumour-promoting EVs from CAFs may provide a novel method to inhibit cancer progression and reverse therapy resistance. In addition, the loss of specific cargoes in CAF-derived EVs may also contribute to cancer initiation and progression. For instance, miR-3188 was abnormally downregulated in CAF-derived exosomes in HNC, and loss of CAF-derived exosomal miR-3188 resulted in HNC progression in vivo and in vitro. Besides, miR-3188-loaded exosomes significantly inhibited tumor growth in vivo [125]. Hence, delivering anti-tumour agents by EVs may effectively inhibit tumour growth and improve patient prognosis. Inspiringly, several EV-delivered vaccines have entered clinical trials. For example, an ongoing phase II clinical trial (NCT01159288) is conducted to evaluate the safety and efficacy of dendritic cell-derived exosomes loaded with tumor antigens in 41 participants. Another early phase I clinical trial (NCT05559177) is designed to explore the therapeutic efficacy of chimeric exosomal tumor vaccines for recurrent or metastatic bladder cancer.
Perspectives and challenges
Multiple lines of evidence have revealed that CAF-derived EVs are critically involved in cancer therapy resistance with promising clinical significance and therapeutic potential. Targeting specific therapy resistance-associated CAF-derived EVs is a promising approach for overcoming cancer therapy resistance. In addition, CAF-derived EVs can function as noninvasive biomarkers in liquid biopsy, and may effectively improve the prediction of cancer patients’ therapeutic responses and prognosis [119]. Furthermore, engineered exosomes can function as a drug delivery system to deliver cargoes to specific cancer sites spatially and release the anti-cancer drugs temporally with enhanced efficacy and low side effects [131]. Recent studies have also illustrated that EVs may exert promising effects in repairing radiotherapy-related tissue damage by various mechanisms, such as promoting angiogenesis and inhibiting inflammation and fibrosis [132, 133]. Also, EVs have promising potential to treat cancer-related cachexia, which is a major complication of human malignancies and seriously affects the quality of life and prognosis of cancer patients [134]. Although promising progress has been achieved in illustrating the biological roles of CAF-derived EVs, several important unanswered questions and limitations should be resolved before the clinical application of CAF-derived EVs in cancer management.
Firstly, the categorization and classification of CAF subgroups lack a unified, standardized framework to address the diverse findings reported by different research teams, often within the same cancer category and utilizing comparable scRNA-seq or CyTOF methodologies. The subdivision and characterization of CAF subgroups appear subjective, leading to potential variations among observers [16]. Consequently, reaching a consensus on key biomarkers and hierarchical clustering patterns defining CAF subgroups through scRNA-seq or CyTOF analyses is imperative for the forthcoming period. Currently, there are still several challenges in applying scRNA-seq to investigate CAFs heterogeneity. For example, CAFs are difficult to isolate, because they are embedded in the ECM. Hence, CAFs are often under-represented in scRNA-seq investigation, and developing tissue-specific protocols for CAFs isolation is warranted [135]. Furthermore, the establishment of a standardized experimental platform featuring particular, specialized, and dependable tools and methodologies is essential for identifying, marking, segregating, cultivating, and functionally evaluating CAF subgroups both in artificial settings and within living organisms, akin to the protocols instituted for investigating immune cell populations. This standardization will facilitate the accurate delineation and description of CAF subgroups with designated functional roles and predictive significance, streamlining the design of tailored therapeutic strategies targeting specific CAF subpopulations or pathways in the future.
Secondly, the classifications for CAF-derived EVs remain elusive, with a recent study proposing isolation and characterization methods for six distinct EV subsets found in tissues. Despite these efforts, a universally accepted international categorization system for CAF-derived EVs is lacking, leaving doubts about the impact of inconsistent isolation and purification techniques on past findings. Future investigations ought to differentiate between various CAF-derived EV subsets and evaluate their anti-tumour potential. Additionally, the absence of systematic guidelines and procedures for the extraction, purification, and preservation of CAF-derived EVs poses a challenge to their clinical utilization. The employment of diverse isolation and purification approaches yields varying subpopulations of CAF-derived EVs possessing unique compositions, properties, and biological functions. Consequently, innovative standardized methodologies aligned with good manufacturing practices (GMP) are imperative for advancing CAF-derived EVs research.
Lastly, inhibiting the biogenesis and secretion of tumour-promoting CAF-derived EVs presents an innovative avenue for inhibiting cancer progression and therapy resistance, necessitating a profound comprehension of the biogenesis and release mechanisms of EVs. For instance, the Tspan family plays critical roles in regulating EV biogenesis, function, and targets, and Tspan proteins can regulate multifaced cancer behaviours and serve as clinical biomarkers [136]. Hence, targeting Tspan proteins may open new avenues for regulating EV biogenesis and functions and inhibiting tumour growth in anticancer therapeutic strategies [136, 137]. Nowadays, several EVs inhibitors have been investigated in cancer treatment, such as imipramine and pantethine for inhibiting EVs production, and bisindolylmaleimide I and macitentan for inhibiting EVs release [138]. Employing targeted siRNA or CRISPR/Cas9-loaded EVs, alongside multi-target pharmacological approaches to inhibit EV activity, also represents a novel therapeutic option for targeted CAF-derived EVs therapy. Nevertheless, challenges such as off-target effects, secure delivery modalities, nonspecific inhibition of non-cancerous EVs, and the triggering of compensatory pathways in EV formation and release pose significant hurdles. Beyond therapeutic efficacy, the reproducibility and efficiency of primary compounds for halting EV production necessitate further evaluation, especially since certain approved therapeutics with EV-targeting capabilities have displayed adverse effects in humans. For instance, imipramine, a tricyclic antidepressant, can result in undesirable outcomes, such as immune suppression and infection due to its interference with EVs formation via acid sphingomyelinase inhibition [139].
Conclusions
In conclusion, we discussed and summarized recent discoveries and research progress on the biological mechanisms of CAF-derived EVs in mediating therapy resistance in various cancers (Fig. 4). We further emphasized the clinical significance and implications of CAF-derived EVs in cancer management, which provides a novel direction for reversing cancer therapy resistance and improving patient prognosis. Due to the key role of CAF-derived EVs in mediating cancer therapy resistance, further in-depth studies are needed to illustrate the detailed mechanisms of CAF-derived EVs in cancer therapy failure and develop novel CAF-derived EVs-targeted therapeutic strategies to overcome cancer therapy resistance in the clinic.

Therapy resistance-related CAF-derived EVs in different cancers. CAF-derived EVs deliver miRNAs, lncRNAs, circRNAs and proteins to cancer cells, thus influence the therapy resistance of multiple cancer types, such as lung cancer, melanoma, and hepatocellular carcinoma. These cargoes in CAF-derived EVs may serve as novel therapeutic targets for reversing therapy resistance and clincal biomarkers for predicting therapy efficacy in cancer management
Availability of data and materials
Not applicable.
Data availability
Not Applicable.
Abbreviations
- ACT:
-
Adoptive cell transfer
- ALK:
-
Anaplastic Lymphoma Kinase
- APAF1:
-
Apoptosis protease activating fator1
- ATC:
-
Anaplastic thyroid cancer
- CAFs:
-
Cancer-associated fibroblasts
- CAR:
-
Chimeric antigen receptor
- cfDNA:
-
Cell free DNA
- circRNAs:
-
Circular RNAs
- CRC:
-
Colorectal cancer
- CSCs:
-
Cancer stem cells
- CTLA4:
-
Cytotoxic T-lymphocyte associated protein 4
- DDP:
-
Cisplatin
- DOX:
-
Doxorubicin
- DTX:
-
Docetaxel
- ECM:
-
Extracellular matrix
- EMT:
-
Epithelial-mesenchymal transition
- EOC:
-
Epithelial ovarian cancer
- EPI:
-
ExoDx Prostate IntelliScore
- ESCC:
-
Esophageal squamous cell carcinoma
- EVs:
-
Extracellular vesicles
- FGF5:
-
Fibroblast growth factor 5
- FMO2:
-
Flavin containing monooxygenase 2
- FOSL1:
-
Recombinant FOS Like Antigen 1
- GC:
-
Gastric Cancer
- GEM:
-
Gemcitabine
- GMP:
-
Good manufacturing practices
- HCC:
-
Hepatocellular carcinoma
- HGF:
-
Hepatocyte growth factor
- HNC:
-
Head and Neck Cancer
- HuR:
-
Human antigen R
- ICIs:
-
Immune Checkpoint Inhibitors
- ITGB4:
-
Integrin β4
- LncRNAs:
-
Long non-coding RNAs
- LSCC:
-
Laryngeal squamous cell carcinoma
- MIF:
-
Migration inhibitory factor
- miRNAs:
-
MicroRNAs
- M-MDSCs:
-
Monocytic myeloid-derived suppressor cells
- MTX:
-
Methotrexate
- ncRNAs:
-
Non-coding RNAs
- NFs:
-
Normal Fibroblasts
- NHEJ:
-
Non-homologous end joining
- NSCLC:
-
Non-small cell lung cancer
- OXA:
-
Oxaliplatin
- PDAC:
-
Pancreatic Ductal Adenocarcinoma
- PIAT:
-
PNI-associated transcript
- PTX:
-
paclitaxel
- SCC:
-
squamous cell carcinoma
- SCLC:
-
small cell lung cancer
- shRNA:
-
short hairpin RNA
- SNAIL:
-
Snail zinc finger transcription factor
- SPP1:
-
secreted phosphoprotein 1
- SUMO1:
-
Small Ubiquitin Like Modifier 1
- TKIs:
-
Tyrosine Kinase Inhibitors
- TP53INP1:
-
Tumor protein p53-inducible nuclear protein 1
- UBC:
-
urothelial bladder cancer
- VEGFA:
-
Vascular Endothelial Growth Factor A
- YBX1:
-
Recombinant Y-Box Binding Protein 1
References
Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA Cancer J Clin. 2023;73(1):17–48.
Gao H, Xi Z, Dai J, Xue J, Guan X, Zhao L, Chen Z, Xing F. Drug resistance mechanisms and treatment strategies mediated by ubiquitin-specific proteases (USPs) in cancers: new directions and therapeutic options. Mol Cancer. 2024;23(1):88.
Kim TK, Vandsemb EN, Herbst RS, Chen L. Adaptive immune resistance at the tumour site: mechanisms and therapeutic opportunities. Nat Rev Drug Discov. 2022;21(7):529–40.
Robertson BM, Fane ME, Weeraratna AT, Rebecca VW. Determinants of resistance and response to melanoma therapy. Nat Cancer. 2024;5(7):964–82.
Andrei L, Kasas S, Ochoa Garrido I, Stankovic T, Suarez Korsnes M, Vaclavikova R, Assaraf YG, Pesic M. Advanced technological tools to study multidrug resistance in cancer. Drug Resist Updat. 2020;48:100658.
Wang X, Jiang W, Du Y, Zhu D, Zhang J, Fang C, Yan F, Chen ZS. Targeting feedback activation of signaling transduction pathways to overcome drug resistance in cancer. Drug Resist Updat. 2022;65:100884.
Chan YT, Wu J, Lu Y, Li Q, Feng Z, Xu L, Yuan H, Xing T, Zhang C, Tan HY, Feng Y, Wang N. Loss of lncRNA LINC01056 leads to sorafenib resistance in HCC. Mol Cancer. 2024;23(1):74.
Zhang H, Dong X, Ding X, Liu G, Yang F, Song Q, Sun H, Chen G, Li S, Li Y, Wang M, Guo T, Zhang Z, Li B, Yang L. Bufalin targeting CAMKK2 inhibits the occurrence and development of intrahepatic cholangiocarcinoma through Wnt/beta-catenin signal pathway. J Transl Med. 2023;21(1):900.
Zhang H, Yang J, Song Q, Ding X, Sun F, Yang L. UBA3 promotes the occurrence and metastasis of intrahepatic cholangiocarcinoma through MAPK signaling pathway. Acta Biochim Biophys Sin (Shanghai). 2024;56(2):199–209.
Soumoy L, Ghanem GE, Saussez S, Journe F. Bufalin for an innovative therapeutic approach against cancer. Pharmacol Res. 2022;184:106442.
Zhang W, Fan Y, Zhang J, Shi D, Yuan J, Ashrafizadeh M, Li W, Hu M, Abd El-Aty AM, Hacimuftuoglu A, Linnebacher M, Cheng Y, Li W, Fang S, Gong P, Zhang X. Cell membrane-camouflaged bufalin targets NOD2 and overcomes multidrug resistance in pancreatic cancer. Drug Resist Updat. 2023;71:101005.
Wu SY, Fu T, Jiang YZ, Shao ZM. Natural killer cells in cancer biology and therapy. Mol Cancer. 2020;19(1):120.
Barry ST, Gabrilovich DI, Sansom OJ, Campbell AD, Morton JP. Therapeutic targeting of tumour myeloid cells. Nat Rev Cancer. 2023;23(4):216–37.
Lavie D, Ben-Shmuel A, Erez N, Scherz-Shouval R. Cancer-associated fibroblasts in the single-cell era. Nat Cancer. 2022;3(7):793–807.
Chen Y, McAndrews KM, Kalluri R. Clinical and therapeutic relevance of cancer-associated fibroblasts. Nat Rev Clin Oncol. 2021;18(12):792–804.
Biffi G, Tuveson DA. Diversity and Biology of Cancer-Associated fibroblasts. Physiol Rev. 2021;101(1):147–76.
Li C, Teixeira AF, Zhu HJ. Ten Dijke, Cancer associated-fibroblast-derived exosomes in cancer progression. Mol Cancer. 2021;20(1):154.
Zhang H, Yue X, Chen Z, Liu C, Wu W, Zhang N, Liu Z, Yang L, Jiang Q, Cheng Q, Luo P, Liu G. Define cancer-associated fibroblasts (CAFs) in the tumor microenvironment: new opportunities in cancer immunotherapy and advances in clinical trials. Mol Cancer. 2023;22(1):159.
Sahai E, Astsaturov I, Cukierman E, DeNardo DG, Egeblad M, Evans RM, Fearon D, Greten FR, Hingorani SR, Hunter T, Hynes RO, Jain RK, Janowitz T, Jorgensen C, Kimmelman AC, Kolonin MG, Maki RG, Powers RS, Pure E, Ramirez DC, Scherz-Shouval R, Sherman MH, Stewart S, Tlsty TD, Tuveson DA, Watt FM, Weaver V, Weeraratna AT, Werb Z. A framework for advancing our understanding of cancer-associated fibroblasts. Nat Rev Cancer. 2020;20(3):174–86.
Zhong D, Wang Z, Ye Z, Wang Y, Cai X. Cancer-derived exosomes as novel biomarkers in metastatic gastrointestinal cancer. Mol Cancer. 2024;23(1):67.
van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19(4):213–28.
Han T, Chen L, Li K, Hu Q, Zhang Y, You X, Han L, Chen T, Li K. Significant CircRNAs in liver cancer stem cell exosomes: mediator of malignant propagation in liver cancer? Mol Cancer. 2023;22(1):197.
Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478):640.
Xi L, Peng M, Liu S, Liu Y, Wan X, Hou Y, Qin Y, Yang L, Chen S, Zeng H, Teng Y, Cui X, Liu M. Hypoxia-stimulated ATM activation regulates autophagy-associated exosome release from cancer-associated fibroblasts to promote cancer cell invasion. J Extracell Vesicles. 2021;10(11):e12146.
Fang T, Lv H, Lv G, Li T, Wang C, Han Q, Yu L, Su B, Guo L, Huang S, Cao D, Tang L, Tang S, Wu M, Yang W, Wang H. Tumor-derived exosomal mir-1247-3p induces cancer-associated fibroblast activation to foster lung metastasis of liver cancer. Nat Commun. 2018;9(1):191.
Zheng S, Hu C, Lin Q, Li T, Li G, Tian Q, Zhang X, Huang T, Ye Y, He R, Chen C, Zhou Y, Chen R. Extracellular vesicle-packaged PIAT from cancer-associated fibroblasts drives neural remodeling by mediating m5C modification in pancreatic cancer mouse models. Sci Transl Med. 2024;16(756):eadi0178.
Holohan C, Van Schaeybroeck S, Longley DB, Johnston PG. Cancer drug resistance: an evolving paradigm. Nat Rev Cancer. 2013;13(10):714–26.
Ghosh S. Cisplatin: the first metal based anticancer drug. Bioorg Chem. 2019;88:102925.
Fang Y, Zhou W, Rong Y, Kuang T, Xu X, Wu W, Wang D, Lou W. Exosomal miRNA-106b from cancer-associated fibroblast promotes gemcitabine resistance in pancreatic cancer. Exp Cell Res. 2019;383(1):111543.
Czekay RP, Cheon DJ, Samarakoon R, Kutz SM, Higgins PJ. Cancer-Associated fibroblasts: mechanisms of Tumor Progression and Novel therapeutic targets. Cancers (Basel). 2022;14(5):1231.
Sun Z, Shi K, Yang S, Liu J, Zhou Q, Wang G, Song J, Li Z, Zhang Z, Yuan W. Effect of exosomal miRNA on cancer biology and clinical applications. Mol Cancer. 2018;17(1):147.
Zhang T, Zhang P, Li HX. CAFs-Derived exosomal miRNA-130a confers Cisplatin Resistance of NSCLC cells through PUM2-Dependent packaging. Int J Nanomed. 2021;16:561–77.
Shi L, Zhu W, Huang Y, Zhuo L, Wang S, Chen S, Zhang B, Ke B. Cancer-associated fibroblast-derived exosomal microRNA-20a suppresses the PTEN/PI3K-AKT pathway to promote the progression and chemoresistance of non-small cell lung cancer. Clin Transl Med. 2022;12(7):e989.
Zhao Q, Huang L, Qin G, Qiao Y, Ren F, Shen C, Wang S, Liu S, Lian J, Wang D, Yu W, Zhang Y. Cancer-associated fibroblasts induce monocytic myeloid-derived suppressor cell generation via IL-6/exosomal miR-21-activated STAT3 signaling to promote cisplatin resistance in esophageal squamous cell carcinoma. Cancer Lett. 2021;518:35–48.
Zhang H-W, Shi Y, Liu J-B, Wang H-M, Wang P-Y, Wu Z-J, Li L, Gu L-P, Cao P-S, Wang G-R, Ma Y-S, Fu D. Cancer-associated fibroblast-derived exosomal microRNA-24-3p enhances colon cancer cell resistance to MTX by down-regulating CDX2/HEPH axis. J Cell Mol Med. 2021;25(8):3699–713.
Mosca L, Ilari A, Fazi F, Assaraf YG, Colotti G. Taxanes in cancer treatment: activity, chemoresistance and its overcoming. Drug Resist Updat. 2021;54:100742.
Shan G, Gu J, Zhou D, Li L, Cheng W, Wang Y, Tang T, Wang X. Cancer-associated fibroblast-secreted exosomal mir-423-5p promotes chemotherapy resistance in prostate cancer by targeting GREM2 through the TGF-β signaling pathway. Exp Mol Med. 2020;52(11):1809–22.
Sun L, Ke M, Yin M, Zeng Y, Ji Y, Hu Y, Fu S, Zhang C. Extracellular vesicle-encapsulated microRNA-296-3p from cancer-associated fibroblasts promotes ovarian cancer development through regulation of the PTEN/AKT and SOCS6/STAT3 pathways. Cancer Sci. 2024;115(1):155–69.
Au Yeung CL, Co NN, Tsuruga T, Yeung TL, Kwan SY, Leung CS, Li Y, Lu ES, Kwan K, Wong KK, Schmandt R, Lu KH, Mok SC. Exosomal transfer of stroma-derived miR21 confers paclitaxel resistance in ovarian cancer cells through targeting APAF1. Nat Commun. 2016;7:11150.
Jiang X, Stockwell BR, Conrad M. Ferroptosis: mechanisms, biology and role in disease. Nat Rev Mol Cell Biol. 2021;22(4):266–82.
Zhang C, Liu X, Jin S, Chen Y, Guo R. Ferroptosis in cancer therapy: a novel approach to reversing drug resistance. Mol Cancer. 2022;21(1):47.
Zhang H, Deng T, Liu R, Ning T, Yang H, Liu D, Zhang Q, Lin D, Ge S, Bai M, Wang X, Zhang L, Li H, Yang Y, Ji Z, Wang H, Ying G, Ba Y. CAF secreted miR-522 suppresses ferroptosis and promotes acquired chemo-resistance in gastric cancer. Mol Cancer. 2020;19(1):43.
Qi R, Bai Y, Li K, Liu N, Xu Y, Dal E, Wang Y, Lin R, Wang H, Liu Z, Li X, Wang X, Shi B. Cancer-associated fibroblasts suppress ferroptosis and induce gemcitabine resistance in pancreatic cancer cells by secreting exosome-derived ACSL4-targeting miRNAs. Drug Resist Updat. 2023;68:100960.
Zhao J, Shen J, Mao L, Yang T, Liu J, Hongbin S. Cancer associated fibroblast secreted Mir-432-5p targets CHAC1 to inhibit ferroptosis and promote acquired chemoresistance in prostate cancer. Oncogene. 2024;43(27):2104–14.
Lin Z, Lu S, Xie X, Yi X, Huang H. Noncoding RNAs in drug-resistant pancreatic cancer: a review. Biomed Pharmacother. 2020;131:110768.
Cui Y, Zhang S, Hu X, Gao F. Tumor-associated fibroblasts derived exosomes induce the proliferation and cisplatin resistance in esophageal squamous cell carcinoma cells through RIG-I/IFN-β signaling. Bioengineered. 2022;13(5):12462–74.
Schuth S, Le Blanc S, Krieger TG, Jabs J, Schenk M, Giese NA, Büchler MW, Eils R, Conrad C, Strobel O. Patient-specific modeling of stroma-mediated chemoresistance of pancreatic cancer using a three-dimensional organoid-fibroblast co-culture system. J Exp Clin Cancer Res. 2022;41(1):312.
Richards KE, Xiao W, Hill RT. On Behalf of the Usc Pancreas Research, Cancer-Associated fibroblasts Confer Gemcitabine Resistance to Pancreatic Cancer cells through PTEN-Targeting miRNAs in Exosomes. Cancers. 2022;14(11):2812.
Erin N, Grahovac J, Brozovic A, Efferth T. Tumor microenvironment and epithelial mesenchymal transition as targets to overcome tumor multidrug resistance. Drug Resist Updat. 2020;53:100715.
Richards KE, Zeleniak AE, Fishel ML, Wu J, Littlepage LE, Hill R. Cancer-associated fibroblast exosomes regulate survival and proliferation of pancreatic cancer cells. Oncogene. 2017;36(13):1770–8.
Shan G, Zhou X, Gu J, Zhou D, Cheng W, Wu H, Wang Y, Tang T, Wang X. Downregulated exosomal microRNA-148b-3p in cancer associated fibroblasts enhance chemosensitivity of bladder cancer cells by downregulating the Wnt/β-catenin pathway and upregulating PTEN. Cell Oncol (Dordr). 2021;44(1):45–59.
Hu JL, Wang W, Lan XL, Zeng ZC, Liang YS, Yan YR, Song FY, Wang FF, Zhu XH, Liao WJ, Liao WT, Ding YQ, Liang L. CAFs secreted exosomes promote metastasis and chemotherapy resistance by enhancing cell stemness and epithelial-mesenchymal transition in colorectal cancer. Mol Cancer. 2019;18(1):91.
Wigerup C, Påhlman S, Bexell D. Therapeutic targeting of hypoxia and hypoxia-inducible factors in cancer. Pharmacol Ther. 2016;164:152–69.
Deng K, Zou F, Xu J, Xu D, Luo Z. Cancer-associated fibroblasts promote stemness maintenance and gemcitabine resistance via HIF-1α/miR-21 axis under hypoxic conditions in pancreatic cancer. Mol Carcinog. 2024;63(3):524–37.
Icard P, Shulman S, Farhat D, Steyaert JM, Alifano M, Lincet H. How the Warburg effect supports aggressiveness and drug resistance of cancer cells? Drug Resist Updat. 2018;38:1–11.
Ganapathy-Kanniappan S, Geschwind JF. Tumor glycolysis as a target for cancer therapy: progress and prospects. Mol Cancer. 2013;12:152.
Kunou S, Shimada K, Takai M, Sakamoto A, Aoki T, Hikita T, Kagaya Y, Iwamoto E, Sanada M, Shimada S, Hayakawa F, Oneyama C, Kiyoi H. Exosomes secreted from cancer-associated fibroblasts elicit anti-pyrimidine drug resistance through modulation of its transporter in malignant lymphoma. Oncogene. 2021;40(23):3989–4003.
Wu B, Shi X, Jiang M, Liu H. Cross-talk between cancer stem cells and immune cells: potential therapeutic targets in the tumor immune microenvironment. Mol Cancer. 2023;22(1):38.
Zhuang J, Shen L, Li M, Sun J, Hao J, Li J, Zhu Z, Ge S, Zhang D, Guo H, Huang R, Yan J. Cancer-Associated fibroblast-derived miR-146a-5p generates a Niche that promotes bladder Cancer Stemness and Chemoresistance. Cancer Res. 2023;83(10):1611–27.
Wang M, Zhuoma C, Liu X, Huang Q, Cai G, Sang G, Zhou L, Du H, Wu C. Loss of exosomal miR-34c-5p in cancer-associated fibroblast for the maintenance of stem-like phenotypes of laryngeal cancer cells. Head Neck. 2022;44(11):2437–51.
Ferrer J, Dimitrova N. Transcription regulation by long non-coding RNAs: mechanisms and disease relevance. Nat Rev Mol Cell Biol. 2024;25(5):396–415.
Luo G, Zhang Y, Wu Z, Zhang L, Liang C, Chen X. Exosomal LINC00355 derived from cancer-associated fibroblasts promotes bladder cancer cell resistance to cisplatin by regulating miR-34b-5p/ABCB1 axis. Acta Biochim Biophys Sin. 2021;53(5):558–66.
Hu JH, Tang HN, Wang YH. Cancer-associated fibroblast exosome LINC00355 promotes epithelial-mesenchymal transition and chemoresistance in colorectal cancer through the miR-34b-5p/CRKL axis. Cancer Gene Ther. 2024;31(2):259–72.
Gao Q, Fang X, Chen Y, Li Z, Wang M. Exosomal lncRNA UCA1 from cancer-associated fibroblasts enhances chemoresistance in vulvar squamous cell carcinoma cells. J Obstet Gynaecol Res. 2021;47(1):73–87.
Sun Y, Hao G, Zhuang M, Lv H, Liu C, Su K. MEG3 LncRNA from Exosomes released from Cancer-Associated fibroblasts enhances Cisplatin Chemoresistance in SCLC via a MiR-15a-5p/CCNE1 Axis. Yonsei Med J. 2022;63(3):229–40.
Zhu S, Mao J, Zhang X, Wang P, Zhou Y, Tong J, Peng H, Yang B, Fu Q. CAF-derived exosomal lncRNA FAL1 promotes chemoresistance to oxaliplatin by regulating autophagy in colorectal cancer. Dig Liver Dis. 2024;56(2):330–42.
Deng X, Ruan H, Zhang X, Xu X, Zhu Y, Peng H, Zhang X, Kong F, Guan M. Long noncoding RNA CCAL transferred from fibroblasts by exosomes promotes chemoresistance of colorectal cancer cells. Int J Cancer. 2020;146(6):1700–16.
Qu X, Liu B, Wang L, Liu L, Zhao W, Liu C, Ding J, Zhao S, Xu B, Yu H, Zhang X, Chai J. Loss of cancer-associated fibroblast-derived exosomal DACT3-AS1 promotes malignant transformation and ferroptosis-mediated oxaliplatin resistance in gastric cancer. Drug Resist Updat. 2023;68:100936.
Chen LL. The expanding regulatory mechanisms and cellular functions of circular RNAs. Nat Rev Mol Cell Biol. 2020;21(8):475–90.
Zhou Y, Tang W, Zhuo H, Zhu D, Rong D, Sun J, Song J. Cancer-associated fibroblast exosomes promote chemoresistance to cisplatin in hepatocellular carcinoma through circZFR targeting signal transducers and activators of transcription (STAT3)/ nuclear factor -kappa B (NF-κB) pathway. Bioengineered. 2022;13(3):4786–97.
Qu Z, Yang KD, Luo BH, Zhang F. CAFs-secreted exosomal cricN4BP2L2 promoted colorectal cancer stemness and chemoresistance by interacting with EIF4A3. Exp Cell Res. 2022;418(2):113266.
Hu C, Xia R, Zhang X, Li T, Ye Y, Li G, He R, Li Z, Lin Q, Zheng S, Chen R. circFARP1 enables cancer-associated fibroblasts to promote gemcitabine resistance in pancreatic cancer via the LIF/STAT3 axis. Mol Cancer. 2022;21(1):24.
Zheng S, Tian Q, Yuan Y, Sun S, Li T, Xia R, He R, Luo Y, Lin Q, Fu Z, Zhou Y, Chen R, Hu C. Extracellular vesicle-packaged circBIRC6 from cancer-associated fibroblasts induce platinum resistance via SUMOylation modulation in pancreatic cancer. J Exp Clin Cancer Res. 2023;42(1):324.
Lin S, Zhu B. Exosome-transmitted FOSL1 from cancer-associated fibroblasts drives colorectal cancer stemness and chemo-resistance through transcriptionally activating ITGB4. Mol Cell Biochem. 2024;479(3):665–77.
Claesson-Welsh L, Welsh M. VEGFA and tumour angiogenesis. J Intern Med. 2013;273(2):114–27.
Shi Y, Zhu H, Jiang H, Yue H, Yuan F, Wang F. Cancer-associated fibroblasts-derived exosomes from chemoresistant patients regulate cisplatin resistance and angiogenesis by delivering VEGFA in colorectal cancer. Anticancer Drugs. 2023;34(3):422–30.
Allen GM, Lim WA. Rethinking cancer targeting strategies in the era of smart cell therapeutics. Nat Rev Cancer. 2022;22(12):693–702.
Huang A, Garraway LA, Ashworth A, Weber B. Synthetic lethality as an engine for cancer drug target discovery. Nat Rev Drug Discov. 2020;19(1):23–38.
Aldea M, Andre F, Marabelle A, Dogan S, Barlesi F, Soria JC. Overcoming resistance to Tumor-targeted and Immune-targeted therapies. Cancer Discov. 2021;11(4):874–99.
Lim ZF, Ma PC. Emerging insights of tumor heterogeneity and drug resistance mechanisms in lung cancer targeted therapy. J Hematol Oncol. 2019;12(1):134.
Nilsson MB, Yang Y, Heeke S, Patel SA, Poteete A, Udagawa H, Elamin YY, Moran CA, Kashima Y, Arumugam T, Yu X, Ren X, Diao L, Shen L, Wang Q, Zhang M, Robichaux JP, Shi C, Pfeil AN, Tran H, Gibbons DL, Bock J, Wang J, Minna JD, Kobayashi SS, Le X, Heymach JV. CD70 is a therapeutic target upregulated in EMT-associated EGFR tyrosine kinase inhibitor resistance. Cancer Cell. 2023;41(2):340–55. e6.
Fiori ME, Di Franco S, Villanova L, Bianca P, Stassi G, De Maria R. Cancer-associated fibroblasts as abettors of tumor progression at the crossroads of EMT and therapy resistance. Mol Cancer. 2019;18(1):70.
Eun JW, Yoon JH, Ahn HR, Kim S, Kim YB, Lim SB, Park W, Kang TW, Baek GO, Yoon MG, Son JA, Weon JH, Kim SS, Cho HJ, Cheong JY. Cancer-associated fibroblast-derived secreted phosphoprotein 1 contributes to resistance of hepatocellular carcinoma to sorafenib and lenvatinib. Cancer Commun (Lond). 2023;43(4):455–79.
Qin W, Wang L, Tian H, Wu X, Xiao C, Pan Y, Fan M, Tai Y, Liu W, Zhang Q, Yang Y. CAF-derived exosomes transmitted Gremlin-1 promotes cancer progression and decreases the sensitivity of hepatoma cells to sorafenib. Mol Carcinog. 2022;61(8):764–75.
Zhao J, Lin E, Bai Z, Jia Y, Wang B, Dai Y, Zhuo W, Zeng G, Liu X, Cai C, Li P, Zou B, Li J. Cancer-associated fibroblasts induce sorafenib resistance of hepatocellular carcinoma cells through CXCL12/FOLR1. BMC Cancer. 2023;23(1):1198.
Zhang Y, Pan Q, Shao Z. Extracellular vesicles derived from cancer-associated fibroblasts carry tumor-promotive microRNA-1228-3p to enhance the resistance of hepatocellular carcinoma cells to sorafenib. Hum Cell. 2023;36(1):296–311.
Gao Y, Li X, Zeng C, Liu C, Hao Q, Li W, Zhang K, Zhang W, Wang S, Zhao H, Fan D, Li M, Zhang Y, Zhang W, Zhang C. CD63(+) Cancer-Associated fibroblasts Confer tamoxifen resistance to breast Cancer cells through Exosomal miR-22. Adv Sci (Weinh). 2020;7(21):2002518.
McLaughlin M, Patin EC, Pedersen M, Wilkins A, Dillon MT, Melcher AA, Harrington KJ. Inflammatory microenvironment remodelling by tumour cells after radiotherapy. Nat Rev Cancer. 2020;20(4):203–17.
Allen C, Her S, Jaffray DA. Radiotherapy for Cancer: Present and Future. Adv Drug Deliv Rev. 2017;109:1–2.
Chandra RA, Keane FK, Voncken FEM, Thomas CR Jr. Contemporary radiotherapy: present and future. Lancet. 2021;398(10295):171–84.
Kim TW. Fisetin, an anti-inflammatory Agent, overcomes Radioresistance by activating the PERK-ATF4-CHOP Axis in Liver Cancer. Int J Mol Sci. 2023;24(10):9076.
Barker HE, Paget JT, Khan AA, Harrington KJ. The tumour microenvironment after radiotherapy: mechanisms of resistance and recurrence. Nat Rev Cancer. 2015;15(7):409–25.
Peng J, Yin X, Yun W, Meng X, Huang Z. Radiotherapy-induced tumor physical microenvironment remodeling to overcome immunotherapy resistance. Cancer Lett. 2023;559:216108.
Gao LR, Zhang J, Huang N, Deng W, Ni W, Xiao Z, Liu M. Tumor-derived exosomal mir-143-3p induces macrophage M2 polarization to cause Radiation Resistance in locally advanced esophageal squamous cell carcinoma. Int J Mol Sci. 2024;25(11):6082.
Liu L, Zhang Z, Zhou L, Hu L, Yin C, Qing D, Huang S, Cai X, Chen Y. Cancer associated fibroblasts-derived exosomes contribute to radioresistance through promoting colorectal cancer stem cells phenotype. Exp Cell Res. 2020;391(2):111956.
Chen X, Liu Y, Zhang Q, Liu B, Cheng Y, Zhang Y, Sun Y, Liu J. Exosomal mir-590-3p derived from cancer-associated fibroblasts confers radioresistance in colorectal cancer. Mol Ther Nucleic Acids. 2021;24:113–26.
Chen X, Liu J, Zhang Q, Liu B, Cheng Y, Zhang Y, Sun Y, Ge H, Liu Y. Exosome-mediated transfer of mir-93-5p from cancer-associated fibroblasts confer radioresistance in colorectal cancer cells by downregulating FOXA1 and upregulating TGFB3. J Exp Clin Cancer Res. 2020;39(1):65.
Yao F, Shi W, Fang F, Lv MY, Xu M, Wu SY, Huang CL. Exosomal miR-196a-5p enhances radioresistance in lung cancer cells by downregulating NFKBIA. Kaohsiung J Med Sci. 2023;39(6):554–64.
Zhou X, Tong Y, Yu C, Pu J, Zhu W, Zhou Y, Wang Y, Xiong Y, Sun X. FAP positive cancer-associated fibroblasts promote tumor progression and radioresistance in esophageal squamous cell carcinoma by transferring exosomal lncRNA AFAP1-AS1. Mol Carcinog. 2024.
Kennedy LB, Salama AKS. A review of cancer immunotherapy toxicity. CA Cancer J Clin. 2020;70(2):86–104.
Riley RS, June CH, Langer R, Mitchell MJ. Delivery technologies for cancer immunotherapy. Nat Rev Drug Discov. 2019;18(3):175–96.
Schoenfeld AJ, Hellmann MD. Acquired Resistance to Immune Checkpoint inhibitors. Cancer Cell. 2020;37(4):443–55.
O’Donnell JS, Teng MWL, Smyth MJ. Cancer immunoediting and resistance to T cell-based immunotherapy. Nat Rev Clin Oncol. 2019;16(3):151–67.
Rui R, Zhou L, He S. Cancer immunotherapies: advances and bottlenecks. Front Immunol. 2023;14:1212476.
Akiyama T, Yasuda T, Uchihara T, Yasuda-Yoshihara N, Tan BJY, Yonemura A, Semba T, Yamasaki J, Komohara Y, Ohnishi K, Wei F, Fu L, Zhang J, Kitamura F, Yamashita K, Eto K, Iwagami S, Tsukamoto H, Umemoto T, Masuda M, Nagano O, Satou Y, Saya H, Tan P, Baba H, Ishimoto T. Stromal reprogramming through dual PDGFRα/β blockade boosts the efficacy of Anti-PD-1 immunotherapy in fibrotic tumors. Cancer Res. 2023;83(5):753–70.
Feins S, Kong W, Williams EF, Milone MC, Fraietta JA. An introduction to chimeric antigen receptor (CAR) T-cell immunotherapy for human cancer. Am J Hematol. 2019;94(S1):S3–9.
Saxena M, van der Burg SH, Melief CJM, Bhardwaj N. Therapeutic cancer vaccines. Nat Rev Cancer. 2021;21(6):360–78.
Restifo NP, Dudley ME, Rosenberg SA. Adoptive immunotherapy for cancer: harnessing the T cell response. Nat Rev Immunol. 2012;12(4):269–81.
Bashash D, Zandi Z, Kashani B, Pourbagheri-Sigaroodi A, Salari S, Ghaffari SH. Resistance to immunotherapy in human malignancies: mechanisms, research progresses, challenges, and opportunities. J Cell Physiol. 2022;237(1):346–72.
Kieffer Y, Hocine HR, Gentric G, Pelon F, Bernard C, Bourachot B, Lameiras S, Albergante L, Bonneau C, Guyard A, Tarte K, Zinovyev A, Baulande S, Zalcman G, Vincent-Salomon A. Mechta-Grigoriou, single-cell analysis reveals fibroblast clusters linked to Immunotherapy Resistance in Cancer. Cancer Discov. 2020;10(9):1330–51.
Jenkins L, Jungwirth U, Avgustinova A, Iravani M, Mills A, Haider S, Harper J, Isacke CM. Cancer-Associated fibroblasts suppress CD8 + T-cell infiltration and Confer Resistance to Immune-Checkpoint Blockade. Cancer Res. 2022;82(16):2904–17.
Zhu GQ, Tang Z, Huang R, Qu WF, Fang Y, Yang R, Tao CY, Gao J, Wu XL, Sun HX, Zhou YF, Song SS, Ding ZB, Dai Z, Zhou J, Ye D, Wu DJ, Liu WR, Fan J, Shi YH. CD36(+) cancer-associated fibroblasts provide immunosuppressive microenvironment for hepatocellular carcinoma via secretion of macrophage migration inhibitory factor. Cell Discov. 2023;9(1):25.
Dou D, Ren X, Han M, Xu X, Ge X, Gu Y, Wang X. Cancer-Associated Fibroblasts-Derived Exosomes Suppress Immune Cell Function in Breast Cancer via the miR-92/PD-L1 Pathway. Front Immunol. 2020;11:2026.
Ziani L, Buart S, Chouaib S, Thiery J. Hypoxia increases melanoma-associated fibroblasts immunosuppressive potential and inhibitory effect on T cell-mediated cytotoxicity. Oncoimmunology. 2021;10(1):1950953.
Yang K, Zhang J, Bao C. Exosomal circEIF3K from cancer-associated fibroblast promotes colorectal cancer (CRC) progression via miR-214/PD-L1 axis. BMC Cancer. 2021;21(1):933.
Yu S, Yang R, Xu T, Li X, Wu S, Zhang J. Cancer-associated fibroblasts-derived FMO2 as a biomarker of macrophage infiltration and prognosis in epithelial ovarian cancer. Gynecol Oncol. 2022;167(2):342–53.
Feng R, Li Z, Ge G, Wang C, Jia Y, Ouyang J. Cancer-associated fibroblast-derived Extracellular vesicles mediate Immune escape of bladder Cancer via PD-L1/PD-1 expression. Endocr Metab Immune Disord Drug Targets. 2023;23(11):1410–20.
Ma S, Zhou M, Xu Y, Gu X, Zou M, Abudushalamu G, Yao Y, Fan X, Wu G. Clinical application and detection techniques of liquid biopsy in gastric cancer. Mol Cancer. 2023;22(1):7.
Yu D, Li Y, Wang M, Gu J, Xu W, Cai H, Fang X, Zhang X. Exosomes as a new frontier of cancer liquid biopsy. Mol Cancer. 2022;21(1):56.
Hoshino A, Kim HS, Bojmar L, Gyan KE, Cioffi M, Hernandez J, Zambirinis CP, Rodrigues G, Molina H, Heissel S, Mark MT, Steiner L, Benito-Martin A, Lucotti S, Di Giannatale A, Offer K, Nakajima M, Williams C, Nogues L, Pelissier Vatter FA, Hashimoto A, Davies AE, Freitas D, Kenific CM, Ararso Y, Buehring W, Lauritzen P, Ogitani Y, Sugiura K, Takahashi N, Aleckovic M, Bailey KA, Jolissant JS, Wang H, Harris A, Schaeffer LM, Garcia-Santos G, Posner Z, Balachandran VP, Khakoo Y, Raju GP, Scherz A, Sagi I, Scherz-Shouval R, Yarden Y, Oren M, Malladi M, Petriccione M, De Braganca KC, Donzelli M, Fischer C, Vitolano S, Wright GP, Ganshaw L, Marrano M, Ahmed A, DeStefano J, Danzer E, Roehrl MHA, Lacayo NJ, Vincent TC, Weiser MR, Brady MS, Meyers PA, Wexler LH, Ambati SR, Chou AJ, Slotkin EK, Modak S, Roberts SS, Basu EM, Diolaiti D, Krantz BA, Cardoso F, Simpson AL, Berger M, Rudin CM, Simeone DM, Jain M, Ghajar CM, Batra SK, Stanger BZ, Bui J, Brown KA, Rajasekhar VK, Healey JH, de Sousa M, Kramer K, Sheth S, Baisch J, Pascual V, Heaton TE, La Quaglia MP, Pisapia DJ, Schwartz R, Zhang H, Liu Y, Shukla A, Blavier L, DeClerck YA, LaBarge M, Bissell MJ, Caffrey TC, Grandgenett PM, Hollingsworth MA, Bromberg J, Costa-Silva B, Peinado H, Kang Y, Garcia BA. E.M. O’Reilly, D. Kelsen, T.M. Trippett, D.R. Jones, I.R. Matei, W.R. Jarnagin, D. Lyden, Extracellular Vesicle and Particle Biomarkers Define Multiple Human Cancers. Cell. 182(4):2020:1044–1061 e18.
McKiernan J, Donovan MJ, O’Neill V, Bentink S, Noerholm M, Belzer S, Skog J, Kattan MW, Partin A, Andriole G, Brown G, Wei JT, Thompson IM Jr., Carroll P. A novel urine exosome gene expression assay to Predict High-grade prostate Cancer at initial biopsy. JAMA Oncol. 2016;2(7):882–9.
Margolis E, Brown G, Partin A, Carter B, McKiernan J, Tutrone R, Torkler P, Fischer C, Tadigotla V, Noerholm M, Donovan MJ, Skog J. Predicting high-grade prostate cancer at initial biopsy: clinical performance of the ExoDx (EPI) prostate intelliscore test in three independent prospective studies. Prostate Cancer Prostatic Dis. 2022;25(2):296–301.
Woo J, Santasusagna S, Banks J, Pastor-Lopez S, Yadav K, Carceles-Cordon M, Dominguez-Andres A, Den RB, Languino LR, Pippa R, Lallas CD, Lu-Yao G, Kelly WK, Knudsen KE, Rodriguez-Bravo V, Tewari AK, Prats JM, Leiby BE, Gomella LG. Domingo-Domenech, urine extracellular vesicle GATA2 mRNA discriminates Biopsy result in men with suspicion of prostate Cancer. J Urol. 2020;204(4):691–700.
Qin X, Guo H, Wang X, Zhu X, Yan M, Wang X, Xu Q, Shi J, Lu E, Chen W, Zhang J. Exosomal miR-196a derived from cancer-associated fibroblasts confers cisplatin resistance in head and neck cancer through targeting CDKN1B and ING5. Genome Biol. 2019;20(1):12.
Wang X, Qin X, Yan M, Shi J, Xu Q, Li Z, Yang W, Zhang J, Chen W. Loss of exosomal miR-3188 in cancer-associated fibroblasts contributes to HNC progression. J Exp Clin Cancer Res. 2019;38(1):151.
Yugawa K, Yoshizumi T, Mano Y, Itoh S, Harada N, Ikegami T, Kohashi K, Oda Y, Mori M. Cancer-associated fibroblasts promote hepatocellular carcinoma progression through downregulation of exosomal miR-150-3p. Eur J Surg Oncol. 2021;47(2):384–93.
Del Re M, Marconcini R, Pasquini G, Rofi E, Vivaldi C, Bloise F, Restante G, Arrigoni E, Caparello C, Bianco MG, Crucitta S, Petrini I, Vasile E, Falcone A, Danesi R. PD-L1 mRNA expression in plasma-derived exosomes is associated with response to anti-PD-1 antibodies in melanoma and NSCLC. Br J Cancer. 2018;118(6):820–4.
Toshida K, Itoh S, Harada N, Morinaga A, Yugawa K, Tomiyama T, Kosai-Fujimoto Y, Tomino T, Kurihara T, Nagao Y, Morita K, Oda Y, Yoshizumi T. Cancer-associated fibroblasts promote tumor cell growth via mir-493-5p in intrahepatic cholangiocarcinoma. Cancer Sci. 2023;114(3):937–47.
Leca J, Martinez S, Lac S, Nigri J, Secq V, Rubis M, Bressy C, Serge A, Lavaut MN, Dusetti N, Loncle C, Roques J, Pietrasz D, Bousquet C, Garcia S, Granjeaud S, Ouaissi M, Bachet JB, Brun C, Iovanna JL, Zimmermann P, Vasseur S, Tomasini R. Cancer-associated fibroblast-derived annexin A6 + extracellular vesicles support pancreatic cancer aggressiveness. J Clin Invest. 2016;126(11):4140–56.
Peng Z, Tong Z, Ren Z, Ye M, Hu K. Cancer-associated fibroblasts and its derived exosomes: a new perspective for reshaping the tumor microenvironment. Mol Med. 2023;29(1):66.
Zhang M, Hu S, Liu L, Dang P, Liu Y, Sun Z, Qiao B, Wang C. Engineered exosomes from different sources for cancer-targeted therapy. Signal Transduct Target Ther. 2023;8(1):124.
Zuo R, Liu M, Wang Y, Li J, Wang W, Wu J, Sun C, Li B, Wang Z, Lan W, Zhang C, Shi C, Zhou Y. BM-MSC-derived exosomes alleviate radiation-induced bone loss by restoring the function of recipient BM-MSCs and activating Wnt/beta-catenin signaling. Stem Cell Res Ther. 2019;10(1):30.
Lin Z, Wu Y, Xu Y, Li G, Li Z, Liu T. Mesenchymal stem cell-derived exosomes in cancer therapy resistance: recent advances and therapeutic potential. Mol Cancer. 2022;21(1):179.
Ru Q, Chen L, Xu G, Wu Y. Exosomes in the pathogenesis and treatment of cancer-related cachexia. J Transl Med. 2024;22(1):408.
Waise S, Parker R, Rose-Zerilli MJJ, Layfield DM, Wood O, West J, Ottensmeier CH, Thomas GJ, Hanley CJ. An optimised tissue disaggregation and data processing pipeline for characterising fibroblast phenotypes using single-cell RNA sequencing. Sci Rep. 2019;9(1):9580.
Zhang H, Song Q, Shang K, Li Y, Jiang L, Yang L. Tspan protein family: focusing on the occurrence, progression, and treatment of cancer. Cell Death Discov. 2024;10(1):187.
Garcia-Mayea Y, Mir C, Carballo L, Sanchez-Garcia A, Bataller M, ME LL. TSPAN1, a novel tetraspanin member highly involved in carcinogenesis and chemoresistance. Biochim Biophys Acta Rev Cancer. 2022;1877(1):188674.
Kim JH, Lee CH, Baek MC. Dissecting exosome inhibitors: therapeutic insights into small-molecule chemicals against cancer. Exp Mol Med. 2022;54(11):1833–43.
Catalano M, O’Driscoll L. Inhibiting extracellular vesicles formation and release: a review of EV inhibitors. J Extracell Vesicles. 2020;9(1):1703244.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from National Natural Science Foundation of China (NO. 81871783 and 82072441), Hunan Outstanding Youth Fund Project Grant (NO. 2022JJ10095).
Author information
Authors and Affiliations
Contributions
ZJL, and TL contributed to conceive, design and revision of the manuscript sections. ZJL, and GQL wrote the manuscript. KJ and GQL designed figures and created Tables. TL and ZH L supervised the manuscript by providing critical feedbacks and revisions. The authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
No datasets were generated or analysed during the current study.
Consent for publication
All authors consent to publication.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lin, Z., Li, G., Jiang, K. et al. Cancer therapy resistance mediated by cancer-associated fibroblast-derived extracellular vesicles: biological mechanisms to clinical significance and implications. Mol Cancer 23, 191 (2024). https://doi.org/10.1186/s12943-024-02106-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12943-024-02106-8