
Abstract
Background
Endometrial cancer is one of the major gynecological cancers, with increasing incidence and mortality in the past decades. Emerging preclinical and clinical data have indicated its close association with obesity and dyslipidemia. Metabolism reprogramming has been considered as the hallmark of cancer, to satisfy the extensive need of nutrients and energy for survival and growth. Particularly, lipid metabolism reprogramming has aroused the researchers’ interest in the field of cancer, including tumorigenesis, invasiveness, metastasis, therapeutic resistance and immunity modulation, etc. But the roles of lipid metabolism reprogramming in endometrial cancer have not been fully understood. This review has summarized how lipid metabolism reprogramming induces oncogenesis and progression of endometrial cancer, including the biological functions of aberrant lipid metabolism pathway and altered transcription regulation of lipid metabolism pathway. Besides, we proposed novel therapeutic strategies of targeting lipid metabolism pathway and concentrated on its potential of sensitizing immunotherapy and hormonal therapy, to further optimize the existing treatment modalities of patients with advanced/metastatic endometrial cancer. Moreover, we expect that targeting lipid metabolism plus hormone therapy may block the endometrial malignant transformation and enrich the preventative approaches of endometrial cancer.
Conclusion
Lipid metabolism reprogramming plays an important role in tumor initiation and cancer progression of endometrial cancer. Targeting the core enzymes and transcriptional factors of lipid metabolism pathway alone or in combination with immunotherapy/hormone treatment is expected to decrease the tumor burden and provide promising treatment opportunity for patients with advanced/metastatic endometrial cancer.
Similar content being viewed by others
Background
Endometrial cancer (EC) has become one of the most prevalent gynecological malignancies with 420, 242 newly diagnosed and 97,704 death patients worldwide [1]. Due to lifestyle risk factors like obesity and the lacking of treatment advancement, the incidence and mortality of EC have continued to rise for decades, directly threatening females’ health and survival [2, 3]. Surgery alone or combined with radiation and/or platinum-based chemotherapy as well as hormone therapy can generate good efficacy in most EC patients [4]. However, the 5-year survival rates of patients with stage III/IV tumors remain only 48% and 15% respectively [5]. What’s worse, the median overall survival (OS) of recurrent/metastatic patient is extremely short, ranging from 12 to 15 months [6]. Recently, adjuvant immunotherapy therapy has provided extra treatment opportunity for advanced and metastatic EC patients, but there is ultimately unmet need in the management of almost half of mismatch repair deficient EC and the majority of mismatch repair proficient EC that include copy-number low and copy-number high (P53 mutant) tumors [7, 8]. Thus, it’s necessary to further explicit the biological mechanism of cancer progression, and identify corresponding new therapeutic strategies in EC.
EC is a metabolism disorders-related malignancy [9,10,11,12]. Until now, over 50% of EC may be attributed to obesity [3]. Besides, obesity is related to earlier age at diagnosis of EC, suggesting its role in accelerating tumorigenesis [13]. Adipose tissues not only contribute to a proinflammatory microenvironment, but also produce overloaded estrogen, both contributing to the risk of endometrial cancer [4]. Remarkably, dyslipidemia is an independent predictor of endometrial cancer, which promotes the tumorigenesis through obesity [14]. The traditional clinicopathological classification of EC is associated with metabolic risk factors. Bokhman divided EC into two subgroups: Type I tumors are mainly endometrioid, characterized by obesity, hyperglycemia, hyperlipidemia and hyperestrogenism, sensitive to hormone therapy, frequently > 50 years old, with favorable prognosis; Type II tumors are almost serous, insensitive to hormone treatment, usually < 50 years old, with worse clinical outcomes [15]. A large-sample epidemiological research has demonstrated metabolism risk factors like obesity, previously linked to type I EC are also significantly associated with the incidence risk of more aggressive type II EC [16], indicating the metabolic disturbance in tumor progression. In consistent with this, there was a significant increasing trend of the mortality risk in overweighted or obese EC patients [17]. Actually, obesity is the intersection of complex biological processes.
Metabolism reprogramming has been recognized as the hallmark of cancer. Metabolism network was rearranged to induce the transformation of normal cells, and to promote the division of tumor cells in nutrient-depleted conditions [18]. Metabolic adaption also affected the tumor immunity and promote metastasis [19,20,21]. Tumor cells tended to exploit glycolysis even in aerobic microenvironment, called the “Warburg effect”, which is rewired by tumorigenic mutations in key genes like TP53 and KRAS. Tumoral glycolysis will provide necessary metabolites for lipid synthesis and regulating lipid metabolism pathway [22]. Lipid plays a pivotal role in oncological biology, including serving as constitutional component of cellular bio-membrane, fueling cell growth and mediating signaling transduction, etc [23]. Lipid metabolism reprogramming involving carcinogenesis, invasiveness, metastasis, therapy resistance and immunity of cancer has greatly drawn the researchers’ attention [24,25,26].
Therefore, we will concentrate on the biologic functions of lipid metabolism reprogramming and aberrant transcriptional regulation of lipid metabolism pathway in endometrial cancer. Importantly, we’ll propose the therapeutic strategies of targeting lipid metabolism pathway and summarize ongoing development of agents under preclinical and clinical assessment, in an attempt to renew the current intervention landscape of EC patients.
The biological functions of lipid metabolism reprogramming in endometrial cancer
Altered lipid uptake promoting tumor proliferation and progression
Abnormal fatty acid uptake
Fatty acid (FA) involves in cellular membrane structure, energy production and signal transduction [27]. Exogeneous FAs serve as an important source of cancer cell growth, dependent on the transportation of several membranous proteins, including CD36, fatty acid transport protein (FATP) and fatty acid binding protein (FABP) [28] (Fig. 1).
It’s reported that CD36 can promote FA uptake via endocytosis mediated by palmitoylation [29]. CD36 was generally upregulated in cancer and could promote tumor growth, metastasis and resistance [30, 31]. CD36 deletion attenuated the liver metastasis of cancer in mice model [32]. Besides, metabolism reprogramming mediated by CD36 induced tumor resistance to anti-HER2 therapy in breast cancer [33]. In pancreatic ductal adenocarcinoma, the expression of CD36 led to gemcitabine-resistant tumor by regulating anti-apoptotic protein [34]. Su et al [35] reported, visceral obesity and metabolic syndrome were directly associated with EC. Adiponectin inhibited the expression of its targeted gene CD36 via downregulating the transcriptional activity of STAT3 signaling, thus reducing peroxisome proliferator-activated receptor (PPAR) activity and suppressing adipocyte differentiation. In contrast, Knapp et al [36] found in grade I, FIGO I endometrioid EC, FA transporters (CD36 and FATP1) were expressed significantly lower than glucose transporter, indicating that CD36 dependent FA uptake may not serve as the main energetic source in early stage, low grade of EC when glucose can be efficiently supplied. But the specific mechanisms of this metabolic switch in EC have not been fully understood. Though some debates seem still existing, the expression and effect of CD36 in glucose-depleted EC will stimulate the research activity.
FABP is able to bind to ligands like long chain FAs. Some literatures reported the dysregulated expression of FABPs facilitated tumor progression [37]. FABP was overexpressed in colon cancer tissue with enhanced lipids accumulation, and promoted the invasiveness and migration by activating AKT pathway and epithelial-mesenchymal transition (EMT) [38]. Also, the upregulation of FABP5 was observed in liver carcinoma, which drove tumor growth via the activation of PI3K/AKT pathway [39]. Moreover, obese breast cancer patients showed significant higher level of circulating FABP, probably because free FABP released from fat tissues stimulated IL-6/STAT3/ALDH signaling pathway, further potentiating cancer cell stemness and invasion [40]. In EC, mass immune blot and immunohistochemistry both indicated significantly increased epidermal FABP (FABP5), in line with spectrometry analysis [41]. Another study demonstrated that compared to normal endometrium and endometrial intraepithelial neoplasia, FABP5 was significantly higher and associated with enhanced immune responsiveness and worse histodifferentiation in EC [42]. However, FABP4 protein and mRNA were reduced in EC. The overexpression of FABP4 exerted inhibitory effect in tumor growth and metastasis by blocking the activation of PI3K/AKT pathway [43], so FABP4 may serve as a tumor suppressor. In short, different subtypes of FABPs have distinct biological effects in EC, which may be the candidate therapeutic target and prognostic biomarker. How to regulate certain isoform of FABPs precisely may be the major challenge in treatment development.

Lipid uptake and its biological effects in endometrial cancer. FA, fatty acid; FABP, fatty acid binding protein; FATP, fatty acid transportation protein; LDL-C, low-density lipoprotein-cholesterol; LDLR, low-density lipoprotein receptor (By Figdraw)
Abnormal cholesterol uptake
Cholesterol belongs to sterols, as an essential constitutional component of cellular membrane, involved in signaling transduction and biosynthesis [44, 45]. Activated cholesterol uptake was usually observed in proliferative cancer cells. Low-density lipoprotein receptor (LDLR) is a single-transmembrane protein, mediating low-density lipoprotein-cholesterol (LDL-C) into cancer cell and maintaining cholesterol homeostasis [46, 47] (Fig. 1). The expression of LDLR and LDL-C were abnormally increased, and the former was an independent prognostic factor in some malignancies [47]. Previous research demonstrated that hypercholesterolemia contributed to the tumor growth and silencing LDLR reversed the tumor-promoting effect [48]. A Mendelian randomization study showed elevated LDL-C can predict good prognosis in EC, including both endometrioid and non-endometrioid tumors [49]. More importantly, after excluding possible mixed effects of obesity, LDL-C was still significantly related to reduced risk of incidence in EC, especially non-endometrioid diseases [49]. In EC, though the tumor inhibitory effect of LDL-C has been approved, accessible data of LDLR in tumor occurrence and patients’ prognosis are quite limited. So, the definite expression level of LDLR should be determined to better explain the involvement of cholesterol uptake process in EC.
FA and cholesterol uptake can not only provide exogeneous energy for tumor growth and proliferation, but also promote tumor invasiveness and migration by signaling transduction. Though many studies have proved enhanced lipid uptake represents unfavorable outcome in a variety of cancers, some studies have reported its contradictory effects in endometrial cancer. As we know, EC is a malignant tumor with lipid metabolism disorder. More basic and clinical studies are required to clarify whether endometrial cancer has a unique relation to lipid uptake dysregulation.
Aberrant de novo lipogenesis facilitating tumor growth and metastasis
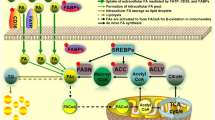
Despite the dependence of exogeneous lipid uptake, endogenous lipid de novo synthesis account for major lipid provision in cancer cell. Lipid biosynthesis mainly contains the synthesis of FA, cholesterol and phospholipid. As the key substrate for FA and cholesterol generation, acetyl-CoA was produced with the catalyzation of ATP-citrate lyase (ACLY) and acetyl-CoA synthetase 2 (ACSS2) [50, 51]. FA production requires acetyl-CoA carboxylase (ACC) and fatty acid synthase (FASN), and monounsaturated fatty acid (MUFA) was then produced by stearic acid desaturase (SCD) [28]. Cholesterol was synthesized by 3-hydroxy-3-methyl glutaryl coenzyme A reductase (HMGCR) and squalene monooxygenase (SM), further esterified into cholesteryl ester by acyl CoA: cholesterol acyltransferase (ACAT). Moreover, phospholipid was synthesized with CTP: phosphocholine cytidylyltransferase catalyzation and phospholipid remodeling was catalyzed by lysophosphatidylcholine acyltransferase. Potentiated lipid synthesis has been considered to promote tumor generation and progression, so comprehensively exploring the influence of disturbed lipid synthesis pathway and related key enzymes expression on tumor development in EC, will be beneficial to predict patients’ prognosis and identify novel hallmarks and therapeutic targets of EC (Fig. 2).

FAs and cholesterol provision in endometrial cancer. Lipid provision in endometrial cancer includes cellular uptake of exogeneous lipids and intracellular lipid synthesis. Fatty acid uptake is mediated by CD36 and fatty acid transport (FATP) and cholesterol is absorbed by low-density lipoprotein receptor (LDLR) in the form of low-density lipoprotein-cholesterol (LDL-C). Acetyl-CoA is the central metabolic source for de novo lipogenesis, which is mainly derived from citrate produced in tricarboxylic acid (TCA) cycle by ATP-citrate lyase (ACLY) catalyzation, thus closely linking the three major nutrients metabolism pathways to downstream lipid metabolism pathway. Also, acetyl-CoA is partly converted from cytoplasmic acetate with acetyl-CoA synthetase 2 (ACSS2) involvement, especially under the condition of ACLY deletion or metabolic stress. FAs and cholesterol are synthesized with acetyl-CoA, under the control of key enzymes. On one hand, Acetyl-CoA is catalyzed by acetyl-CoA carboxylase (ACC) to produce malonyl CoA. Then, palmitic acid is synthesized by fatty acid synthase (FASN), and subsequently desaturated to monounsaturated fatty acid (MUFA) by stearic acid desaturase 1 (SCD1). MUFAs can be further transformed to PUFAs and/or used to generate glycerophospholipids. On the other hand, HMG-CoA is synthesized with acetyl-CoA and acetoacetyl-CoA, then to produce mevalonate by 3-hydroxy-3-methyl glutaryl coenzyme A reductase (HMGCR) catalyzation. Squalene is converted to 2,3-oxidoaualene with SQLE involvement, to produce cholesterol after some steps. In particular, palmitic acid participates in triglyceride (TG) formation and cholesterol is esterified by acyl CoA: cholesterol acyltransferase (ACAT) to CE. Both TG and cholesteryl ester (CE) can be stored in lipid droplets (LDs) (By Figdraw)
Enhanced acetyl-CoA biosynthesis
Acetyl-CoA is at the core of cancer metabolism, which plays a critical role in FA and cholesterol synthesis, and histone acetylation, thus affecting the proliferative and invasive ability of cancer cell [52]. ACLY could drive transformation of citrate into acetyl-CoA, linking three major nutrients catabolic pathways to lipid biosynthesis [53]. ACLY was highly expressed in malignant tumors and can be an independent predictor of cancer recurrence risk [54,55,56]. Additionally, various mechanisms-induced ACLY upregulation has strongly promote the stemness and/or progression in colon cancer and breast cancer [57,58,59]. In contrast to this, downregulating ACLY would reduce intracellular lipid synthesis and accumulation by the ubiquitination and degradation of PPARγ, a transcriptional regulator of lipid metabolism pathway [60]. What’s more, acetyl-CoA can epigenetically regulate cancer cell proliferation via histone acetylation. Dai et al [61] found that ACLY mRNA and protein were both hyper-expressed in EC and were obviously correlated to some malignant clinicopathological characteristics and relatively higher BMI. Moreover, following mechanism exploration indicated AKT contributed to tumor cell proliferation via the intracellular nucleus translocation of ACLY, to potentiate histone acetylation and the expression of pyrimidine metabolism genes (like DHODH). Caspase-10 could inhibit acetyl-CoA synthesis by decomposing ACLY, and simultaneously decrease lipid and histone acetylation, effectively suppressing tumor proliferation and metastasis [62]. Thus, ACLY may elicit tumor-stimulatory effects by activating acetyl-CoA production through directly activating downstream lipid metabolism and other mechanisms like epigenetic regulation of DNA biosynthesis in EC.
In recent years, ACSS has been increasingly reported to capture cytoplasmic acetate as a carbon source in cancer, particularly in condition of ACLY genetic deletion or metabolic stress [51, 63, 64]. Among ACSS family members, the oncological biofunction of ACSS2 in cancer has been better described [65]. ACSS2 overexpression was observed in some solid tumor, including breast cancer, liver cancer and prostate cancer, predicting patients’ negative prognosis [66]. ACSS2 can also mediate histone acetylation to augment tumorigenesis. In particular, ACSS2 was highly expressed in obese myeloma patients, and prompted interferon regulator 4 (oncogene) mediated gene transcription by activating acetylation. At the same time, ACSS2 inhibition suppressed tumor growth in obese mouse model [67], indicating that ACSS2 may also play an important role in other obesity-induced tumors, especially EC.
The upregulation of acetyl-CoA synthesis pathway, featured by activated key enzymes like ACLY, is linked to cancer progression of obese malignant tumors and EC, which can fuel tumor growth and metastasis via stimulating acetylation and driving the downstream lipid anabolism pathway. Due to this, key enzymes in acetyl-CoA production will be potential target for EC with unbalanced lipid metabolism.
Potentiated FA biosynthesis
Acetyl-CoA being converted to malonyl CoA by ACC is the first and speed-limiting step in FA synthesis pathway. Then, acetyl-CoA added to malonyl CoA by FASN is to generate long chain saturated FA (or called palmitate), which will be desaturated to monounsaturated fatty acid (MUFA) by stearoyl-CoA desaturase [68].
ACC is divided into ACC1 and ACC2. ACC1 is located in cytoplasm, responsible for FA synthesis while ACC2 is situated at mitochondrial outer-membrane, mainly regulating FA uptake and β-oxidation [69]. ACC can enhance FA synthesis to provide needy energy for tumor cell survival and growth. Adenosine monophosphate activates protein kinase (AMPK) mediated dysregulated ACC1 and ACC2 phosphorylation may potentiate fat accumulation and cell proliferation [70]. Compared to the healthy tissues, the expression levels of ACC1 and ACC2 were increased in EC tissues [71]. Modesitt et al [72] found that in EC tissue of obese women, the inhibitory phosphorylation of ACC by AMPK was reduced with hepatic kinase B1 at a low level, partly explaining the enhanced fat production in these patients. These studies suggest that increased expression/activity of ACC can promote the cancer cell growth through activating FA biosynthesis, which is likely to be associated with obese EC. But direct proofs concerning the influences of ACC overexpression on cancer invasion and metastasis should be provided, and whether ACC can serve as a targetable molecule also requires further exploration in EC.
FASN took control of cancer cell viability and growth by synthesizing FA [73]. In EC, the expression and function of FASN have drawn much attention. It’s reported that the expression level of FASN gradually increased with disease progression as benign endometrium hyperplasia and endometrial dysplasia developed into EC [74]. FASN was expressed in most endometrioid adenocarcinoma and closely associated with the ability of tumor invasiveness and metastasis as well as higher BMI [75], which may independently predict the recurrence risk of EC [76]. Moreover, the overexpression of FASN tends to strengthen the carcinogenicity of EC. FASN expression was upregulated by SNHG25 and RAC3, facilitating the tumoral proliferation, invasiveness and migration [77, 78], while FASN inhibition might induce apoptosis of Ishikawa cell line [79]. In term of this, FASN upregulation is likely to accelerate malignant transformation of endometrium and aggressive biological behaviors, and can be an unfavorable prognostic marker of EC patients.
SCD catalyzed the production of MUFAs, mainly includes palmitoleic acid (16:1) and oleic acid (18:1), which subsequently were used to synthesize more complex lipids, like triglycerides and phospholipids [27, 80]. There are two types of SCD in human, SCD1 and SCD5, and the role of SCD1 is relatively definite [81]. A series of researches demonstrated SCD1 protected cancer cells from ferroptosis threatening, and inhibiting SCD1 effectively suppressed tumor growth and induced cell death [82,83,84], thus potentially beneficial to prevent cancer recurrence and metastasis. Interestingly, wild-type P53 was able to downregulate the expression of SCD1 by binding to its transcriptional promoter [85], suggesting that P53 mutant tumors may present aberrant lipid metabolism. A study reported that compared to secretory-phase and postmenopausal endometrium, SCD1 was significantly highly expressed in proliferative-phase, hyperplastic and malignant endometrium. More importantly, reduced SCD1 expression showed inhibitory effect on tumor growth in mice [86]. These findings reveal that SCD1 overexpression may contribute to maintaining cancer cell viability and promoting cancer progression, and SCD1 inhibition has significant killing effect on cancer cell, shedding light on treatment strategy establishment for EC.
Abnormally activated FA biosynthesis lays a solid foundation of cancer cell growth in EC, which can be effectively reversed by inhibiting FA production. The upregulation of FA de novo synthesis can be an unfavorable prognostic marker of EC patients, characterized by increased FASN and SCD expression, suggesting that enhanced FA synthesis may be involved in EC progression.
Increased cholesterol biosynthesis
Cholesterol biosynthesis is manipulated by HMGCR and SM, which are responsible for converting HMG-CoA to MVA (namely mevaleric acid pathway) and squalene to 2,3-epoxy squalene, respectively [87]. Recently, increased cholesterol synthesis has been considered to accelerate oncogenesis and tumor progression.
HMGCR was reported to be correlated with the risk of various cancers, and be involved in tumor growth and metastasis regulation [88,89,90,91]. FASN silencing could upregulate the expression of HMGCR by activating sterol regulatory element binding protein 2 (SREBP2), to enhance cholesterol production and cancer cell growth [92]. Besides, genetic alternation-induced HMGCR inactivation indicated reduced risk of ovarian cancer and breast cancer [93, 94], which lays a theoretic basis for the reuse of statins in anti-tumor treatment. Notably, analysis of transcriptomic sequencing data based on ovarian cancer, EC and cervix cancer showed that cholesterol related genes may contribute to tumoral resistance through cholesterol homeostasis and FA metabolism pathway [95]. These preliminary findings inspire researchers to explore the practicability of targeting HMGCR and predict tumoral response to anti-tumor therapy based on HMGCR expression status in EC.
SM, also called squalene cyclooxygenase (SQLE), was reported to promote tumor growth and progression in cancer [96,97,98,99,100]. Ma et al [101] has recently reported that significantly overexpressed SQLE was observed in EC patients by TCGA data analysis and IHC validation. Besides, SQLE drove the proliferation and metastasis of EC cell lines (Ishikawa and KLE) via upregulating PI3K/AKT signaling in vitro, which added the proofs of abnormal cholesterol metabolism to existing literatures in EC. Coates et al [102] initially revealed the upregulated truncation of SQLE could contribute to lipogenesis in EC. Hence, SQLE is commonly overexpressed to endogenously provide cholesterol for cancer cell and will become another therapeutic target in EC.
Acyl-CoA: cholesterol acyltransferase (ACAT), can transform free cholesterol into cholesterol ester, thus alleviating the cellular toxicity by overloaded free cholesterol [103]. There are two subtypes of ACAT in mammal animal as reported: ACAT1 (widely distributed) and ACAT2 (preferred in intestine and liver) [104]. ACAT was significantly correlated with recurrence-free survival in cancer [105]. For EC, a study demonstrated the activity of ACAT in malignant endometrium was significantly greater than healthy secretory endometrium [106]. A study based on bioinformation analysis demonstrated that compared to normal tissue, seven cholesterol homeostasis related genes (like ACAT2) were significantly upregulated, and the constructed cholesterol homeostasis related genes signature could serve as a predictive tool of poor survival outcome for EC patients [107]. In all, increased expression of ACAT is able to facilitate the prognosis prediction of cancers, including EC, and reducing the expression of ACAT inhibits tumor growth. So, ACAT-mediated cholesterol esterification may play a critical role in tumor cell survival, which is worth further investigation in EC.
Previous researches have showed that the upregulated cholesterol biosynthesis pathway can boost malignant tumor survival and growth in EC. In addition, the key enzyme of cholesterol esterification, ACAT, is highly expressed in EC tissues, indicating the cholesterol metabolism related tumor protective mechanisms to some degree.
Abnormal lipid catabolism exerting diverse roles
Fat acid oxidation
Fat acid oxidation (FAO) is mainly conducted by mitochondrial β-oxidation and is able to drive the tumoral proliferation, metastasis and resistance, etc [108]. The initial program of FAO is that FAs are activated to acyl-CoA by long-chain acyl-CoA synthetase (ACSL). In the following, lipoylcarnitine is synthesized with acyl-CoA, catalyzed by carnitine palmitoyltransferase1 (CPT1) in outer mitochondrial membrane, and subsequently entry into the mitochondria, which is the speed-decisive step in FAO process [108] (Fig. 3). Quan et al [109] clarified that ACSL3 was upregulated by TGFβ1 via SREBP1 and stimulated ATP and NADPH generation by triggering FAO pathway, thus facilitating EMT and tumor spread. Tang et al [110] reported that high CPT1 expression greatly enhanced tumor proliferation, non-anchored growth and lesions formation. In EC, the abnormal expression or genetic changes of ACSL is linked to malignant biological characteristics and patients’ prognosis. Zhou et al. [111] reported that ACSL1 overexpression upregulated FAO process and energy provision by activating AMPK/CPT1C/ATP pathway, thus promoting cancer cell to proliferate and migrate, which was also predominantly associated with malignant advancement of EC. Yu et al [112] using UALCAN database found that compared to normal tissue, ACSL4 protein was relatively higher in EC lesions, consistent with the results from TIMER and GEPIA databases. According to HPA database, ACSL4 protein presented strongly positive in EC tissue. At the same time, ACSL4 genetic mutation suggested longer progression-free survival period of patients with EC while copy number amplification of ACSL4 predicted much worse outcome of the patients [112]. In short, FAO pathway upregulation may serve as a useful predictor for unfavorable prognosis of EC, and key enzymes like ACSL and CPT1 represent promising treatment targets in EC.

The key enzymes and products of FAO in endometrial cancer. ACSL, long-chain acyl-CoA synthetase; CPT1, carnitine palmitoyltransferase1; FAO, fatty acid oxidation; TCA, tricarboxylic acid (By Figdraw)
Cholesterol efflux
Cholesterol homeostasis is necessary for maintaining cancer cell survival and growth. Of note, over-accumulation of cholesterol is harmful to cancer cells. Cholesterol efflux has become an effective strategy to avoid lipid toxicity in cancer cell, because cholesterol is difficult to absolutely break down in human peripheral tissues. Four conventional cholesterol efflux pathways include: passive diffusion, SR-B1 promoting diffusion, ATP binding cassette subfamily A member 1 (ABCA1) mediating efflux to pre-β1 high-density lipoprotein (HDL) and ATP binding cassette subfamily G member 1 (ABCG1) mediating efflux to mature HDL [113, 114]. In EC, Antmen et al [115] discovered that ABCA1 was significantly higher in tumor tissues that control group. Interestingly, as the histodifferentiation of endometrial adenocarcinoma went worse (from grade 1 to grade 3), ABCA1 expression level presented a relatively decreasing trend, suggesting the tumor promoting role of increased cholesterol in cancer cell by reduced ABCA1 in endometrioid cancer. Besides, ABCA1 activation by NNMT1 can enhance cholesterol efflux and cellular membrane fluidity, thus stimulating EMT process of EC [116]. Additionally, higher HDL-cholesterol apparently increased the risk of non-endometrioid tumor [49], indicating cholesterol efflux may accelerate the development of EC with more malignant histological subtypes. Until now, ABCA1-mediated cholesterol efflux has been considered to be an adaptive approach of cancer cell to maintain cholesterol homeostasis and survival in EC. But whether some molecules of other cholesterol efflux ways will be involved in the intra-and extra-cellular cholesterol regulation and the underlying mechanisms should be further elucidated in EC.
Altered transcriptional regulation of lipid metabolism
SREBP
SREBP is a kind of transcription factors. Nuclear SREBP will activate lipid uptake and synthesis-related genes of lipid metabolism pathway by sterol regulating element (SRE). There have been three well-known SREBPs: SREBP1a, mainly regulating reactive genes in FA and cholesterol de novo synthesis processes; SREBP1c, affecting FA synthesis related genes and involving lipocyte differentiation, whose downstream enzymes includes ACC, FASN and SCD1;SREBP2, responsible for regulating cholesterol uptake and synthesis by influencing LDLR and HMGCR [24, 117,118,119].
SREBP1 and SREBP2 as well as downstream enzymes regulated by them were involved in lipid metabolism reprogramming and disease progression in cancer [120, 121]. Shafiee et al [122] reported SREBP1 was significantly elevated in EC tissues than in health tissues, positively associated with serum triglycerides. Li et al [123] found that compared to normal tissues, SREBP1 was significantly upregulated in poorly differentiated EC tissues and mainly located in nucleus, indicating its tumor-promoting effect in EC. Besides, SREBP1 target molecules (like FASN, SCD and ACLY) were overexpressed in AN3CA EC cell line, and the expression of these key enzymes were accordingly reduced by SREBP1 downregulation by shRNA. Subsequent experiments demonstrated SREBP1 knockdown hindered proliferation and growth, and showed tumor killing effects in EC. Qiu et al [124] analyzed the SREBP1 gene in EC lesions (n = 30) and benign specimens (n = 6) based on high throughput sequencing, with single nucleotides polymorphisms (rs2297508) as the target marker. Consequently, single nucleotides polymorphisms (rs2297508) and C alleles was detected in 40% of EC, significantly more frequent than control group and linked to the histological differentiation and involvement of myometrium. The above studies demonstrate that the upregulation and some specific genetic variations of SREBP1 define malignant characteristics of EC, so SREBP1 inhibition may facilitate tumor regression by reducing lipid provision (endogenous synthesis and exogeneous uptake).
Liver X receptor
Liver X receptor (LXR), a nuclear receptor super family member, includes LXRα and LXRβ. LXR can act as a transcriptional regulator of SREBP1c and maintain cholesterol homeostasis [118, 125]. For one thing, LXR is combined and activated by a variety of oxidized sterols, then binding to retinol X receptor to form a heterodimer, specifically enhancing SREBP1c transcription to increase FA synthesis [126, 127], further promoting free cholesterol esterification to reduce extra toxicity from free cholesterol. For another, LXR activation could positively regulate ABCA1 expression to potentiate cholesterol efflux, and ubiquitously suppress LDLR by inducing Idol expression at transcription level, thus reducing cholesterol uptake [125, 128]. Otherwise, LXR activation could also induce the transcription of phospholipid remodeling enzyme (LPCAT3), preferentially driving the incorporation of unsaturated FAs into phospholipids to maintain the membrane homeostasis [129]. Fang et al [130] found by immunohistochemistry analysis that compared to normal and endometrial polyp tissues, LXRα was apparently overexpressed in endometrioid EC lesion and mainly distributed in cytoplasm of Ishikawa cell line. In addition, LXR impaired endometrial adenocarcinoma cell viability by inhibiting cyclin D1 and cyclin E expression. 27- hydroxycholesterol, a product of cholesterol metabolism, could be an endogenous activator of LXR and estrogen receptor [131]. Gibson et al [132] reported 27- hydroxycholesterol selectively activated estrogen receptor signaling to stimulate proliferation of Ishikawa and RL95-2 cells, which was reversed by external activation of LXR. Given that LXR has multiple effects on lipid metabolism regulation, systematic knowledge of LXR and of its upstream/downstream regulation mechanisms in EC should be established, to guide the exploration of effective therapeutic agents targeting LXR.
PPAR
PPAR belongs to nuclear hormone receptor super family and regulates lipid metabolism as a ligand-induced transcription factor [133, 134]. There are three subtypes of PPAR: PPARα, regulating FA transportation, esterification and oxidation processes; PPARβ/δ, mainly involving in FA oxidation; PPARγ, enhancing energy accumulation by affecting FA transportation, fat production and FA biosynthesis [133]. PPAR was reported to participate in diverse biological processes, such as tumor proliferation, vessels formation, invasion/metastasis, self-renewing, metabolism and immunology, etc [135].
Until now, related studies have demonstrated PPAR activation may exert anti-tumor effects in EC. A series of data supported PPARα and PPAR β/δ were expressed more strongly in EC tissues than non-malignant tissues, while the expression level of PPARγ was relatively decreased in EC [136,137,138,139,140]. But another study has reported that the expression of PPARα was lower in EC tissues than normal tissues recently [141]. Despite the distinct expression profiles of PPARα and PPARγ, their activation both inhibited tumor cell proliferation and angiogenesis in EC [140, 141]. Moreover, PPARγ activation attenuated proliferation, migration and invasion abilities of EC cells (especially EEC-1) and downregulated the estrogen receptor expression in estrogen receptor-positive EEC-1 cell line [138]. Additionally, activated PPARβ/δ exerted anti-tumor effect by inhibiting AKT/GSK3β/β-catenin pathway [142]. Previous studies have revealed both PPAR activation and inhibition have shown anti-tumor potential, which broaden our perspectives of complicated lipid metabolism transcriptional regulation and will arouse researchers’ interest to further investigate the underlying mechanisms and evaluate the effectiveness of targeting PPARs in EC. The key point is how to balance the status of PPARs to hinder the tumor progression of EC.
Targeting lipid metabolism as monotherapy in endometrial cancer
Growing studies have shown lipid metabolism alternations are associated with the occurrence, invasion, metastasis and recurrence of various cancers (including EC). Therefore, targeting key enzymes and related transcription factor of lipid metabolism pathway could be potential therapeutic strategy to inhibit cancer progression in EC (Fig. 4). Until now, a series of drugs targeting lipid metabolism pathway have been used as monotherapy to evaluate preclinical and clinical anti-tumor activity in EC.

Potential molecular targets and relevant agents of lipid metabolism in endometrial cancer. sterol regulatory element binding protein (SREBP) activates lipid uptake and synthesis-related genes of lipid metabolism pathway by sterol regulating element (SRE). SREBP2 is responsible for regulating cholesterol uptake and synthesis by influencing low-density lipoprotein receptor (LDLR) and 3-hydroxy-3-methyl glutaryl coenzyme A reductase (HMGCR). Activated liver X receptor (LXR) binds to retinol X receptor (RXR) to form a heterodimer, specifically enhancing SREBP1c transcription to increase fatty acid (FA) synthesis and maintaining cholesterol homeostasis. LXR activation can also positively regulate ATP binding cassette subfamily A member 1 (ABCA1) expression to potentiate cholesterol efflux, and ubiquitously suppress LDLR. Peroxisome proliferation activates receptor (PPAR) regulates lipid metabolism by influencing FA uptake, FA synthesis and fatty acid oxidation (FAO). Notably, fatty acid synthase (FASN), stearic acid desaturase (SCD1), 3-hydroxy-3-methyl glutaryl coenzyme A reductase (HMGCR), squalene cyclooxygenase (SQLE), SREBP and PPAR are potential therapeutic targets of aberrant lipid metabolism in endometrial cancer (By Figdraw)
FASN inhibitor
FASN is speed-limiting in FA synthesis pathway, and its overexpression will activate FA synthesis process, promoting EC invasion and migration. Due to this, inhibiting FA metabolism is a charming tumor interventional strategy and FASN inhibitors are increasingly developed in anti-tumor treatment. Orlistat, a robust FASN inhibitor, has been proved to reprogram oncological biology [143]. Wysham et al [144] assessed the anti-tumor activity of orlistat in ECC-1 and KLE cell lines and found orlistat at 500µM level could make the proliferative activity of cancer cells decrease by 61% and 57%, respectively. What’s more, Western blot analysis indicated orlistat reduced the expression of core enzymes (like FASN, ACC and CPT1A) in lipid metabolism pathway, upregulated phosphorylated AMPK expression and downregulated phosphorylated AKT expression. Mycotoxin cerulenin is a covalently bound inactivator of FASN and C75 is a novel small molecular inhibitor of FASN. Menendez et al [79] reported that cerulenin and C75 treatment remarkably abrogated tamoxifen-activated estrogen response element-relied transcription and estradiol-induced upregulation of oncogenes (like ERα and HER2), thus probably preventing malignant transformation of endometrium and inhibiting the viability and proliferation of EC cells. Excitingly, a clinical trial first evaluated safety and pharmacology of FASN inhibitor TVB-2640 (denifanstat) alone (n = 76) or combined with paclitaxel (n = 60) in advanced solid tumors (EC included). TVB-2460 monotherapy was able to accomplish a disease control rate of 42%. TVB-2640 combined with paclitaxel would reach 70% disease control rate and 12% partial response rate, with manageable drug safety [145]. Preclinical studies reveal that FASN inhibitors have initial anti-tumor activity and more clinical trials are in need to evaluate the feasibility of applying FASN inhibitors in EC patients with lipid metabolic disorder.
SCD1 inhibitor
SCD1 is responsible for the generation of unsaturated FAs, involving cancer ferroptosis resistance and metastasis, which has been considered as a potent target for cancer therapy [84, 146]. SCD1 knockdown or inhibited by A939572 damaged the tumor cell growth, promoted apoptotic program in vitro and abrogated the capability of metastatic lesions formation of EC AN3CA cell line in vivo [86].
HMGCR inhibitor
HMGCR, key enzyme of cholesterol biosynthesis, is highly expressed in cancer, to satisfy the extra need of cholesterol for tumor growth. Statins are able to regulate lipids level by inhibiting HMGCR and have been widely used in cardiovascular diseases management. More importantly, statins were reported to reduce the risk of tumorigenesis and improve prognosis benefits in cancers [147]. Hence, the repurposing of statins in anti-tumor field has become a hot topic [148]. Simvastatin is first-generation HMGCR inhibitor. Schointuch et al [149] found simvastatin treatment significantly inhibited the proliferation, invasion and migration ability of ECC-1 and Ishikawa cell lines as well as primary cancer cells derived from EC patients. Zhou et al. revealed [150] that simvastatin could block cancer cell proliferation and invasion by downregulating the RAS/MAPK signaling and induce reactive oxygen species and ferroptosis. Moreover, simvastatin plus metformin showed synergistic anti-tumor effects in RL95-2, HEC-1B and Ishikawa EC cell lines, possibly mediated by mTOR signaling and apoptosis [151]. Fluvastatin is the first completely synthesized second-generation HMGCR inhibitor. Cai et al [152]reported that fluvastatin impaired aggressive characteristics of EC cells and induced cell death. It should be emphasized that a clinical trial (NCT0276736) has accomplished the preliminary assessment of anti-proliferation activity of atorvastatin in EC.
SQLE inhibitor
SQLE is another rate-limiting enzyme for cholesterol biosynthesis. SQLE inhibitor like terbinafine and NB-598 effectively promoted the apoptosis and inactivated MAPK and PI3K/AKT pathway [153]. SQLE knockdown by shRNA or chemical inhibition by terbinafine could repress the P53-deficiency induced tumorigenesis and growth [154]. Du et al [155] proposed that SQLE was a promising therapeutic target in P53 mutant colorectal cancer, enlightening the novel treatment strategy of P53 mutant tumors. Silencing or pharmacological inhibition by terbinafine obviously reduced tumor growth of EC, indicating the therapeutic potential of SQLE inhibitor [101].
SREBP inhibitor
SREBP transcriptionally regulates lipid biosynthesis in various malignant tumors, and inhibiting SREBP may become a potent anti-tumor strategy. Fatostatin is an inhibitory agent of SREBP. Gao et al [156] revealed that fatostatin treatment could suppress the viability, clone formation and aggressiveness of EC cells, and exerted tumor killing effects by initiating cancer cell apoptosis, where SREBP1 and its downstream lipid synthases were noticeably downregulated with decreased intracellular free FAs and cholesterol. Similarly, Yao et al [157] also discovered fatostatin resulted in tumor regression by targeting lipid metabolism by in vitro and in vivo experiments. Research results in xenograft model have demonstrated fatostatin suppressed tumor growth and expansion, simultaneously disturbing tumor cell cycle and promoting cell death in EC. BF175, a new SREBP inhibitor, has shown lipid-lowering effects. Wang et al [158] initially reported BF175 dramatically hindered cell proliferation and migration of AN3CA cells by triggering autophagy and mitochondrial pathway associated apoptosis. In addition, BF175 also effectively downregulated SREBP and its downstream genes, to reduce FAs and cholesterol production. Furthermore, novel anti-tumor drug silibinin was considered to block lipid synthesis by inactivating STAT3 and SREBP signaling, causing damaged cell proliferation and activated apoptosis in EC [159]. Though in vitro and in vivo experiments suggest that SREBP inhibitor contribute to tumor inhibition, its safety and activity should be determined in more basic and clinical studies.
PPAR modulator
PPAR acted as a transcriptional regulator of lipid metabolism, involved in a variety of oncological biological processes, including lipids transportation, synthesis and catabolism, etc. PPAR signaling also plays an important role in tumor immune response. Interestingly, PPARs have shown bilateral functions in cancer development. PPAR modulators (agonist and antagonist) were both reported to have great cancer management potential [133]. Fibrates can serve as PPARα agonist. Fenofibrate effectively improved the sensitivity of breast cancer, prostate cancer and non-small cell lung cancer to chemotherapy or targeted therapy [160,161,162]. In EC, fenofibrate was able to suppress cancer cell expansion and stimulate apoptosis probably by inhibiting PPAR regulating FA metabolism pathway [163, 164]. Irbesartan, as a PPAR agonist, can also inhibit the proliferation of EC HEC-1 A and Ishikawa cell lines, by simultaneously inactivating the SREBP1 and activating the tumor suppressing genes [141]. Classical PPARγ agonists are known as isthiazolidinediones, containing troglitazone, rosiglitazone, pioglitazone, ciglitazone and inolitazone/efatutazone, etc. Kumari et al [165] first investigated the anti-tumor efficacy of pioglitazone in EC mouse model (n = 50) and found that compared to standard paclitaxel treatment, pioglitazone inhibited tumor lesions formation and growth in a dose dependent manner. Furthermore, GW0742 and GW501516 were two highly selective PPARβ/δ activators, able to activate PPAR signaling pathway, leading to the inhibition of tumoral proliferative capacity and apoptosis in EC [142]. Thus, the anti-tumor effect on part of PPAR modulators (mainly agonists), has been preliminarily determined in EC, but the anti-tumor efficacy of some PPAR inhibitors should be highlighted in future studies.
Collectively, targeting lipid metabolism pathway and related transcriptional regulation factors is a promising anti-tumor therapy in malignant tumors, and relevant agents has entered clinical trials (Table 1). The expansion of clinical trials in pan-cancer has encouraged the basic and clinical exploration of lipid metabolism-targeted therapies in EC (Table 2). Therefore, we could reasonably predict that targeting lipid metabolism will become spotlight of EC research in the approaching future, to prevent the development of EC and to improve the prognosis of EC patients, particularly in women with obesity or lipid metabolism disturbance.
Targeting lipid metabolism in combinational therapy of endometrial cancer
Given that solely targeting lipid metabolism pathway demonstrates promising therapeutic potential in EC management, its combination with immunotherapy is increasingly in development. Among lipid metabolism pathway targeted drugs, PPAR modulators have been mostly reported to improve the anti-tumor efficacy of immunotherapy. Interestingly, in diet-induced obese female mice, GW9662 combined with anti-PD-L1 immunotherapy showed anti-tumor activity. A clinical study (NCT04114136) has been designed to explore whether PPARγ antagonist (rosiglitazone) can remodel tumor immune microenvironment to potentiate tumor killing effects of T cells and improve the tumoral sensitivity to anti-PD-1 treatment in solid tumors. Also, the safety and tolerance of rosiglitazone combined with nivolumab/pembrolizumab are closely monitored. In addition, NCT03829436 is a phase 1 clinical trial, aiming to determine the drug toxicity and maximum tolerant dosage of PPARα antagonist (TSPT-1120) alone or combined with nivolumab (PD-L1 inhibitor) in advanced solid tumor. NCT04524871 is designed to evaluate clinical efficacy of different combinations of immunotherapy in advanced liver cancer, including atezolizumab (PD-1 inhibitor), bevacizumab and TSPT-1120.
Simultaneously, progesterone is applied in systematic management of EC, but tumor resistance to progesterone has severely decreased its clinical efficacy. Inspiringly, the strategy of targeting lipid metabolism pathway combined with progesterone is undergoing preliminary clinical investigation. A phase 2 clinical study (NCT05675787) is recruiting patients, in order to assess the efficacy of MPA plus atorvastatin in young patients with early-stage EC. Another phase 2 clinical trial (NCT04491643) is to investigate the anti-tumor effects of MA in combination with rosuvastatin in young, stage I EC patients. Similarly, Progesterone regimen and several statins are applied in patients with early-stage EC (NCT06102863).
Therefore, these active clinical researches strongly support that targeting lipid metabolism pathway sensitizing immunotherapy has become a potentially promising anti-tumor strategy in advanced solid tumor, and particularly in EC (Table 3). On one hand, anti-PD-1/PD-L1 immunotherapy has made breakthrough in EC chemical intervention, so it’s necessary to further investigate whether lipid metabolism targeted therapy added to immunotherapy can rescue more patients with EC. On the other hand, the therapeutic efficacy of lipid metabolism pathway targeted drugs (especially statins) plus progesterone is required to elucidate its potential as a conservative treatment for women at high risk of EC (such as obesity, metabolic syndrome and dyslipidemia) and to improve the survival of EC patients.
Conclusion
The increasing incidence and morality of EC can be largely attributed to metabolic disorders like obesity and hyperlipidemia. Lipid metabolism reprogramming is initiated by genetic alternations especially P53 mutation, which is remarkably associated with the worst prognosis of EC patients. In general, the activated endogenous lipogenesis (FA and cholesterol biosynthesis) and FAO process play a pivotal role in inducing tumor initiation, proliferation, invasion, metastasis and stemness, as well as tumor microenvironment remodeling and therapeutic resistance in EC. Besides, various signaling pathways are rewired by aberrant lipid metabolism to elicit tumor-promoting functions (Fig. 5). The alternations of some key enzymes (especially FASN, SCD1, HMGCR and SQLE) and transcriptional factors (SREBP, LXR and PPAR) define the lipid metabolism dysregulation in tumorigenesis and progression of EC, which has become the excellent candidate targets for treatment exploration. To date, drugs targeting lipid metabolism related pathway have shown preliminary anti-tumor activity in EC. Importantly, sensitizing hormonal therapy by targeting lipid metabolism pathway is undergoing assessment in early phase clinical trials in EC.

The landscape of aberrant lipid metabolism and potential mechanisms in tumorigenesis and cancer progression of endometrial cancer (By Figdraw)
Although the close association between lipid metabolism reprogramming and EC has been established, there is still a long way to go to dissect the roles of lipid metabolism pathways. Firstly, to what degree the lipid uptake and its core driving molecules can be involved in the initiation and progression of EC remains contradictory. Next, the transcriptional factor, PPARs, show diverse expression profiles, which leads to the exploration of their precision targeting strategy challenging. What’s more, hormone in combination with lipid metabolism targeted treatment has been applied in clinical trials, but the definite efficacy of preventing endometrial malignant transformation and inhibiting cancer progression is awaiting robust evidence. More importantly, sensitizing immunotherapy by rewiring the lipid metabolism is a promising direction for advanced/metastatic EC management due to the encouragement of several clinical trials in other malignancies, and the theoretical basis and clinical benefits should be further determined.
In conclusion, lipid metabolism reprogramming has become a hallmark of EC, which not only participates in the occurrence and advancement of tumor, but also has proposed as an interventional strategy. As studies increasingly accumulate, it’s expected to depict a comprehensive landscape of lipid metabolism in EC. In the future, researchers should focus on clarifying the pathological roles of different branches of lipid metabolism pathway and their interaction with the tumor microenvironment, and developing novel targetable molecules to decrease the tumor burden and hinder the cancer progression.
Availability of data and materials
No datasets were generated or analysed during the current study.
Abbreviations
- ABCA1:
-
ATP binding cassette subfamily A member 1
- ABCG1:
-
ATP binding cassette subfamily G member 1
- ACAT:
-
Acyl-CoA: cholesterol acyltransferase
- ACC:
-
Acetyl-CoA carboxylase
- ACLY:
-
ATP-citrate lyase
- ACSS2:
-
Acetyl-CoA synthetase 2
- ACSL:
-
Long-chain acyl-CoA synthetase
- CPT1:
-
Carnitine palmitoyltransferase1
- EC:
-
Endometrial cancer
- EMT:
-
Epithelial-mesenchymal transition
- FA:
-
Fatty acid
- FAO:
-
Fatty acid oxidation
- FASN:
-
Fatty acid synthase
- FATP:
-
Fatty acid transport protein
- HDL:
-
High-density lipoprotein
- HMGCR:
-
3-hydroxy-3-methyl glutaryl coenzyme A reductase
- LDLR:
-
Low-density lipoprotein receptor
- LDL-C:
-
Low-density lipoprotein-cholesterol
- LXR:
-
Liver X receptor
- MUFA:
-
Monounsaturated fatty acid
- PPAR:
-
Peroxisome proliferator-activated receptor
- PUFA:
-
Polyunsaturated fatty acid
- SCD1:
-
Stearic acid desaturase 1
- SM:
-
Squalene monooxygenase
- SQLE:
-
Squalene cyclooxygenase
- SRE:
-
Sterol regulating element
- SREBP:
-
Sterol regulatory element binding protein
References
Bray F, Laversanne M, Sung H, Ferlay J, Siegel RL, Soerjomataram I, Jemal A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024;74(3):229–63.
Siegel RL, Giaquinto AN, Jemal A. Cancer statistics, 2024. CA Cancer J Clin. 2024;74(1):12–49.
Onstad MA, Schmandt RE, Lu KH. Addressing the role of obesity in Endometrial Cancer Risk, Prevention, and treatment. J Clin Oncol. 2016;34(35):4225–30.
Crosbie EJ, Kitson SJ, McAlpine JN, Mukhopadhyay A, Powell ME, Singh N. Endometrial cancer. Lancet. 2022;399(10333):1412–28.
Gordhandas S, Zammarrelli WA, Rios-Doria EV, Green AK, Makker V. Current evidence-based systemic therapy for Advanced and recurrent endometrial Cancer. J Natl Compr Canc Netw. 2023;21(2):217–26.
Brooks RA, Fleming GF, Lastra RR, Lee NK, Moroney JW, Son CH, Tatebe K, Veneris JL. Current recommendations and recent progress in endometrial cancer. CA Cancer J Clin. 2019;69(4):258–79.
Bogani G, Monk BJ, Powell MA, Westin SN, Slomovitz B, Moore KN, Eskander RN, Raspagliesi F, Barretina-Ginesta MP, Colombo N, Mirza MR. Adding immunotherapy to first-line treatment of advanced and metastatic endometrial cancer. Ann Oncol. 2024;35(5):414–28.
Marín-Jiménez JA, García-Mulero S, Matías-Guiu X, Piulats JM. Facts and hopes in Immunotherapy of Endometrial Cancer. Clin Cancer Res. 2022;28(22):4849–60.
Wang L, Du ZH, Qiao JM, Gao S. Association between metabolic syndrome and endometrial cancer risk: a systematic review and meta-analysis of observational studies. Aging. 2020;12(10):9825–39.
Ding Y, Fan Y, Li X, Wang Y, Wang J, Tian L. Metabolic syndrome is an independent risk factor for time to complete remission of fertility-sparing treatment in atypical endometrial hyperplasia and early endometrial carcinoma patients. Reprod Biol Endocrinol. 2022;20(1):134.
Rosato V, Zucchetto A, Bosetti C, Dal Maso L, Montella M, Pelucchi C, Negri E, Franceschi S. La Vecchia, metabolic syndrome and endometrial cancer risk. Ann Oncol. 2011;22(4):884–9.
Jo H, Kim SI, Wang W, Seol A, Han Y, Kim J, Park IS, Lee J, Yoo J, Han KD, Song YS. Metabolic syndrome as a risk factor of Endometrial Cancer: a Nationwide Population-based Cohort Study of 2.8 million women in South Korea. Front Oncol. 2022;12:872995.
Nevadunsky NS, Van Arsdale A, Strickler HD, Moadel A, Kaur G, Levitt J, Girda E, Goldfinger M, Goldberg GL, Einstein MH. Obesity and age at diagnosis of endometrial cancer. Obstet Gynecol. 2014;124(2 Pt 1):300–6.
Zhang H, Kong W, Han C, Liu T, Li J, Song D. Correlation of metabolic factors with endometrial atypical Hyperplasia and Endometrial Cancer: Development and Assessment of a New Predictive Nomogram. Cancer Manag Res. 2021;13:7937–49.
Bokhman JV. Two pathogenetic types of endometrial carcinoma. Gynecol Oncol. 1983;15(1):10–7.
Grandi G, Perrone AM, Chiossi G, Friso S, Toss A, Sammarini M, Facchinetti F, Botticelli L, Palma F, De Iaco P. Increasing BMI is associated with both endometrioid and serous histotypes among endometrial rather than ovarian cancers: a case-to-case study. Gynecol Oncol. 2019;154(1):163–8.
Calle EE, Rodriguez C, Walker-Thurmond K, Thun MJ. Overweight, obesity, and mortality from cancer in a prospectively studied cohort of U.S. adults. N Engl J Med. 2003;348(17):1625–38.
Pavlova NN, Zhu J, Thompson CB. The hallmarks of cancer metabolism: still emerging. Cell Metab. 2022;34(3):355–77.
Martínez-Reyes I, Chandel NS. Cancer metabolism: looking forward. Nat Rev Cancer. 2021;21(10):669–80.
Xia L, Oyang L, Lin J, Tan S, Han Y, Wu N, Yi P, Tang L, Pan Q, Rao S, Liang J, Tang Y, Su M, Luo X, Yang Y, Shi Y, Wang H, Zhou Y, Liao Q. The cancer metabolic reprogramming and immune response. Mol Cancer. 2021;20(1):28.
Bergers G, Fendt SM. The metabolism of cancer cells during metastasis. Nat Rev Cancer. 2021;21(3):162–80.
Faubert B, Solmonson A, DeBerardinis RJ. Metabolic reprogramming and cancer progression. Science. 2020;368(6487):eaaw5473.
Martin-Perez M, Urdiroz-Urricelqui U, Bigas C, Benitah SA. The role of lipids in cancer progression and metastasis. Cell Metab. 2022;34(11):1675–99.
Cheng C, Geng F, Cheng X, Guo D. Lipid metabolism reprogramming and its potential targets in cancer. Cancer Commun (Lond). 2018;38(1):27.
Bian X, Liu R, Meng Y, Xing D, Xu D, Lu Z. Lipid metabolism and cancer. J Exp Med. 2021;218(1):e20201606.
Broadfield LA, Pane AA, Talebi A, Swinnen JV, Fendt SM. Lipid metabolism in cancer: new perspectives and emerging mechanisms. Dev Cell. 2021;56(10):1363–93.
Koundouros N, Poulogiannis G. Reprogramming of fatty acid metabolism in cancer. Br J Cancer. 2020;122(1):4–22.
Snaebjornsson MT, Janaki-Raman S, Schulze A. Greasing the Wheels of the Cancer machine: the role of lipid metabolism in Cancer. Cell Metab. 2020;31(1):62–76.
Hao JW, Wang J, Guo H, Zhao YY, Sun HH, Li YF, Lai XY, Zhao N, Wang X, Xie C, Hong L, Huang X, Wang HR, Li CB, Liang B, Chen S, Zhao TJ. CD36 facilitates fatty acid uptake by dynamic palmitoylation-regulated endocytosis. Nat Commun. 2020;11(1):4765.
Ruan C, Meng Y, Song H. CD36: an emerging therapeutic target for cancer and its molecular mechanisms. J Cancer Res Clin Oncol. 2022;148(7):1551–8.
Wang J, Li Y. CD36 tango in cancer: signaling pathways and functions. Theranostics. 2019;9(17):4893–908.
Yang P, Qin H, Li Y, Xiao A, Zheng E, Zeng H, Su C, Luo X, Lu Q, Liao M, Zhao L, Wei L, Varghese Z, Moorhead JF, Chen Y, Ruan XZ. CD36-mediated metabolic crosstalk between tumor cells and macrophages affects liver metastasis. Nat Commun. 2022;13(1):5782.
Feng WW, Wilkins O, Bang S, Ung M, Li J, An J, Del Genio C, Canfield K, DiRenzo J, Wells W, Gaur A, Robey RB, Guo JY, Powles RL, Sotiriou C, Pusztai L, Febbraio M, Cheng C, Kinlaw WB, Kurokawa M. CD36-Mediated metabolic rewiring of breast Cancer cells promotes resistance to HER2-Targeted therapies. Cell Rep. 2019;29(11):3405–e34205.
Kubo M, Gotoh K, Eguchi H, Kobayashi S, Iwagami Y, Tomimaru Y, Akita H, Asaoka T, Noda T, Takeda Y, Tanemura M, Mori M, Doki Y. Impact of CD36 on Chemoresistance in Pancreatic Ductal Adenocarcinoma. Ann Surg Oncol. 2020;27(2):610–9.
Su T, Huang C, Yang C, Jiang T, Su J, Chen M, Fatima S, Gong R, Hu X, Bian Z, Liu Z, Kwan HY. Apigenin inhibits STAT3/CD36 signaling axis and reduces visceral obesity. Pharmacol Res. 2020;152:104586.
Knapp P, Chabowski A, Harasiuk D, Górski J. Reversed glucose and fatty acids transporter expression in human endometrial cancer. Horm Metab Res. 2012;44(6):436–41.
McKillop IH, Girardi CA, Thompson KJ. Role of fatty acid binding proteins (FABPs) in cancer development and progression. Cell Signal. 2019;62:109336.
Tian W, Zhang W, Zhang Y, Zhu T, Hua Y, Li H, Zhang Q, Xia M. FABP4 promotes invasion and metastasis of colon cancer by regulating fatty acid transport. Cancer Cell Int. 2020;20:512.
Tang Y, Li K, Hu B, Cai Z, Li J, Tao H, Cao J. Fatty acid binding protein 5 promotes the proliferation, migration, and invasion of hepatocellular carcinoma cells by degradation of Krüppel-like factor 9 mediated by mir-889-5p via cAMP-response element binding protein. Cancer Biol Ther. 2022;23(1):424–38.
Hao J, Zhang Y, Yan X, Yan F, Sun Y, Zeng J, Waigel S, Yin Y, Fraig MM, Egilmez NK, Suttles J, Kong M, Liu S, Cleary MP, Sauter E, Li B. Circulating adipose fatty acid binding protein is a New Link underlying obesity-Associated Breast/Mammary Tumor Development. Cell Metab. 2018;28(5):689–e7055.
Li Z, Min W, Huang C, Bai S, Tang M, Zhao X. Proteomics-based approach identified differentially expressed proteins with potential roles in endometrial carcinoma. Int J Gynecol Cancer. 2010;20(1):9–15.
Li Z, Huang C, Bai S, Pan X, Zhou R, Wei Y, Zhao X. Prognostic evaluation of epidermal fatty acid-binding protein and calcyphosine, two proteins implicated in endometrial cancer using a proteomic approach. Int J Cancer. 2008;123(10):2377–83.
Wu Z, Jeong JH, Ren C, Yang L, Ding L, Li F, Jiang D, Zhu Y, Lu J. Fatty acid-binding protein 4 (FABP4) suppresses Proliferation and Migration of Endometrial Cancer cells via PI3K/Akt pathway. Onco Targets Ther. 2021;14:3929–42.
Giacomini I, Gianfanti F, Desbats MA, Orso G, Berretta M, Prayer-Galetti T, Ragazzi E, Cocetta V. Cholesterol metabolic reprogramming in Cancer and its pharmacological modulation as therapeutic strategy. Front Oncol. 2021;11:682911.
Simons K, Ehehalt R. Cholesterol, lipid rafts, and disease. J Clin Invest. 2002;110(5):597–603.
Ding X, Zhang W, Li S, Yang H. The role of cholesterol metabolism in cancer. Am J Cancer Res. 2019;9(2):219–27.
Gu J, Zhu N, Li HF, Zhao TJ, Zhang CJ, Liao DF, Qin L. Cholesterol homeostasis and cancer: a new perspective on the low-density lipoprotein receptor. Cell Oncol (Dordr). 2022;45(5):709–28.
Scully T, Ettela A, Kase N, LeRoith D, Gallagher EJ. Unregulated LDL cholesterol uptake is detrimental to breast cancer cells. Endocr Relat Cancer. 2023;30(1):e220234.
Kho PF, Amant F, Annibali D, Ashton K, Attia J, Auer PL, Beckmann MW, Black A, Brinton L, Buchanan DD, Chanock SJ, Chen C, Chen MM, Cheng THT, Cook LS, Crous-Bous M, Czene K, De Vivo I, Dennis J, Dörk T, Dowdy SC, Dunning AM, Dürst M, Easton DF, Ekici AB, Fasching PA, Fridley BL, Friedenreich CM, García-Closas M, Gaudet MM, Giles GG, Goode EL, Gorman M, Haiman CA, Hall P, Hankinson SE, Hein A, Hillemanns P, Hodgson S, Hoivik EA, Holliday EG, Hunter DJ, Jones A, Kraft P, Krakstad C, Lambrechts D, Le Marchand L, Liang X, Lindblom A, Lissowska J, Long J, Lu L, Magliocco AM, Martin L, McEvoy M, Milne RL, Mints M, Nassir R, Otton G, Palles C, Pooler L, Proietto T, Rebbeck TR, Renner SP, Risch HA, Rübner M, Runnebaum I, Sacerdote C, Sarto GE, Schumacher F, Scott RJ, Setiawan VW, Shah M, Sheng X, Shu XO, Southey MC, Tham E, Tomlinson I, Trovik J, Turman C, Tyrer JP, Van Den Berg D, Wang Z, Wentzensen N, Xia L, Xiang YB, Yang HP, Yu H, Zheng W, Webb PM, Thompson DJ, Spurdle AB, Glubb DM, O’Mara TA. Mendelian randomization analyses suggest a role for cholesterol in the development of endometrial cancer. Int J Cancer. 2021;148(2):307–19.
He W, Li Q, Li X. Acetyl-CoA regulates lipid metabolism and histone acetylation modification in cancer. Biochim Biophys Acta Rev Cancer. 2023;1878(1):188837.
Schug ZT, Peck B, Jones DT, Zhang Q, Grosskurth S, Alam IS, Goodwin LM, Smethurst E, Mason S, Blyth K, McGarry L, James D, Shanks E, Kalna G, Saunders RE, Jiang M, Howell M, Lassailly F, Thin MZ, Spencer-Dene B, Stamp G, van den Broek NJ, Mackay G, Bulusu V, Kamphorst JJ, Tardito S, Strachan D, Harris AL, Aboagye EO, Critchlow SE, Wakelam MJ, Schulze A, Gottlieb E. Acetyl-CoA synthetase 2 promotes acetate utilization and maintains cancer cell growth under metabolic stress. Cancer Cell. 2015;27(1):57–71.
Guertin DA, Wellen KE. Acetyl-CoA metabolism in cancer. Nat Rev Cancer. 2023;23(3):156–72.
Icard P, Wu Z, Fournel L, Coquerel A, Lincet H, Alifano M. ATP citrate lyase: a central metabolic enzyme in cancer. Cancer Lett. 2020;471:125–34.
Nwosu ZC, Battello N, Rothley M, Piorońska W, Sitek B, Ebert MP, Hofmann U, Sleeman J, Wölfl S, Meyer C, Megger DA, Dooley S. Liver cancer cell lines distinctly mimic the metabolic gene expression pattern of the corresponding human tumours. J Exp Clin Cancer Res. 2018;37(1):211.
Chen Y, Li K, Gong D, Zhang J, Li Q, Zhao G, Lin P. A biomarker of recurrence in breast cancer. Pathol Res Pract. 2020;216(9):153076.
Liu Q, Ge W, Wang T, Lan J, Martínez-Jarquín S, Wolfrum C, Stoffel M, Zenobi R. High-throughput single-cell Mass Spectrometry reveals abnormal lipid metabolism in pancreatic ductal adenocarcinoma. Angew Chem Int Ed Engl. 2021;60(46):24534–42.
Adorno-Cruz V, Hoffmann AD, Liu X, Dashzeveg NK, Taftaf R, Wray B, Keri RA, Liu H. ITGA2 promotes expression of ACLY and CCND1 in enhancing breast cancer stemness and metastasis. Genes Dis. 2021;8(4):493–508.
Guo H, Wang B, Xu K, Nie L, Fu Y, Wang Z, Wang Q, Wang S, Zou X. M(6)a reader HNRNPA2B1 promotes esophageal Cancer Progression via Up-Regulation of ACLY and ACC1. Front Oncol. 2020;10:553045.
Qiao C, Huang W, Chen J, Feng W, Zhang T, Wang Y, Liu D, Ji X, Xie M, Sun M, Fan D, Wu K, Xia L. IGF1-mediated HOXA13 overexpression promotes colorectal cancer metastasis through upregulating ACLY and IGF1R. Cell Death Dis. 2021;12(6):564.
Noh KH, Kang HM, Yoo W, Min Y, Kim D, Kim M, Wang S, Lim JH, Jung CR. Ubiquitination of PPAR-gamma by pVHL inhibits ACLY expression and lipid metabolism, is implicated in tumor progression. Metabolism. 2020;110:154302.
Dai M, Yang B, Chen J, Liu F, Zhou Y, Zhou Y, Xu Q, Jiang S, Zhao S, Li X, Zhou X, Yang Q, Li J, Wang Y, Zhang Z, Teng Y. Nuclear-translocation of ACLY induced by obesity-related factors enhances pyrimidine metabolism through regulating histone acetylation in endometrial cancer. Cancer Lett. 2021;513:36–49.
Kumari R, Deshmukh RS, Das S. Caspase-10 inhibits ATP-citrate lyase-mediated metabolic and epigenetic reprogramming to suppress tumorigenesis. Nat Commun. 2019;10(1):4255.
Ly CH, Lynch GS, Ryall JG. A metabolic Roadmap for somatic stem cell fate. Cell Metab. 2020;31(6):1052–67.
Zhao S, Torres A, Henry RA, Trefely S, Wallace M, Lee JV, Carrer A, Sengupta A, Campbell SL, Kuo YM, Frey AJ, Meurs N, Viola JM, Blair IA, Weljie AM, Metallo CM, Snyder NW, Andrews AJ. Wellen, ATP-Citrate lyase controls a glucose-to-acetate metabolic switch. Cell Rep. 2016;17(4):1037–52.
Comerford S.A., Huang Z, Du X, Wang Y, Cai L, Witkiewicz A.K., Walters H, Tantawy M.N., Fu A, Manning H.C., Horton J.D., Hammer R.E., McKnight S.L., Tu B.P. Acetate dependence of tumors. Cell. 2014;159(7):1591–602.
Liu M, Liu N, Wang J, Fu S, Wang X, Chen D. Acetyl-CoA Synthetase 2 as a Therapeutic Target in Tumor Metabolism. Cancers (Basel). 2022;14(12):2896.
Li Z, Liu H, He J, Wang Z, Yin Z, You G, Wang Z, Davis RE, Lin P, Bergsagel PL, Manasanch EE, Wong STC, Esnaola NF, Chang JC, Orlowski RZ, Yi Q, Yang J. Acetyl-CoA synthetase 2: a critical linkage in obesity-Induced Tumorigenesis in Myeloma. Cell Metab. 2021;33(1):78–e937.
Yu Y, Nie Q, Wang Z, Di Y, Chen X, Ren K. Targeting acetyl-CoA carboxylase 1 for cancer therapy. Front Pharmacol. 2023;14:1129010.
Hunkeler M, Hagmann A, Stuttfeld E, Chami M, Guri Y, Stahlberg H, Maier T. Structural basis for regulation of human acetyl-CoA carboxylase. Nature. 2018;558(7710):470–4.
Lally JSV, Ghoshal S, DePeralta DK, Moaven O, Wei L, Masia R, Erstad DJ, Fujiwara N, Leong V, Houde VP, Anagnostopoulos AE, Wang A, Broadfield LA, Ford RJ, Foster RA, Bates J, Sun H, Wang T, Liu H, Ray AS, Saha AK, Greenwood J, Bhat S, Harriman G, Miao W, Rocnik JL, Westlin WF, Muti P, Tsakiridis T, Harwood HJ Jr., Kapeller R, Hoshida Y, Tanabe KK, Steinberg GR, Fuchs BC. Inhibition of Acetyl-CoA carboxylase by phosphorylation or the inhibitor ND-654 suppresses lipogenesis and Hepatocellular Carcinoma. Cell Metab. 2019;29(1):174–e1825.
Byrne FL, Poon IK, Modesitt SC, Tomsig JL, Chow JD, Healy ME, Baker WD, Atkins KA, Lancaster JM, Marchion DC, Moley KH, Ravichandran KS, Slack-Davis JK, Hoehn KL. Metabolic vulnerabilities in endometrial cancer. Cancer Res. 2014;74(20):5832–45.
Modesitt SC, Hsu JY, Chowbina SR, Lawrence RT, Hoehn KL. Not all fat is equal: differential gene expression and potential therapeutic targets in subcutaneous adipose, visceral adipose, and endometrium of obese women with and without endometrial cancer. Int J Gynecol Cancer. 2012;22(5):732–41.
Fhu CW, Ali A. Fatty Acid Synthase: An Emerging Target in Cancer. Molecules. 2020;25(17):3935.
Anagnostou E, Miliaras D, Meditskou S, Grimbizis G. Immunohistochemical investigation of metabolic markers fatty acid synthase (FASN) and glucose transporter 1 (GLUT1) in normal endometrium, endometrial hyperplasia, and endometrial malignancy. Hippokratia. 2017;21(4):169–74.
Tsuji T, Yoshinaga M, Togami S, Douchi T, Nagata Y. Fatty acid synthase expression and clinicopathological findings in endometrial cancer. Acta Obstet Gynecol Scand. 2004;83(6):586–90.
Sebastiani V, Visca P, Botti C, Santeusanio G, Galati GM, Piccini V, Capezzone B, de Joannon U, Di Tondo PL, Alo. Fatty acid synthase is a marker of increased risk of recurrence in endometrial carcinoma. Gynecol Oncol. 2004;92(1):101–5.
He Y, Xu S, Qi Y, Tian J, Xu F. Long noncoding RNA SNHG25 promotes the malignancy of endometrial cancer by sponging microRNA-497-5p and increasing FASN expression. J Ovarian Res. 2021;14(1):163.
Meijuan C, Fang L, Min F, Qian W. Hypomethylated gene RAC3 induces cell proliferation and invasion by increasing FASN expression in endometrial cancer. Int J Biochem Cell Biol. 2022;150:106274.
Menendez JA, Oza BP, Atlas E, Verma VA, Mehmi I, Lupu R. Inhibition of tumor-associated fatty acid synthase activity antagonizes estradiol- and tamoxifen-induced agonist transactivation of estrogen receptor (ER) in human endometrial adenocarcinoma cells. Oncogene. 2004;23(28):4945–58.
Kubota CS, Espenshade PJ. Targeting Stearoyl-CoA desaturase in solid tumors. Cancer Res. 2022;82(9):1682–8.
Tracz-Gaszewska Z, Dobrzyn P. Stearoyl-CoA Desaturase 1 as a Therapeutic Target for the Treatment of Cancer. Cancers (Basel). 2019;11(7):948.
Tesfay L, Paul BT, Konstorum A, Deng Z, Cox AO, Lee J, Furdui CM, Hegde P, Torti FM. Torti, Stearoyl-CoA desaturase 1 protects ovarian Cancer cells from ferroptotic cell death. Cancer Res. 2019;79(20):5355–66.
Wang C, Shi M, Ji J, Cai Q, Zhao Q, Jiang J, Liu J, Zhang H, Zhu Z, Zhang J. Stearoyl-CoA desaturase 1 (SCD1) facilitates the growth and anti-ferroptosis of gastric cancer cells and predicts poor prognosis of gastric cancer. Aging. 2020;12(15):15374–91.
Luis G, Godfroid A, Nishiumi S, Cimino J, Blacher S, Maquoi E, Wery C, Collignon A, Longuespée R, Montero-Ruiz L, Dassoul I, Maloujahmoum N, Pottier C, Mazzucchelli G, Depauw E, Bellahcène A, Yoshida M, Noel A, Sounni NE. Tumor resistance to ferroptosis driven by Stearoyl-CoA Desaturase-1 (SCD1) in cancer cells and fatty acid biding Protein-4 (FABP4) in tumor microenvironment promote tumor recurrence. Redox Biol. 2021;43:102006.
Ding Z, Pan Y, Shang T, Jiang T, Lin Y, Yang C, Pang S, Cui X, Wang Y, Feng XF, Xu M, Pei M, Chen Y, Li X, Ding J, Tan Y, Wang H, Dong L, Wang L. URI alleviates tyrosine kinase inhibitors-induced ferroptosis by reprogramming lipid metabolism in p53 wild-type liver cancers. Nat Commun. 2023;14(1):6269.
Li W, Bai H, Liu S, Cao D, Wu H, Shen K, Tai Y, Yang J. Targeting stearoyl-CoA desaturase 1 to repress endometrial cancer progression. Oncotarget. 2018;9(15):12064–78.
Xu H, Zhou S, Tang Q, Xia H, Bi F. Cholesterol metabolism: new functions and therapeutic approaches in cancer. Biochim Biophys Acta Rev Cancer. 2020;1874(1):188394.
Qiu Z, Yuan W, Chen T, Zhou C, Liu C, Huang Y, Han D, Huang Q. HMGCR positively regulated the growth and migration of glioblastoma cells. Gene. 2016;576(1 Pt 1):22–7.
Chushi L, Wei W, Kangkang X, Yongzeng F, Ning X, Xiaolei C. HMGCR is up-regulated in gastric cancer and promotes the growth and migration of the cancer cells. Gene. 2016;587(1):42–7.
Orho-Melander M, Hindy G, Borgquist S, Schulz CA, Manjer J, Melander O, Stocks T. Blood lipid genetic scores, the HMGCR gene and cancer risk: a mendelian randomization study. Int J Epidemiol. 2018;47(2):495–505.
Bjarnadottir O, Feldt M, Inasu M, Bendahl PO, Elebro K, Kimbung S, Borgquist S. Statin use, HMGCR expression, and breast cancer survival - the Malmö Diet and Cancer Study. Sci Rep. 2020;10(1):558.
Che L, Chi W, Qiao Y, Zhang J, Song X, Liu Y, Li L, Jia J, Pilo MG, Wang J, Cigliano A, Ma Z, Kuang W, Tang Z, Zhang Z, Shui G, Ribback S, Dombrowski F, Evert M, Pascale RM, Cossu C, Pes GM, Osborne TF, Calvisi DF, Chen X, Chen L. Cholesterol biosynthesis supports the growth of hepatocarcinoma lesions depleted of fatty acid synthase in mice and humans. Gut. 2020;69(1):177–86.
Yarmolinsky J, Bull CJ, Vincent EE, Robinson J, Walther A, Smith GD, Lewis SJ, Relton CL, Martin RM. Association between genetically proxied inhibition of HMG-CoA reductase and epithelial ovarian Cancer. JAMA. 2020;323(7):646–55.
Sun L, Ding H, Jia Y, Shi M, Guo D, Yang P, Wang Y, Liu F, Zhang Y, Zhu Z. Associations of genetically proxied inhibition of HMG-CoA reductase, NPC1L1, and PCSK9 with breast cancer and prostate cancer. Breast Cancer Res. 2022;24(1):12.
Wang G, Liu X, Wang D, Sun M, Yang Q. Identification and development of subtypes with poor prognosis in Pan-gynecological Cancer based on Gene expression in the glycolysis-cholesterol synthesis Axis. Front Oncol. 2021;11:636565.
Zhang HY, Li HM, Yu Z, Yu XY, Guo K. Expression and significance of squalene epoxidase in squamous lung cancerous tissues and pericarcinoma tissues. Thorac Cancer. 2014;5(4):275–80.
Kim NI, Park MH, Kweon SS, Cho N, Lee JS. Squalene epoxidase expression is associated with breast tumor progression and with a poor prognosis in breast cancer. Oncol Lett. 2021;21(4):259.
Li C, Wang Y, Liu D, Wong CC, Coker OO, Zhang X, Liu C, Zhou Y, Liu Y, Kang W, To KF, Sung JJ, Yu J. Squalene epoxidase drives cancer cell proliferation and promotes gut dysbiosis to accelerate colorectal carcinogenesis. Gut. 2022;71(11):2253–65.
Jiang H, Tang E, Chen Y, Liu H, Zhao Y, Lin M, He L. Squalene synthase predicts poor prognosis in stage I-III colon adenocarcinoma and synergizes squalene epoxidase to promote tumor progression. Cancer Sci. 2022;113(3):971–85.
Kalogirou C, Linxweiler J, Schmucker P, Snaebjornsson MT, Schmitz W, Wach S, Krebs M, Hartmann E, Puhr M, Müller A, Spahn M, Seitz AK, Frank T, Marouf H, Büchel G, Eckstein M, Kübler H, Eilers M, Saar M, Junker K, Röhrig F, Kneitz B, Rosenfeldt MT, Schulze A. MiR-205-driven downregulation of cholesterol biosynthesis through SQLE-inhibition identifies therapeutic vulnerability in aggressive prostate cancer. Nat Commun. 2021;12(1):5066.
Ma L, Huang W, Liang X, Bai G, Wang X, Jiang H, Xin Y, Hu L, Chen X, Liu C. Inhibition of squalene epoxidase linking with PI3K/AKT signaling pathway suppresses endometrial cancer. Cancer Sci. 2023;114(9):3595–607.
Coates HW, Nguyen TB, Du X, Olzomer EM, Farrell R, Byrne FL, Yang H, Brown AJ. The constitutively active form of a key cholesterol synthesis enzyme is lipid droplet-localized and upregulated in endometrial cancer tissues. J Biol Chem. 2024;300(5):107232.
Luo J, Yang H, Song BL. Mechanisms and regulation of cholesterol homeostasis. Nat Rev Mol Cell Biol. 2020;21(4):225–45.
Bhattacharjee P, Rutland N, Iyer MR. Targeting sterol O-Acyltransferase/Acyl-CoA:cholesterol acyltransferase (ACAT): a perspective on small-molecule inhibitors and their therapeutic potential. J Med Chem. 2022;65(24):16062–98.
Eckhardt C, Sbiera I, Krebs M, Sbiera S, Spahn M, Kneitz B, Joniau S, Fassnacht M, Kübler H, Weigand I, Kroiss M. High expression of Sterol-O-Acyl transferase 1 (SOAT1), an enzyme involved in cholesterol metabolism, is associated with earlier biochemical recurrence in high risk prostate cancer. Prostate Cancer Prostatic Dis. 2022;25(3):484–90.
Omsjø IH, Norum KR. Cholesterol esterification in human secretory endometrium and in endometrial cancer tissue. Demonstration of microsomal acyl-CoA-cholesterol acyl-transferase (ACAT) activity. Acta Obstet Gynecol Scand. 1985;64(6):473–6.
Chen Y, Lee K, Liang Y, Qin S, Zhu Y, Liu J, Yao S. A cholesterol homeostasis-related gene signature predicts prognosis of Endometrial Cancer and correlates with Immune Infiltration. Front Genet. 2021;12:763537.
Ma Y, Temkin SM, Hawkridge AM, Guo C, Wang W, Wang XY, Fang X. Fatty acid oxidation: an emerging facet of metabolic transformation in cancer. Cancer Lett. 2018;435:92–100.
Quan J, Cheng C, Tan Y, Jiang N, Liao C, Liao W, Cao Y, Luo X. Acyl-CoA synthetase long-chain 3-mediated fatty acid oxidation is required for TGFβ1-induced epithelial-mesenchymal transition and metastasis of colorectal carcinoma. Int J Biol Sci. 2022;18(6):2484–96.
Tang M, Dong X, Xiao L, Tan Z, Luo X, Yang L, Li W, Shi F, Li Y, Zhao L, Liu N, Du Q, Xie L, Hu J, Weng X, Fan J, Zhou J, Gao Q, Wu W, Zhang X, Liao W, Bode AM, Cao Y. CPT1A-mediated fatty acid oxidation promotes cell proliferation via nucleoside metabolism in nasopharyngeal carcinoma. Cell Death Dis. 2022;13(4):331.
Zhou Y, Li Y, Chen G, Guo X, Gao X, Meng J, Xu Y, Zhou N, Zhang B, Zhou X. ACSL1-Mediated fatty acid β-Oxidation enhances metastasis and proliferation in Endometrial Cancer. Front Biosci (Landmark Ed). 2024;29(2):66.
Yu Y, Sun X, Chen F, Liu M. Genetic alteration, prognostic and immunological role of Acyl-CoA synthetase Long-Chain Family Member 4 in a Pan-cancer Analysis. Front Genet. 2022;13:812674.
Duan Y, Gong K, Xu S, Zhang F, Meng X, Han J. Regulation of cholesterol homeostasis in health and diseases: from mechanisms to targeted therapeutics. Signal Transduct Target Ther. 2022;7(1):265.
Sharma B, Agnihotri N. Role of cholesterol homeostasis and its efflux pathways in cancer progression. J Steroid Biochem Mol Biol. 2019;191:105377.
Antmen E, Yalaza ŞC, Canacankatan N, Aytan H, Tuncel F, Erden S, Ertürk. Efficacy of ABCA1 transporter proteins in patients with endometrial Cancer: an in Vitro Study. Turk J Pharm Sci. 2024;21(3):219–23.
Wen Q, Xie X, Chen C, Wen B, Liu Y, Zhou J, Lin X, Jin H, Shi K. Lipid reprogramming induced by the NNMT-ABCA1 axis enhanced membrane fluidity to promote endometrial cancer progression. Aging. 2023;15(21):11860–74.
Shimano H, Sato R. SREBP-regulated lipid metabolism: convergent physiology - divergent pathophysiology. Nat Rev Endocrinol. 2017;13(12):710–30.
DeBose-Boyd RA, Ye J. SREBPs in lipid metabolism, insulin signaling, and Beyond. Trends Biochem Sci. 2018;43(5):358–68.
Shao W, Espenshade PJ. Expanding roles for SREBP in metabolism. Cell Metab. 2012;16(4):414–9.
Zhao Q, Lin X, Wang G. Targeting SREBP-1-Mediated lipogenesis as potential strategies for Cancer. Front Oncol. 2022;12:952371.
Xue L, Qi H, Zhang H, Ding L, Huang Q, Zhao D, Wu BJ, Li X. Targeting SREBP-2-Regulated Mevalonate Metabolism for Cancer Therapy. Front Oncol. 2020;10:1510.
Shafiee MN, Mongan N, Seedhouse C, Chapman C, Deen S, Abu J, Atiomo W. Sterol regulatory element binding protein-1 (SREBP1) gene expression is similarly increased in polycystic ovary syndrome and endometrial cancer. Acta Obstet Gynecol Scand. 2017;96(5):556–62.
Li W, Tai Y, Zhou J, Gu W, Bai Z, Zhou T, Zhong Z, McCue PA, Sang N, Ji JY, Kong B, Jiang J, Wang C. Repression of endometrial tumor growth by targeting SREBP1 and lipogenesis. Cell Cycle. 2012;11(12):2348–58.
Qiu CP, Lv QT, Dongol S, Wang C, Jiang J. Single nucleotide polymorphism of SREBF-1 gene associated with an increased risk of endometrial cancer in Chinese women. PLoS ONE. 2014;9(3):e90491.
Lin CY, Gustafsson J. Targeting liver X receptors in cancer therapeutics. Nat Rev Cancer. 2015;15(4):216–24.
Repa JJ, Liang G, Ou J, Bashmakov Y, Lobaccaro JM, Shimomura I, Shan B, Brown MS, Goldstein JL, Mangelsdorf DJ. Regulation of mouse sterol regulatory element-binding protein-1c gene (SREBP-1c) by oxysterol receptors, LXRalpha and LXRbeta. Genes Dev. 2000;14(22):2819–30.
Zhang X, Liu J, Su W, Wu J, Wang C, Kong X, Gustafsson J, Ding J, Ma X, Guan Y. Liver X receptor activation increases hepatic fatty acid desaturation by the induction of SCD1 expression through an LXRα-SREBP1c-dependent mechanism. J Diabetes. 2014;6(3):212–20.
Zelcer N, Hong C, Boyadjian R, Tontonoz P. LXR regulates cholesterol uptake through Idol-dependent ubiquitination of the LDL receptor. Science. 2009;325(5936):100–4.
Wang B, Tontonoz P. Liver X receptors in lipid signalling and membrane homeostasis. Nat Rev Endocrinol. 2018;14(8):452–63.
Fang F, Li D, Zhao L, Li Y, Zhang T, Cui B. Expression of NR1H3 in endometrial carcinoma and its effect on the proliferation of Ishikawa cells in vitro. Onco Targets Ther. 2019;12:685–97.
Abdalkareem Jasim S, Kzar HH, Haider Hamad M, Ahmad I, Al-Gazally ME, Ziyadullaev S, Sivaraman R, Abed Jawad M, Thaeer Hammid A, Oudaha KH, Karampoor S, Mirzaei R. The emerging role of 27-hydroxycholesterol in cancer development and progression: an update. Int Immunopharmacol. 2022;110:109074.
Gibson DA, Collins F, Cousins FL, Esnal Zufiaurre A, Saunders PTK. The impact of 27-hydroxycholesterol on endometrial cancer proliferation. Endocr Relat Cancer. 2018;25(4):381–91.
Christofides A, Konstantinidou E, Jani C, Boussiotis VA. The role of peroxisome proliferator-activated receptors (PPAR) in immune responses. Metabolism. 2021;114:154338.
Mirza AZ, Althagafi II, Shamshad H. Role of PPAR receptor in different diseases and their ligands: physiological importance and clinical implications. Eur J Med Chem. 2019;166:502–13.
Wagner N, Wagner KD. Peroxisome Proliferator-Activated Receptors and the Hallmarks of Cancer. Cells. 2022;11(15):2432.
Tong BJ, Tan J, Tajeda L, Das SK, Chapman JA, DuBois RN, Dey SK. Heightened expression of cyclooxygenase-2 and peroxisome proliferator-activated receptor-delta in human endometrial adenocarcinoma. Neoplasia. 2000;2(6):483–90.
Knapp P, Chabowski A, Błachnio-Zabielska A, Jarząbek K, Wołczyński S. Altered peroxisome-proliferator activated receptors expression in human endometrial cancer. PPAR Res. 2012;2012:471524.
Zhang G, Hou X, Gao S. Stimulation of peroxisome proliferator-activated receptor γ inhibits estrogen receptor α transcriptional activity in endometrial carcinoma cells. Oncol Rep. 2015;33(3):1227–34.
Ota K, Ito K, Suzuki T, Saito S, Tamura M, Hayashi S, Okamura K, Sasano H, Yaegashi N. Peroxisome proliferator-activated receptor gamma and growth inhibition by its ligands in uterine endometrial carcinoma. Clin Cancer Res. 2006;12(14 Pt 1):4200–8.
Nickkho-Amiry M, McVey R, Holland C. Peroxisome proliferator-activated receptors modulate proliferation and angiogenesis in human endometrial carcinoma. Mol Cancer Res. 2012;10(3):441–53.
Lu YU, Miyamoto T, Takeuchi H, Tsunoda F, Tanaka N, Shiozawa T. PPARα activator irbesartan suppresses the proliferation of endometrial carcinoma cells via SREBP1 and ARID1A. Oncol Res. 2023;31(3):239–53.
Ma JJ, Monsivais D, Dyson MT, Coon JS, Malpani S, Ono M, Zhao H, Xin H, Pavone ME, Kim JJ, Chakravarti D, Bulun SE. Ligand-activated peroxisome proliferator-activated receptor β/δ modulates human endometrial cancer cell survival. Horm Cancer. 2013;4(6):358–70.
Schcolnik-Cabrera A, Chávez-Blanco A, Domínguez-Gómez G, Taja-Chayeb L, Morales-Barcenas R, Trejo-Becerril C, Perez-Cardenas E, Gonzalez-Fierro A. Dueñas-González, Orlistat as a FASN inhibitor and multitargeted agent for cancer therapy. Expert Opin Investig Drugs. 2018;27(5):475–89.
Wysham WZ, Roque DR, Han J, Zhang L, Guo H, Gehrig PA, Zhou C, Bae-Jump VL. Effects of fatty acid synthase inhibition by Orlistat on Proliferation of Endometrial Cancer Cell lines. Target Oncol. 2016;11(6):763–9.
Falchook G, Infante J, Arkenau HT, Patel MR, Dean E, Borazanci E, Brenner A, Cook N, Lopez J, Pant S, Frankel A, Schmid P, Moore K, McCulloch W, Grimmer K, O’Farrell M, Kemble G, Burris H. First-in-human study of the safety, pharmacokinetics, and pharmacodynamics of first-in-class fatty acid synthase inhibitor TVB-2640 alone and with a taxane in advanced tumors. EClinicalMedicine. 2021;34:100797.
Sun Q, Xing X, Wang H, Wan K, Fan R, Liu C, Wang Y, Wu W, Wang Y, Wang R. SCD1 is the critical signaling hub to mediate metabolic diseases: mechanism and the development of its inhibitors. Biomed Pharmacother. 2024;170:115586.
Wang KH, Liu CH, Ding DC. Statins as Repurposed Drugs in Gynecological Cancer: A Review. Int J Mol Sci. 2022;23(22):13937.
Jiang W, Hu JW, He XR, Jin WL, He XY. Statins: a repurposed drug to fight cancer. J Exp Clin Cancer Res. 2021;40(1):241.
Schointuch MN, Gilliam TP, Stine JE, Han X, Zhou C, Gehrig PA, Kim K, Bae-Jump VL. Simvastatin, an HMG-CoA reductase inhibitor, exhibits anti-metastatic and anti-tumorigenic effects in endometrial cancer. Gynecol Oncol. 2014;134(2):346–55.
Zhou D, Wu Q, Qiu H, Li M, Ji Y. Simvastatin inhibits endometrial cancer malignant behaviors by suppressing RAS/Mitogen-Activated Protein Kinase (MAPK) pathway-mediated Reactive Oxygen Species (ROS) and ferroptosis. Evid Based Complement Alternat Med. 2022;2022:6177477.
Kim JS, Turbov J, Rosales R, Thaete LG, Rodriguez GC. Combination simvastatin and metformin synergistically inhibits endometrial cancer cell growth. Gynecol Oncol. 2019;154(2):432–40.
Cai Y, Zhao F. Fluvastatin suppresses the proliferation, invasion, and migration and promotes the apoptosis of endometrial cancer cells by upregulating Sirtuin 6 (SIRT6). Bioengineered. 2021;12(2):12509–20.
Xu R, Song J, Ruze R, Chen Y, Yin X, Wang C, Zhao Y. SQLE promotes pancreatic cancer growth by attenuating ER stress and activating lipid rafts-regulated Src/PI3K/Akt signaling pathway. Cell Death Dis. 2023;14(8):497.
Sun H, Li L, Li W, Yang F, Zhang Z, Liu Z, Du W. p53 transcriptionally regulates SQLE to repress cholesterol synthesis and tumor growth. EMBO Rep. 2021;22(10):e52537.
Du Y, Rokavec M, Hermeking H. Squalene epoxidase/SQLE is a candidate target for treatment of colorectal cancers with p53 mutation and elevated c-MYC expression. Int J Biol Sci. 2023;19(13):4103–22.
Gao S, Shi Z, Li X, Li W, Wang Y, Liu Z, Jiang J. Fatostatin suppresses growth and enhances apoptosis by blocking SREBP-regulated metabolic pathways in endometrial carcinoma. Oncol Rep. 2018;39(4):1919–29.
Yao L, Chen S, Li W. Fatostatin inhibits the development of endometrial carcinoma in endometrial carcinoma cells and a xenograft model by targeting lipid metabolism. Arch Biochem Biophys. 2020;684:108327.
Wang Q, Hu R, Li W, Tai Y, Gu W, Das BC, Yang F, Ji J, Wang C, Zhou J. BF175 inhibits endometrial carcinoma through SREBP-regulated metabolic pathways in vitro. Mol Cell Endocrinol. 2021;523:111135.
Shi Z, Zhou Q, Gao S, Li W, Li X, Liu Z, Jin P, Jiang J. Silibinin inhibits endometrial carcinoma via blocking pathways of STAT3 activation and SREBP1-mediated lipid accumulation. Life Sci. 2019;217:70–80.
Sun J, Zheng Z, Chen Q, Pan Y, Quan M, Dai Y. Fenofibrate potentiates chemosensitivity to human breast cancer cells by modulating apoptosis via AKT/NF-κB pathway. Onco Targets Ther. 2019;12:773–83.
Luty M, Piwowarczyk K, Łabędź-Masłowska A, Wróbel T, Szczygieł M, Catapano J, Drabik G, Ryszawy D, Kędracka-Krok S, Madeja Z, Siedlar M, Elas M, Czyż J. Fenofibrate Augments the Sensitivity of Drug-Resistant Prostate Cancer Cells to Docetaxel. Cancers (Basel). 2019;11(1):77.
Wang MS, Han QS, Jia ZR, Chen CS, Qiao C, Liu QQ, Zhang YM, Wang KW, Wang J, Xiao K, Ding XS. PPARα agonist fenofibrate relieves acquired resistance to gefitinib in non-small cell lung cancer by promoting apoptosis via PPARα/AMPK/AKT/FoxO1 pathway. Acta Pharmacol Sin. 2022;43(1):167–76.
Holland CM, Saidi SA, Evans AL, Sharkey AM, Latimer JA, Crawford RA, Charnock-Jones DS, Print CG, Smith SK. Transcriptome analysis of endometrial cancer identifies peroxisome proliferator-activated receptors as potential therapeutic targets. Mol Cancer Ther. 2004;3(8):993–1001.
Saidi SA, Holland CM, Charnock-Jones DS, Smith SK. In vitro and in vivo effects of the PPAR-alpha agonists fenofibrate and retinoic acid in endometrial cancer. Mol Cancer. 2006;5:13.
Kumari GK, Kiran A, Krishnamurthy PT. Preliminary evaluation on the beneficial effects of pioglitazone in the treatment of endometrial cancer. Med Oncol. 2021;38(6):71.
Acknowledgements
Relevant clinical trial data were downloaded from https://clinicaltrials.gov/. All figures in the manuscript were created by Figdraw (https://www.figdraw.com/#/).
Funding
This work was supported by the National Key Research & Development Program of China (grant ID: 2021YFC2701402).
Author information
Authors and Affiliations
Contributions
Xiangyu Wang contributed to manuscript writing and visualization. Yinuo Li contributed to manuscript revision and editing. Xin Hou and Jingfang Li contributed to literature collection. Xiangyi Ma contributed to conceptualization and supervision. All authors listed have approved this manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wang, X., Li, Y., Hou, X. et al. Lipid metabolism reprogramming in endometrial cancer: biological functions and therapeutic implications. Cell Commun Signal 22, 436 (2024). https://doi.org/10.1186/s12964-024-01792-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12964-024-01792-7