
Abstract
Background
Southeast Asia is well known as a hotspot of biodiversity. However, very little is known about cave-dwelling hematophagous insects that are medically important. Taxonomic knowledge and ecology of phlebotomine sand flies are very poorly studied in Laos, as well as in other countries in the region. Herein, we report species diversity data and some notes on the ecology of the detected species from these karstic limestone areas of Laos.
Methods
Phlebotomine sand flies were collected using Centers for Disease Control and Prevention (CDC) light traps from limestone cave locations in three districts of Vientiane Province, Laos. Both morphological and molecular techniques were used for sand fly identification. Species diversity and abundance were analyzed according to sites, locations, collection seasons, and trapping positions.
Results
A total of 6564 sand flies, of which 5038 were females and 1526 were males, were morphologically identified into 20 species belonging to five genera (Chinius, Idiophlebotomus, Phlebotomus, Sergentomyia, and Grassomyia). The most abundant species were Chinius eunicegalatiae, Phlebotomus stantoni, Sergentomyia hivernus, Se. siamensis, and Idiophlebotomus longiforceps. Cytochrome b analysis results supported the morphological identification and revealed that Se. siamensis was separated from other members of the Se. barraudi group. Two new species, Se. dvoraki n. sp. and Se. marolii n. sp., were described. Sand fly density was generally high except in a cave in Vangvieng, with species richness ranging from 14 to 18 across different caves. Outside caves had higher species richness (R = 20) and diversity (H = 2.50) than cave entrances (R = 18, H = 2.41) and interiors (R = 16, H = 2.13). Seasonal variations showed high sand fly density in Feung and Hinheup during both dry and rainy seasons, while Vangvieng had a notable decrease in density during the dry season (D = 6.29).
Conclusions
This study revealed that the diversity of phlebotomine sand fly fauna in Laos, particularly in karstic limestone areas, is greater than previously known. However, the taxonomic status of many species in Laos, as well as Southeast Asia, still needs more in-depth study using both morphological characters and molecular methods. Many species could be found from inside, at the entrance, and outside of caves, indicating a wide range of host-seeking behavior or possible natural breeding in the karstic cave areas.
Graphical abstract

Similar content being viewed by others


Background
Vector-borne diseases (VBDs) are the second most common cause of emerging infectious disease (EID) events after zoonotic pathogens [1]. Over the past few decades, there has been significant emergence and re-emergence of several VBDs, including malaria, leishmaniasis, dengue, yellow fever, Zika, chikungunya, and plague [2,3,4]. The emergence of new and resurgent known vector-borne pathogens can be associated with several factors, including adaptation to and changes in microorganisms, habitat, globalization, tourism, and travel [5, 6].
Karstic areas and cave ecosystems are specific environmental conditions that provide suitable places for insects and harbor many different opportunistic pathogens, such as viruses, bacteria, and fungi [7, 8]. Many of these pathogens infect cave-dwelling vertebrates, especially bats [9,10,11,12,13]. Some of these viruses can be transmitted from one vertebrate host to another via hematophagous arthropod vectors (e.g., mosquitoes, sand flies, bat flies, and biting midges) [14,15,16]. Humans and other animals that visit these environments are at risk of exposure to arthropod vectors in caves and in surrounding areas. Additionally, visitors themselves may inadvertently introduce pathogens to these areas where vectors are present. This introduces the possibility of pathogens being introduced to cave ecosystems through human activities, highlighting the need for careful consideration and monitoring of potential disease transmission pathways in these environments.
In Southeast Asia (SE Asia), cave visits are increasingly popular for many reasons, whether by local populations looking for resources, for economic purposes such as ecotourism, or for spiritual purposes (cave-dwelling monks). Therefore, such growing human incursions into caves may increase the risk of exposure and spillover of emerging pathogens that circulate among cave-dwelling vertebrates and vectors. SE Asia is a well-known biodiversity hotspot [17]. However, very little is known about cave-dwelling hematophagous insects that are medically important, such as mosquitoes (Culicidae), sand flies (Psychodidae), and biting midges (Ceratopogonidae). Taxonomic knowledge of sand flies is very poor among countries in SE Asia. The last revision of sand flies in this region was made by Lewis in 1978 [18]. The first autochthonous human case of leishmaniasis was reported in Thailand in 1996, followed by several cases caused by Leishmania (Mundinia) martiniquensis (formerly identified as Leishmania siamensis) [19,20,21]. According to these results, Thailand is the only country in the region with advanced studies on sand flies. Leishmania DNA has been detected in the Sergentomyia barraudi and Sergentomyia iyengari sensu Raynal 1936 groups, notably including Sergentomyia khawi, which was probably misidentified as Sergentomyia gemmea [22]. However, polymerase chain reaction (PCR) alone cannot conclusively prove whether an insect can transmit pathogens. Additional methods are necessary to confirm the insect’s role as a disease vector, as PCR detects pathogen genetic material but does not demonstrate transmission capability.
In Laos, before this study, only one cavernicolous species of sand fly belonging to the genus Chinius Leng had been reported, in 2010 [23], while two other species of Phlebotomus and five species of Sergentomyia were reported in 1962 by Quate [24]. In 2019, one more species of Sergentomyia from Laos was reported [22]. To our knowledge, there are no data on the diversity of cavernicolous or karstic insects of medical or veterinary interest in Laos. Thus, to fill this gap, we investigated sand flies in karstic areas of Vientiane Province in Laos in 2019, and two more new species were described [25]. Here, we report additional data on species diversity and some notes on the ecology of the detected species in karstic limestone areas of Laos.
Methods
Study sites
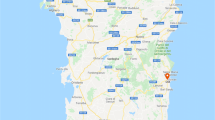
A total of seven caves in three districts with karstic limestone mountains were selected as study sites: Fueng district (18°30′N, 101°59′E), Hinheup district (18°46′N, 102°16′E), and Vangvieng district (18°58′N, 102°20′E) (Fig. 1). Among the three districts, Vangvieng is well known for cave exploration, with increasing numbers of tourists visiting each year. It is one of the top tourism destinations in Laos. In Fueng, three locations where large caves were located in each of three isolated limestones were selected for our sampling, namely, Tham Phanokkok (PNK), Tham Nam (TN), and Tham Pha (TP); in Hinheup, two caves in one isolated limestone were selected, namely, Tham Yao (TY) and Tham Phaluesy (PLS); and in Vangvieng, one large cave, Tham Nang Oau Khiem (NOK), and one touristic site in karstic areas with no large cave, namely, Angluang (AL), in a long and large limestone were selected (Additional file 1: Table S1).

Study sites and sand fly species composition from karstic limestones in three districts of Vientiane Province. Red dots represent sampling locations
Sand fly collection
To gather baseline data on the diversity and ecology of sand flies in karstic caves and peri-karstic caves within limestone areas, we analyzed sand flies collected over a total of 202 trap-nights. Sampling occurred twice in 2019 across seven locations in three selected districts. Specifically, sand flies were collected over 72 trap-nights in Feung (21 in May and 51 in October), 40 trap-nights in Hinheup (13 in February and 27 in September), and 90 trap-nights in Vangvieng (42 in January and 48 in July). Due to limited resources and logistical constraints, the collection effort was not equal across locations. Collection times coincided with Laos’s dry season (approximately November to May) and rainy season (approximately June to October). The standard Centers for Disease Control and Prevention light traps (CDC traps) were used by setting between 04:00–06:00 p.m. and 07:00–08:00 a.m. on the next day. Sand flies were collected from three main positions—outside, at the entrance, and inside caves (as described in Additional file 1: Table S1). All insects from the traps were frozen at −20 °C for 20–30 min, after which the sand flies were sorted and counted by sex. Half of the sand fly specimens were randomly stored at −20 °C for future pathogen detection, and the remaining specimens were stored in 70–90% ethanol and then transported to the Institut Pasteur du Laos (IP-Laos) laboratory in Vientiane Capital for morphological identification.
Specimen preparation and identification
Sand fly specimens were mounted whole-body (in toto) on slides using polyvinyl alcohol (PVA) media. To study their genetics, some sand fly specimens were prepared as follows: The head, wings, and genitalia were cut under a stereomicroscope using sterile needles. The thorax was then transferred individually into a tube labeled with the same identification number as the slide and stored at −20 °C until the molecular study was performed. The head, wings, and genitalia were mounted immediately on a slide using PVA media. All slides were morphologically identified using the dichotomous keys of Lewis and other related original description references [18, 22,23,24,25,26,27,28,29]. Males of the genus Sergentomyia were not identified at the species level due to the high similarity of the male morphology within this genus. Similarly, all female specimens with poor-quality slide mounting were not identified at the species level. For the descriptions of the new species, the consensual terminology of Galati et al. [30] was used.
Molecular study
Thorax samples were homogenized for 2–5 min using a TissueLyser II system (Qiagen) with 0.5 ml of 1× phosphate-buffered saline (PBS) and Lysing Matrix E zirconium beads (MP Biomedicals). A NucleoSpin®8 (Macherey–Nagel) extraction kit was used to extract 100 µl of the supernatant, following the manufacturer’s protocol. All PCR amplifications were carried out in a 50 μl volume containing 5 µl of extracted DNA and 45 μl of PCR Master Mix (Promega) containing 50 pmol of each primer targeting the cytochrome b gene (Cyt-b), i.e., C3B-PDR (5′-CAYATTCAACCWGAATGATA-3′) and N1N-PDR (5′-GGTAYWTTGCCTCGAWTTCGWTATGA-3′), according to previously published conditions [31]. Sequencing reactions were performed using the BigDye Terminator v1.1 cycle sequencing kit (Applied Biosystems) at the IP-Laos.
Data analysis
The sand fly density (D) was estimated as the number of sand flies collected per trap per night (trap-night). For specimens identified to species level, species richness (R) was determined as the number of sand fly species collected. All the statistical analyses were performed using R software (https://www.r-project.org/). The alpha diversity of the communities, the Shannon index (H) [32], was calculated, and the Kruskal‒Wallis test was subsequently used to compare the Shannon index between sites via the Rstatix package. To investigate the similarity between sites, trapping positions, and seasons in terms of species composition and density (beta diversity), the Bray‒Curtis index between sites was calculated for each trap-night. Principle coordinate analysis (PCoA) was used to display the beta diversity indices for sand flies among sites and locations. Permutational multivariate analysis of variance (PERMANOVA) was used to compare the beta diversity indices using the vegan package.
Newly generated sequences from this study were aligned with Cyt-b sequences available in GenBank and used for genetic analysis (Additional file 2: Table S2). Genetic distances were calculated within species and between species groups using the Tamura–Nei model. A maximum likelihood (ML) tree was constructed in MEGA 11 [33] using the substitution models selected by MODELTEST [34].
Results
Sand fly morphological species identification and composition
A total of 6564 sand flies were morphologically identified, of which 5038 (76.8%) were females and 1526 (23.3%) were males. The sand flies were classified into 20 species belonging to five genera: Chinius (one species): Ch. eunicegalatiae, 1555 (23.7%); Idiophlebotomus (one species): Id. longiforceps, 351 (5.4%); Phlebotomus (six species): Ph. barguesae, three (0.1%); Ph. breyi, 90 (1.4%); Ph. kiangsuensis, 51 (0.8%); Ph. mascomai, 307 (4.7%); Ph. sinxayarami, 338 (5.2%); Ph. stantoni, 606 (9.2%), and unclassified as mounting slides were not clear, Phlebotomus spp., 52 (0.8%); Sergentomyia (11 species): Se. anodontis, 337 (5.1%); Se. siamensis, 462 (7.0%); Se. brevicaulis-like, five (0.08%); Se. hivernus, 536 (8.1%); Se. khawi, 416 (6.3%); Se. perturbans, four (0.1%); Se. phasukae, 33 (0.5%); Se. rudnicki, 164 (2.5%); Se. sylvatica, 110 (1.7%); Se. dvoraki n. sp., 184 (2.8%); Se. marolii n. sp., two (0.04%); Sergentomyia spp., 898 (13.7%); and Grassomyia (one species): Gr. indica, 60 (0.1%); (Fig. 1 and Table 1).
Regarding the taxonomy of sand flies within the genus Sergentomyia in this study, specimens were classified as Se. siamensis based on the following characters: around 54 comb-like teeth (posterior teeth) with two rows of anterior teeth (fore-teeth) of about 22 and six on the cibarium, short antenna flagellomer1 (f1) less than 190 µm, and its length about half of the length of proboscis, ovoid and smooth spermathecae (Fig. 2A, D, G, and J); as Se. brevicaulis-like for those that have long antennae f1, extending beyond the tip of the proboscis with about 50 posterior comb-like teeth on cibarium (Fig. 2B, E, H, and K); and as Se. rudnicki for those that have long antennae f1, longer than 250 µm, extending beyond the tip of the proboscis, with about 80 posterior comb-like teeth on the cibarium and two rows of anterior teeth of more than 30 on each row, with oblong and annulate spermathecae (Fig. 2C, F, I, and K). Main morphological characteristics, cibarial teeth, and spermathecae for the diagnosis of Se. khawi, Se. hivernus, Se. dvoraki n. sp., Se. marolii n. sp., Se. phasukae, Se. perturbans, and Se. sylvatica are shown in Figs. 3 and 4. Descriptions of Se. dvoraki n. sp. and Se. marolii n. sp. are provided below. Comments on the taxonomy of the sand flies in this study are provided in the Discussion section.

Female morphological characteristics of Se. siamensis, Se. brevicaulis-like, and Se. rudnicki found in this study. Sergentomyia siamensis: head with short flagellomere 1 (A), cibarium with about 54 teeth (D), pharyngeal teeth long (G), smooth and ovoid spermathecae (J); Se. brevicaulis-like teeth: head with long flagellomere 1 (B), cibarium with about 50 comb-like teeth (E), pharyngeal teeth long (H), oblong and wrinkled spermathecae (K); and Se. rudnicki: head with long flagellomere 1 (C), cibarium with about 80 comb-like teeth (F), pharyngeal teeth long (I), and oblong and wrinkled spermathecae (L)

Female morphological characteristics of Sergentomyia. Sergentomyia khawi: cibarium with three rows of vertical teeth (A), smooth spermathecae and wide individual duct (D); Se. hivenus: cibarium (B) and tubular spermathecae (E); Se. dvoraki n. sp.: cibarium of paratype (C), spermathecae with long and narrow individual ducts (F); Se. gemmea (holotype): red arrow points to the spur of the ascoid (I); Se. phasukae: cibarial teeth, no pigment patch (G), spermathecae (J); and Se. sylvatica: cibarium teeth (H), and spermathecae (K)

Sergentomyia perturbans: cibarium (A), pharynx unarmed (D), and mesofurca strongly pigmented (F); and Se. marolii n. sp.: cibarium (B), pharynx armed with teeth (E), and mesofurca strongly pigmented (C)
Phylogenetic analysis and genetic distance
A total of 129 samples were successfully amplified, and the following Cyt-b sequences were obtained: Ch. eunicegalatiae (four specimens), Id. longiforceps (10), Ph. breyi (10), Ph. kiangsuensis (one), Ph. mascomai (13), Ph. sinxayarami (nine), Ph. stantoni (12), Se. anodontis (one), Se. hivernus (seven), Se. dvoraki n. sp. (10), Se. khawi (19), Se. phasukae (six), Se. rudnicki (11), Se. siamensis (11), Se. sylvatica (one), and Gr. indica (four). We did not sequence Cyt-b of other rare species, such as Ph. barguesae, Se. brevicaulis-like, Se. perturbans, and Se. marolii n. sp., because the whole bodies of all these samples were mounted (in toto). Together with Cyt-b sequences available from Thailand, a total of 170 sequences were analyzed (Additional file 2: Table S2). A phylogenetic ML tree constructed using the Tamura–Nei model, which was the best-fit nucleotide substitution model for this study, showed that all species used in these analyses formed monophyletic clades, in particular for the Chinius, Grassomyia, Idiophlebotomus, and Phlebotomus genera. Interestingly, for the genus Sergentomyia, all the Se. siamensis from this study formed a subclade with two other subclades of the Se. barraudi group from Thailand, labeled Se. barraudi group 1 (MG770912, MK442497, MK442494, MK442496, and MK442500) and group 2 (MG770903) (Fig. 5). The mean pairwise distance between Se. siamensis and Se. barraudi group 1 and group 2 was 0.08 and 0.07, respectively, and between Se. rudnicki and Se. siamensis it was 0.10. Genetic analysis supported our morphological observation on the most closely related species. The mean pairwise distance between Se. dvoraki n. sp. and Se. khawi was 0.12, and between Se. dvoraki n. sp. and Se. hivernus it was 0.13. The mean pairwise distances within groups ranged from 0 to 0.018, and between group species it ranged from 0.07 to 0.41. (Further details on the pairwise mean distances between and within species are also provided in Additional file 3: Table S3.)

Maximum likelihood tree constructed from cytochrome b gene using the Tamura–Nei model
Sand fly diversity and species assemblages across sampling sites and cave locations
Among the sand flies identified, 3294 (50.18%) were collected from Feung (72 trap-nights, density (D) = 45.75 [Min–Max: 2–202]), 1494 (22.76%) from Hinheup (40 trap-nights, D = 37.35 [Min–Max: 2–115]), and 1776 (27.06%) from Vangvieng (90 trap-nights, D = 19.73 [Min—Max: 1–102]) (Table 1). For the diversity analysis, species accumulation curves were plotted by site, season, and position of trapping to estimate whether they could be representative of total sand fly identification. Species accumulation reached a plateau in this study, especially during the rainy season, with a high overall Shannon diversity index H in the three districts (Additional file 4: Fig. S1). In Feung district, the species richness (R) = 19, Shannon index (H) = 2.46, and the most abundant species were Ch. eunicegalatiae (20.31%), Id. longiforceps (8.80%), Ph. stantoni (12.60%), and Se. anodontis (8.08%); in Hinheup district, R = 17 species, H = 2.24, and the most abundant species were Ch. eunicegalatiae (31.86%), Ph. stantoni (8.03%), Se. khawi (9.17%), and Se. rudnicki (7.3%); and in Vangvieng district, R = 17 species, H = 2.26, and the most abundant species were Ch. eunicegalatiae (23.09%), Se. hivernus (19.43%), and Se. siamensis (9.23%) (Table 1). Regarding the species assemblages between districts, the Bray‒Curtis index values for all the trap-nights differed significantly across the three sampling districts, especially between Vangvieng and Hinheup and between Vangvieng and Feung, but there was no significant difference between Hinheup and Feung (Additional file 5: Fig. S2).
At the cave location level, we observed that sand fly density was high in all cave areas (D > 30 sand flies/trap-night), except in NOK cave in the Vangvieng district, where only eight sand flies per trap-night were found. The species richness ranged from 14 to 18, of which the lowest was in the NOK cave in Vangvieng district (Additional file 6: Table S4). Species assemblages differed greatly among our seven sampling locations (Additional file 5: Fig. S2).
Different density and H index values were also observed among the trapping positions. For sand flies outside caves, R = 20, D = 34.29, and H = 2.50; and for cave entrances, R = 18, D = 40.28, and H = 2.41. These values were significantly greater than those for inside caves: R = 16, D = 22.57, and H = 2.13. Among all the sand fly species, Ch. eunicegalatiae and Ph. stantoni were the most abundant species inside, at the entrance, and outside caves (Additional file 7: Table S5).
From two collections, one in the dry season and another in the rainy season, the sand fly density was high in both the rainy and dry seasons in Fueng and Hinheup. However, it was low in the dry season in Vangvieng (D = 6.29) (Additional file 8: Table S6).
Descriptions of new species
Family Psychodidae Newman, 1834
Genus Sergentomyia França and & Parrot, 1920
Subgenus ungrouped awaiting a Sergentomyia taxonomic revision.
Sergentomyia dvoraki n. sp. Randrianambinintsoa, Vongphayloth and Depaquit, 2024 (Fig. 6 and Table 2).

Sergentomyia dvoraki n. sp. female. A Head (paratype LAOS#251-2); B flagellomeres 1, 2, and 3 (= AIII, AIV, and AV) (paratype LAOS#251-2); C pharynx and cibarium (holotype LAOS#251-3); D palp (paratype LAOS#251-2); E third segment of the palp (P3) (paratype LAOS#251-2); F mouth parts (labrum, hypopharynx, mandible, maxilla [paratype LAOS#251-2], and labial furca [paratype LAOS#251-2] from left to right); G wing (holotype LAOS#251-3); H furca and spermathecae (holotype LAOS#251-3)
Type locality Pha Nok Kok cave (18°30′N, 101°59′E), Feung district, Vientiane Province, Laos.
Type specimens The female holotype (voucher LAOS-251-3) of Se. dvoraki n. sp. and two female paratypes (voucher LAOS-251-1, LAOS-251-2) have been deposited at the terrestrial arthropod collection of the Muséum national d’Histoire naturelle (MNHN, Paris) under inventory numbers MNHN-ED-115949–MNHN-ED-11951.
ZooBank registration To comply with the regulations set out in Article 8.5 of the amended 2012 version of the International Code of Zoological Nomenclature (ICZN) [35], details of the new species have been submitted to ZooBank. The Life Science Identifier (LSID) of the article is urn:lsid:zoobank.org:pub:AB876763-B1CC-4414-A45E-24247A3F33CD. The LSID for the new species Se. dvoraki is urn:lsid:zoobank.org:act:566AB258-2548-41BF-ADE1-1878E0730B85.
Etymology The species Se. dvoraki n. sp. is dedicated to our colleague Vit Dvořák for his important contribution to research in the field of phlebotomine sand flies and leishmaniasis.
Description
Measurements and counts indicated are those of the holotype (voucher LAOS-251).
Female: Head: occiput with two narrow lines of well-individualized setae. Clypeus 115 µm long and 86 µm wide with 42 setae randomly distributed. Eyes 177 µm long, 97 µm wide, with about 90 facets. Interocular sutures incomplete. Interantennal sutures do not reach the interocular sutures. Flagellomere 1 longer than f2 + f3. Presence of two ascoids from f1 to f13. Absence on f14. Ascoidal formula: 2/f3–f13 with long ascoids, not reaching the next article. One papilla on f1 and f2. Absence of papilla from f3 to f9, two papillae on f10 and f11 (one basal and one apical), four on f12 and 13 (two basal and two apical), and four basal papillae on f14. No simple setae on f1 to f4, one on f5, f6, f8, and f9; two on f7; 20–25 on f14. Palpal formula: 1, 2, 3, 4, 5. Presence of a group of about 10 club-like Newstead’s sensillae implanted proximally on the third palpal segment. Presence of one distal simple seta on p3, eight on p4, and 16 on p5 on the paratype LAOS#251-2 (the p5 of the holotype is broken). Labrum 164 µm long. About 20 distal teeth on each side. Hypopharynx with around 15 distal undulations on each side of the salivary canal. Mandibles with a little more than 30 small lateral teeth. Maxillary lacinia exhibits eight external and about 25 internal teeth. Labial furca incomplete. Cibarium armed with 18 posterior teeth, and two central rows of anterior teeth: six bigger and six smaller. Sclerotized area rounded, not reaching the lateral side of the cibarium. Sclerotized arc not observed. Thorax: dark brown sclerites. Mesonotum: absence of post-alar seta. Absence of proepimeral; absence of upper and lower anapisternal seta; absence of anepimeral seta; absence of metaepisternal seta; absence of metaepimeral seta; presence of some fine setae on the anterior region of the katepisternum. Metafurca with long and separated vertical and horizontal arms. Two cervical sensillae on each side. One ventro-cervical sensilla. Wings: length = 1536 μm; width = 401 μm. r5 = 1049 μm, α (r2) = 224 μm, β (r2 + 3) = 324 μm, δ (r2 + 3-r1) = 80 μm, γ (r2 + 3 + 4) = 250 μm, ε (r3) = 343 μm, θ (r4) = 782 μm, π = 157 μm. Width/γ = 1.60, α (r2)/β (r2 + 3) = 0.69. Legs: anterior leg: coxa = 254 µm; femur = 510 µm; tibia = 492 µm; tarsomere i = 274 µm; sum of tii, tiii, tiv, tv = 455 µm.
Median leg: coxa = 226 µm; femur = 555 µm; tibia = 597 µm; tarsomere i = 322 µm; sum of tii, tiii, tiv, tv = not observed. Posterior leg: coxa = 270 µm; femur = 648 µm; tibia = 748 µm; tarsomere i = 387 µm; sum of tii, tiii, tiv, tv = 564 µm. Abdomen: setae randomly distributed on the first tergite. Genitalia: narrow wrinkled spermathecae, with thin walls. Terminal knob deeply embedded in the capsule. Short smooth common duct. Long smooth individual ducts. Genital furca with two thin and well-developed lateral process.
Male: Unknown.
Family Psychodidae Newman, 1834
Genus Sergentomyia França and & Parrot, 1920
Subgenus ungrouped awaiting a Sergentomyia taxonomic revision.
Sergentomyia marolii n. sp. Vongphayloth, Randrianambinintsoa and Depaquit, 2024 (Fig. 7A–H and Table 3).

Sergentomyia marolii n. sp. female. A Head (paratype S-1610-2); B pharynx and cibarium (holotype S-2553-4); C mouth parts (labrum, hypopharynx, mandible, maxilla, and labial furca from left to right) (paratype S-1610-2); D flagellomeres 1, 2, and 3 (= AIII, AIV, and AV) (holotype S-2553-4); E third segment of the palp (P3) (holotype S-2553-4); F palp (paratype S-1610-2); G wing (holotype S-2553-4); and H furca and spermathecae (paratype)
Type locality Pha Nok Kok cave (18°30′N, 101°59′E), Feung district, Vientiane Province, Laos.
Type specimens The holotype female (voucher S-2553-4) and one female paratype (voucher S-1610-2) of Se. marolii n. sp. have been deposited at the terrestrial arthropod collection of the Muséum national d’Histoire naturelle (MNHN, Paris) under inventory numbers MNHN-ED-115952 and MNHN-ED-115953.
ZooBank registration To comply with the regulations set out in Article 8.5 of the amended 2012 version of the International Code of Zoological Nomenclature (ICZN) [35], details of the new species have been submitted to ZooBank. The Life Science Identifier (LSID) of the article is urn:lsid:zoobank.org:pub:AB876763-B1CC-4414-A45E-24247A3F33CD. The LSID for the new species Se. marolii is urn:lsid:zoobank.org:act:FB152938-A7FD-4B14-90F7-73485F508B94.
Etymology The species Se. marolii n. sp. is dedicated to our colleague Michele Maroli for his important contribution to research in the field of phlebotomine sand flies and leishmaniasis, and in hosting the first International Symposium on Phlebotomine Sandflies (ISOPS).
Description
Measurements indicated are those of the holotype (voucher S-2553-4.)
Female: Head: occiput with two narrow lines of well-individualized setae. Clypeus 112 µm long and 77 µm wide with 34 setae randomly distributed. Eyes 167 µm long, 93 µm wide, with about 80 facets. Interocular sutures incomplete. Interantennal sutures do not reach the interocular sutures. Flagellomere 1 shorter than f2 + f3. Presence of two ascoids from f1 to f13 and absence on f14. Ascoidal formula: 2/f3-f13 with ascoids not reaching the next article. One papilla on f1 and f2. Absence of papilla from f3 to f9. One on f10 and f11, three from f12–f14. No simple setae from f1 to f11, one on f12, two on f13 and less than 25 on f14. Palpal formula: 1, 2, 3, 4, 5. Presence of a group of about 30 club-like Newstead’s sensillae implanted on the proximal half on the third palpal segment. Presence of one distal simple seta on p3, 17 on p4, and about 30 on p5. Labrum 228 µm long with about 15 lateral teeth. Smooth lateral sides of the hypopharynx. Thin maxillary lacinia with about 90 lateral little teeth. Labial furca complete. Cibarium armed with 11 big teeth organized along a very slightly curved line. No denticles (anterior teeth). Sclerotized area strongly pigmented, thick and narrow (not reaching the lateral walls of the cibarium). Pharynx strongly armed with pointed translucent teeth oriented backwards, on its posterior quarter. Thorax: light brown sclerites. Mesonotum: absence of post-alar seta. Proepimeral setae non-observed; absence of upper anepisiternal seta; absence of lower anepisiternal setae; absence of anepimeral seta; absence of metaepisternal seta; absence of metaepimeral seta; presence of a tuft of long and fine setae on the anterior region of the katepisternum. Metafurca with separated long vertical and short horizontal arms. Mesofurca dark brown.
Cervix with two cervical sensillae on each side. Two ventro-cervical sensillae. Wings: length = 1582 μm; width = 435 μm. r5 = 1151 μm, α (r2) = 203 μm, β (r2 + 3) = 355 μm, δ (r2 + 3-r1) = 76 μm, γ (r2 + 3 + 4) = 289 μm, ε (r3) = 362 μm, θ (r4) = 815 μm, π = 80 μm. Width/γ = 1.5, α (r2)/β (r2 + 3) = 0.57. Legs (on S-2553-4): anterior leg: coxa = 247 µm; femur = 566 µm; tibia = 604 µm; tarsomere i = 339 µm; sum of tii, tiii, tiv, tv = 516 µm. Median leg: coxa = 263 µm; femur = 617 µm; tibia = 763 µm; tarsomere i = 427 µm; sum of tii, tiii, tiv, tv = 563 µm. Posterior leg: coxa = 273 µm; femur = 627 µm; tibia = 967 µm; tarsomere i = 485 µm; sum of tii, tiii, tiv, tv = 617 µm. Abdomen: setae randomly distributed on the first tergite. Genitalia: short, wrinkled oblong spermathecae, with thin walls. Terminal knob not embedded in the capsule, with setae around the knob and lateral structures surrounding it. On paratype S-1610-2: smooth individual ducts about 160 µm, common duct was not observed. Genital furca with a wide handle.
Male: Unknown.
Discussion
This is the first inventory of the sand fly fauna in karstic limestone areas known in Laos. Before these data, a total of 15 species were reported from Laos, including (1) Ch. eunicegalatiae, (2) Gr. indica [as Phlebotomus (Se.) squamipleuris], (3), Ph. argentipes, (4) Ph. breyi, (5) Ph. sinxayarami, (6) Ph. shadenae, (7) Ph. stantoni, (8) Se. bailyi, (9) Se. barraudi, (10) Se. gemmea, (11) Se. hivernus, (12) Se. iyengari (may correspond to Se. dvoraki n. sp.), (13) Se. khawi, (14) Se. perturbans, and (15) Se. sylvatica [18, 22,23,24,25]. Here, we added 10 more species found in Laos, including (1) Id. longiforceps, (2) Ph. barguesae, (3) Ph. kiangsuensis, (4) Ph. mascomai, (5) Se. anodontis, (6) Se. brevicaulis-like, (7) Se. dvoraki n. sp., (8) Se. marolii n. sp., (9) Se. phasukae, and (10) Se. rudnicki. Dichotomous keys for females and males found in this study are provided in Tables 4 and 5. The updated list of the sand fly species of Laos includes now 25 species.
The present study also highlights the high abundance and diversity of sand flies in all cave locations and districts in karstic areas of Vientiane Province. Heterogeneous species composition according to caves, trapping positions, and seasons was also observed. Similar results of high density and diversity, especially in karstic cave areas, and seasonal variety were also observed in Thailand, where more than 27 species of sand flies have been identified [36,37,38,39]. Differences in sand fly diversity and species composition between different specific environments suitable for sand flies were also observed elsewhere in SE Asia, Africa, and South America [40,41,42]. The lowest sand fly density found in this study was at NOK cave in Vangvieng district, which could be explained by the fact that this large cave is located higher from the ground than other caves are, and the cave floor might not be suitable for the development of sand fly larvae due to the substrate with low levels of organic materials. Although many sand fly species are widely distributed among SE Asian countries, many of them might be restricted to a specific habitat within an area. However, our study has limitations. The sampling method used may not have been optimal. The traps and their placement might not have captured all species, possibly underestimating the true diversity and richness. Differences in trap effectiveness, weather conditions, and timing during sampling can also cause biases in the data. To address these limitations, future studies should use more standardized and extensive sampling methods, including multiple techniques and repeated sampling over time. A more detailed analysis of beta diversity could also offer better insights into the ecological relationships and processes affecting sand fly populations in different districts. More intensive studies of sand flies in Laos, as well as other countries in the region, will provide a clearer picture of sand fly diversity and ecology.
Taxonomic and biological comments on the species recorded in Laos
Genus Chinius
Chinius eunicegalatiae was originally described from a cave in Vangvieng district [23], where this area was also selected for collection in this study. This species, as well as the genus Chinius, was considered exclusively cave-dwelling [23]. Interestingly, we observed in the present study that Ch. eunicegalatiae was not restricted only to caves, taking into consideration we also caught it inside caves or at the entrance of the caves, as well as outside caves. In the future, new studies placing traps at successive distances (at least 5 m) from the cave entrance would make it possible to learn more about the habitat of this species. It seems that this species is distributed only in the limestone areas of Laos. Until now, it has not been reported in other countries in this region. Chinius eunicegalatiae shares with Ch. barbazani the lack of r2 veins on the wing in both genders. They differ mostly in males by the position of the most proximal spine of the gonostyle, by the beveled or truncated end of the aedeagal ducts, and by the sperm pump/aedeagal ducts ratio. The females differ by the diameter of the spermathecae and by the lengths of (spermathecae + individual duct)/length of common duct ratio.
Genus Idiophlebotomus
Idiophlebotomus longiforceps had never been recorded in Laos before this study. It was found inside caves or at the entrance of the caves, as well as outside caves. Our morphological characteristics and genetic analysis revealed that the Laotian specimens were similar to those recorded in Thailand and identified as Id. longiforceps [43]. However, this species was described from China several decades ago, in the genus Sergentomyia [44], and the differential characters within the genus Idiophlebotomus are not precisely detailed. Lewis, in his review of sand flies of the Eastern Region [18], did not report this species from SE Asia, but it has recently been reported from Vietnam [42] and Thailand [27, 45]. A revision of this genus with more specimens and with molecular studies together with comparisons to type specimens is still warranted. But here, we identified this species based on morphological characteristics similar to those of male genitalia according to the original description and identification key provided by Loyer et al. [43]. The male has a slender and rounded aedeagus, with three spines on the style: one apical, one proximal that is inserted at 2/3 of the style, and the last one is inserted near the proximal one. The coxite from the original description of this species was broader than that of other known Idiophlebotomus species in the revision of Lewis [18].
Genus Phlebotomus
Before the studies carried out by our laboratories, only two species were reported from Laos by Quate [24]: Ph. argentipes from Luangphabang and Ph. stantoni from Vientiane. We recently recorded and described three new species in Laos, including Ph. breyi, Ph. shadenae, and Ph. sinxayarami [25, 27]. In the present study, we also report Ph. barguesae, Ph. kiangsuensis, and Ph. mascomai in Laos. Phlebotomus mascomai, Ph. sinxayarami, and Ph. stantoni were the most abundant among this genus and were found inside, at the entrance, and outside caves. It is likely that these species could seek a host from both inside and outside caves. It was already observed that Ph. kiangsuensis can bite humans and attack bats [18]. For an accurate identification of this species, a comparison with Chinese topotypes is necessary to obtain both molecular and morphological characters. Phlebotomus stantoni is a widespread species in SE Asia, which seems not restricted to the karstic habitat, as it was recorded from different environments in neighboring countries of Laos [38, 42].
Phlebotomus argentipes was first described in India, and its geographical variety in terms of morphology and biology has been discussed [18, 46]. The recorded Ph. argentipes in Laos as well as in other SE Asian countries may be confused with closely related species, i.e., Ph. mascomai. A wide-scale study including many specimens studied by both morphology and genetics should be welcome. Phlebotomus argentipes was not found in the present study.
Genus Sergentomyia
The genus Sergentomyia is considered a catch-all group. As its role in the transmission of pathogens is quite limited, it has been less studied than the genus Phlebotomus. Some subgenera seem to be monophyletic, i.e., Sergentomyia sensu stricto, Sintonius, Capensomyia, Vattieromyia, Trouilletomyia, or Ranavalonomyia. The taxonomy of the genus Sergentomyia remains quite confusing, and the validity of some subgenera seems questionable to us, i.e., Neophlebotomus or Parrotomyia. Moreover, some species remain dubious, and the boundaries of other species are quite vague.
We report Se. anodontis sensu lato for the first time in Laos. Sergentomyia anodontis s.s. was described by Quate and Fairchild from Batu Cave in peninsular Malaysia. The main characteristics of this species were described as follows: “an unarmed cibarium, bur with spine-like projections from fold in membrane above sclerotized part and with median projection over which is inverted V-shaped bar, and spermatheca long and tubular, no differentiation between spermatheca and duct until point of junction of individual duct, apex with thick, hairy, sunken knob, several annulations below knob” [47]. The originality of its cibarium makes it very easily identifiable. For this reason, this species is not observed in detail in routine, and it seems to us that Se. anodontis s.l. could actually constitute a species complex. The specimens found in this study agree with this description. However, among the samples in this study, a pair of ascoids was found on flagellomere 1 (f1) to f13, which is different from the original description where a pair of ascoids found on f1 to f11 and a single ascoid on f12 to f14 have been observed. Different cibarial teeth were also observed in specimens from Vietnam [42]. Moreover, a recent genetic study of this species using cytochrome c oxidase I (COI) in Thailand showed that this species was divided into two clusters [48]. The genetic analysis using Cyt-b in this study showed that our samples were identical to some specimens from Thailand [27]. However, the number of sequence samples used for our analysis is small, and further larger-scale studies are needed to clarify the taxonomy status of this probable complex of species. For this study, we considered this species as Se. anodontis s.l.
Sergentomyia bailyi s.l. was recorded in Laos by Quate from burned tree holes [24] and was not found in the present study. As already discussed by Vu et al. [42] on sand flies from Vietnam, the reports of this species in Laos as well as other SE Asian countries still need to be revised by studying more samples and genetic studies.
The Se. barraudi group was also recorded in Laos by Quate [24]. The Se. barraudi group should be further studied because of the existence of considerable morphological and molecular heterogeneity in the morphological characters, such as the number and distribution of teeth on the cibarium. The cibarium in the original description made [49] from Indian specimens included 40 teeth and a forked anterior part of the sclerotized area. The specimens caught in Laos exhibited a similar forked sclerotized area and 50 to 54 cibarial teeth. Consequently, we identified the specimens of the present study as Se. siamensis considering that both morphological characters (Fig. 2A, D, G, J) and molecular markers were similar to those reported and already discussed in Thailand [27]. The results of the Cyt-b analysis in this study, together with the results of the COI analysis of the Se. barraudi group in Thailand [48], highlights the need for further taxonomic investigation in the SE Asian region. A complete review of the barraudi group is highly desirable. It will have to include numerous populations from various countries and combine morphological and molecular approaches.
Sergentomyia rudnicki is closely related to Se. brevicaulis and Se. barraudi group in presenting of comb-like teeth on cibarium. The Se. barraudi group could be separated from Se. rudnicki and Se. brevicaulis by their shorter antennae flagellomere 1, sclerotized area of the cibarium, and ovoid and smooth spermathecae (Fig. 2A and J). The specimens classified as Se. rudnicki from the present study agree with the original description of Se. rudnicki by female cibarium presenting about 90 comb-like teeth, two rows of about 20 vertical teeth, long pharyngeal teeth oriented backwards, and oblong and annulate spermatheca [18]. However, among the Laotian specimens, about 80 comb-like teeth on the cibarium with two rows of vertical teeth of more than 30 on each row were observed (Fig. 2F). Regarding Se. brevicaulis-like found in this study, cibarium has about 50 comb-like teeth (Fig. 2E), which is similar to the original description of Se. brevicaulis [24], but pharyngeal teeth long (Fig. 2H) like those in Se. rudnicki (Fig. 2I). Further studies are needed to clarify the taxonomic status of Se. rudnicki and the Se. brevicaulis group.
Sergentomyia dvoraki n. sp. is a species belonging to the iyengari group of Sergentomyia. Historically, Se. iyengari Sinton was described in 1933 from specimens caught in Southwest India without anterior teeth (formerly fore-teeth) and without any description of spermatheca [50]. Later, in 1935–1936, Raynal redescribed both the female and male of Se. iyengari Sinton using specimens from Vietnam. Raynal also illustrated both anterior teeth and wrinkled spermathecae [51] without suspecting that the samples from Vietnam could be different from those from Southwest India, making the situation confusing. For us, the description by Raynal of Se. iyengari from Vietnam may probably be the species that we described here. Sergentomyia dvoraki n. sp. could easily be separated from Se. iyengari because the latter species does not exhibit anterior teeth on the cibarium. The recently described Se. ashwanii [52] from India has a spermatheca very different from that of Se. dvoraki n. sp., without the terminal knob embedded in a deep notch. Sergentomyia dvoraki n. sp. has cibarial teeth similar to those in Se. hivernus by presenting 2–12 anterior teeth (Fig. 3B and C) but can be distinguished from each other by spermathecae that are tubular without limits between the body and duct in Se. hivernus [53], as shown in Fig. 3E and F. Sergentomyia hivernus was also historically wrongly considered a junior synonym of Se. iyengari Sinton in SE Asia. After more samples became available and additional genetics were analyzed recently, the previous records of Se. iyengari in Laos may refer to Se. hivernus. The spermathecae of Se. dvoraki n. sp. are similar to those of Se. khawi but with wrinkled bodies, and longer and narrower individual ducts than those of Se. khawi (Fig. 3D and F). In Se. khawi, the cibarium has many anterior (vertical) teeth arranged along 2–3 rows [54] (Fig. 3A). Sergentomyia gemmea can be easily separated from other species by the ascoids on antennae with spurs (Fig. 3I) but absent in Se. khawi. The records of Se. gemmea [18] in Laos are pending revision, as the cibarium appears to be different, and the pairwise distance of the Cyt-b gene between specimens from Laos and Thailand specimens was 0.06 [22]. A complete review of the iyengari group is highly desirable. It will have to include numerous populations from various countries and combine morphological and molecular approaches.
Sergentomyia phasukae recorded in Laos in this study has the same characteristics as those described by Curler [55]. This species is closely related to Se. quatei. Our specimens had no pigment patch on the cibarium (Fig. 3G). The arrangement of cibarial teeth looks somewhat different from the original description, but we think this is because of the mounting. The common spermathecal duct is as long (Fig. 3J) as in the original description of Se. phasukae.
Sergentomyia sylvatica was recorded in Laos by Quate in Vientiane collected from tree holes [24]. In this study, Se. sylvatica was found in all cave locations in karstic limestone in Vientiane Province. The spermathecae and cibarial (Fig. 3H and K) agree with the original description of this species in Vietnam by Raynal [56].
Sergentomyia perturbans was recorded in Laos by Quate as Phlebotomus (Se.) sylvestris collected from tree holes [24]. Later, this species was synonymized with Se. perturbans. As already discussed by Vu et al. [42] on sand flies from Vietnam, the taxonomic status of Se. perturbans remains doubtful. The record of this species in Laos should be considered as uncertain. We followed the morphological characteristics described by Lewis [18]: “cibarium with eight or nine distinct pointed teeth merging into about 10 spicules on each side, arising from thick refractive band, fore-teeth absent, pigment patch dark reddish brown anteriorly and grey posteriorly, with transverse and oblique line, bearing anteriorly about eight longitudinal lines; distinct cibarial bulge present. Pharynx less than twice as wide posteriorly as anteriorly, with faint ridges bearing minute spicules.” Images of the cibarium and pharynx of the Laotian samples are shown in Fig. 4A, D, and F.
Sergentomyia marolii n. sp. is morphologically closely related to Se. bigtii [57] from the Philippines, as indicated by the original description of the arrangement of 10 to 12 teeth on the cibarium and long teeth on the pharynx (Fig. 4B and E). However, this species can be easily distinguished from Se. bigtii by its shorter antennae f1. The metafurca of this species is strongly pigmented, similar to that of the specimens of Se. perturbans included in this study (Fig. 4C and F). It is interesting to further investigate whether this character could be used at subgenus level. Only two specimens were found during this study in Vientiane Province. Both were mounted in toto, and no DNA sequence is available for this new species.
Genus Grassomyia
Grassomyia indica s.l. was first reported in Laos from Vientiane and Luangphabang provinces as Phlebotomus (Se.) squamipleuris by Quate [24]. The genus Grassomyia, also considered as a subgenus of Sergentomyia by several authors without consensus, is not implicated in the transmission of pathogens, and consequently, its taxonomy remains poorly documented. In this study, we consider our specimens as Gr. indica s.l. because we assume that our specimens from Laos, and more broadly from SE Asia, probably belong to a species other than that described from India, but this hypothesis must be justified by a global morphological and molecular study including all Grassomyia species, as discussed previously [42]. In the present study, Grassomyia specimens were found mainly from cave entrances in Feung district. Their morphology and genetics are similar to those reported from Thailand [58] and may be as those that found in Vietnam, as the number of cibarial teeth ranged from 25 to 33.
Conclusions
This study highlighted the high diversity of phlebotomine sand flies fauna in Laos, which was previously underestimated in karstic limestone areas. However, the taxonomic status of many species in Laos, as well as in other countries of Southeast Asia, still needs more in-depth study using both morphological characters and molecular methods. Many species could be found from inside, at the entrance, and outside of caves, indicating a wide range of host-seeking behavior in the karstic cave areas.
Availability of data and materials
All specimens collected during this study were examined and deposited at the Institut Pasteur du Laos Collection Room. Sequences of Cyt-b obtained from this study were deposited in the GenBank database (PQ151806–PQ151934). No datasets were generated or analyzed during the current study.
Abbreviations
- AL:
-
Angluang karstic areas
- COI:
-
Cytochrome c oxidase I
- Cyt-b :
-
Cytochrome b
- f1–f14:
-
Flagellomere 1–14
- NOK:
-
Tham Nang Oau Khiem cave
- PLS:
-
Tham Phaluesy cave
- PNK:
-
Tham Phanokkok cave
- r1–r5:
-
Radial wing vein branches 1–5
- ti–v:
-
Tarsomere i–v
- TN:
-
Tham Nam cave
- TP:
-
Tham Pha cave
- TY:
-
Tham Yao cave
References
Jones KE, Patel NG, Levy MA, Storeygard A, Balk D, Gittleman JL, et al. Global trends in emerging infectious diseases. Nature. 2008;451:990–3.
Chan JF, Choi GK, Yip CC, Cheng VC, Yuen KY. Zika fever and congenital Zika syndrome: an unexpected emerging arboviral disease. J Infect. 2016;72:507–24.
Gratz NG. Emerging and resurging vector-borne diseases. Annu Rev Entomol. 1999;44:51–75.
Vasconcelos PF, Calisher CH. Emergence of human arboviral diseases in the Americas, 2000–2016. Vector Borne Zoonotic Dis. 2016;16:295–301.
Harrus S, Baneth G. Drivers for the emergence and re-emergence of vector-borne protozoal and bacterial diseases. Int J Parasitol. 2005;35:1309–18.
Dash AP, Bhatia R, Sunyoto T, Mourya DT. Emerging and re-emerging arboviral diseases in Southeast Asia. J Vector Borne Dis. 2013;50:77–84.
Obame-Nkoghe J, Leroy EM, Paupy C. Diversity and role of cave-dwelling hematophagous insects in pathogen transmission in the Afrotropical region. Emerg Microbes Infect. 2017;6:e20.
Jurado V, Laiz L, Rodriguez NV, Boiron P, Hermosin B, Sanchez MS, et al. Pathogenic and opportunistic microorganisms in caves. Int J Speleol. 2010;39:15–24.
Konstantinov OK, Diallo SM, Inapogi AP, Ba A, Kamara SK. The mammals of Guinea as reservoirs and carriers of arboviruses. Med Parazitol. 2006;1:34–9.
Gomez-Hernandez C, Bento EC, Rezende-Oliveira K, Nascentes GAN, Barbosa CG, Batista LR, et al. Leishmania infection in bats from a non-endemic region of Leishmaniasis in Brazil. Parasitol. 2017;144:1980–6.
Maganga GD, Bourgarel M, Vallo P, Dallo TD, Ngoagouni C, Drexler JF, et al. Bat distribution size or shape as determinant of viral richness in african bats. PLoS ONE. 2014;9:e100172.
McKee CD, Kosoy MY, Bai Y, Osikowicz LM, Franka R, Gilbert AT, et al. Diversity and phylogenetic relationships among Bartonella strains from Thai bats. PLoS ONE. 2017;12:e0181696.
Anh PH, Van Cuong N, Son NT, Tue NT, Kosoy M, Woolhouse ME, et al. Diversity of Bartonella spp. in Bats Southern Vietnam. Emerg Infect Dis. 2015;21:1266–7.
Kamani J, Baneth G, Mitchell M, Mumcuoglu KY, Gutierrez R, Harrus S. Bartonella species in bats (Chiroptera) and bat flies (Nycteribiidae) from Nigeria West Africa. Vector Borne Zoonotic Dis. 2014;14:625–32.
Adam JP. Transmission of Hemosporidia by Anopheles Cavernicolus in the Caves of the Congo (Brazzaville). Bull World Health Organ. 1965;32:598–602.
Grard G, Fair JN, Lee D, Slikas E, Steffen I, Muyembe JJ, et al. A novel rhabdovirus associated with acute hemorrhagic fever in central Africa. PLoS Pathog. 2012;8:e1002924.
Mittermeier RA, Turner WR, Larsen FW, Brooks TM, Gascon C. Global biodiversity conservation: the critical role of hotspots. In: Zachos FE, Habel JC, editors. Biodiversity hotspots. Berlin Heidelberg: Springer; 2011. p. 3–22.
Lewis DJ. The phlebotomine sandflies (Diptera: Psychodidae) of the oriental region. Bull br Mus nat Hist Entomol. 1978;37:217–343.
Thisyakorn U, Jongwutiwes S, Vanichsetakul P, Lertsapcharoen P. Visceral leishmaniasis: the first indigenous case report in Thailand. Trans R Soc Trop Med Hyg. 1999;93:23–4.
Pothirat T, Tantiworawit A, Chaiwarith R, Jariyapan N, Wannasan A, Siriyasatien P, et al. First isolation of Leishmania from Northern Thailand: case report, identification as Leishmania martiniquensis and phylogenetic position within the Leishmania enriettii complex. PLoS Negl Trop Dis. 2014;8:e3339.
Leelayoova S, Siripattanapipong S, Manomat J, Piyaraj P, Tan-Ariya P, Bualert L, et al. Leishmaniasis in Thailand: a review of causative agents and situations. Am J Trop Med Hyg. 2017;96:534–42.
Depaquit J, Vongphayloth K, Siriyasatien P, Polseela R, Phumee A, Loyer M, et al. On the true identity of Sergentomyia gemmea and description of a closely related species: Se. raynali. n. sp. Med Vet Entomol. 2019;33:521–9.
Léger N, Depaquit J, Gay F. Chinius eunicegalatiae n. sp. (Diptera; Psychodidae), a cavernicolous sandfly from Laos. Ann Trop Med Parasitol. 2010;104:595–600.
Quate LW. A review of the Indo-Chinese Phlebotominae. Pac Insects. 1962;4:251–67.
Vongphayloth K, Randrianambinintsoa FJ, Lakeomany K, Phommavanh N, Mekarnia N, Khadri MS, et al. On the systematics of Phlebotomus betisi and two new related species from Laos with proposal of the new subgenus Lewisius. Parasite. 2023;30:21.
Lewis DJ. A Taxonomic review of the genus Phlebotomus (Diptera, Psychodidae). Bull br Mus nat Hist Entomol 1982.
Renaux Torres MC, Pellot C, Somwang P, Khositharattanakool P, Vongphayloth K, Randrianambinintsoa FJ, et al. Phlebotomine sand flies (Diptera, Psychodidae) from Pha Tong cave, Northern Thailand with a description of two new species and taxonomical thoughts about Phlebotomus stantoni. PLoS Negl Trop Dis. 2023;17:e0011565.
Depaquit J, Muller F, Léger N. Phlebotomus (Euphlebotomus) barguesae n. sp. from Thailand (Diptera-Psychodidae). Parasit Vectors. 2009;2:5.
Muller F, Depaquit J, Léger N. Phlebotomus (Euphlebotomus) mascomai n. sp. (Diptera-Psychodidae). Parasitol Res. 2007;101:1597–602.
Galati EAB, Galvis-Ovallos F, Lawyer P, Léger N, Depaquit J. An illustrated guide for characters and terminology used in descriptions of Phlebotominae (Diptera, Psychodidae). Parasite. 2017;24:26.
Esseghir S, Ready PD, Killick-Kendrick R, Ben-Ismail R. Mitochondrial haplotypes and geographical vicariance of Phlebotomus vectors of Leishmania major. Insect Mol Biol. 1997;6:211–25.
Shannon CE. The mathematical theory of communication 1963. MD Comput. 1997;14:306–17.
Tamura K, Stecher G, Kumar S. MEGA11: molecular evolutionary genetics analysis version 11. Mol Biol Evol. 2021;38:3022–7.
Posada D, Crandall K. MODELTEST: testing the model of DNA substitution. Bioinform. 1998;14:817–8.
International Commission On Zoological Nomenclature. Amendment of articles 8, 9, 10, 21 and 78 of the international code of zoological nomenclature to expand and refne methods of publication. Zookeys. 2012;219:1–10.
Apiwathnasorn C, Samung Y, Prummongkol S, Phayakaphon A, Panasopolkul C. Cavernicolous species of phlebotomine sand flies from Kanchanaburi Province, with an updated species list for Thailand Southeast Asian. J Trop Med Public Health. 2011;42:1405–9.
Apiwathnasorn C, Sucharit S, Rongsriyam Y, Leemingsawat S, Kerdpibule V, Deesin T, et al. A brief survey of phlebotomine sandflies in Thailand Southeast Asian. J Trop Med Public Health. 1989;20:429–32.
Polseela R, Apiwathnasorn C, Samung Y. Seasonal variation of cave-dwelling phlebotomine sandflies (Diptera:Psychodidae) in Phra Phothisat Cave, Saraburi Province, Thailand Southeast Asian. J Trop Med Public Health. 2007;38:1011–5.
Polseela R, Depaquit J, Apiwathnasorn C. Description of Sergentomyia phadangensis n. sp. (Diptera, Psychodidae) of Thailand. Parasit Vectors. 2016;9:21.
Obame-Nkoghe J, Rahola N, Ayala D, Yangari P, Jiolle D, Allene X, et al. Exploring the diversity of blood-sucking Diptera in caves of Central Africa. Sci Rep. 2017;7:250.
Carvalho GM, Brazil RP, Rego FD, Ramos MC, Zenobio AP, Andrade Filho JD. Molecular detection of Leishmania DNA in wild-caught phlebotomine sand flies (Diptera: Psychodidae) from a cave in the State of Minas Gerais. Brazil J Med Entomol. 2017;54:196–203.
Vu SN, Tran HS, Tran VP, Tran CT, Tran ND, Dang DA, et al. Taxonomical insights and ecology of sandfly (Diptera, Psychodidae) species in six provinces of Northern Vietnam. Parasite. 2021;28:85.
Loyer M, Depaquit J, Gay F. A new cavernicolous sand fly from Cambodia: Idiophlebotomus nicolegerae n. sp. (Diptera: Psychodidae). Acta Trop. 2016;155:43–50.
Wang CS, Ku YM, Yuan TC. New records of sandflies in Kweichow province with a description of a new species, Sergentomyia longiforceps sp. nov. Acta Ent Sin. 1974;17:334–8.
Jaturas N, Vitta A, Samung Y, Apiwathnasorn C, Polseela R. Species composition and nocturnal activity of phlebotomine sand flies (Diptera: Psychodidae) inhabiting a limestone cave in Thailand. J Vector Ecol. 2018;43:52–8.
Ilango K. A taxonomic reassessment of the Phlebotomus argentipes species complex (Diptera: Psychodidae: Phlebotominae). J Med Entomol. 2010;47:1–15.
Quate LW, Fairchild GB. Phlebotomus sandflies of Malaya and Borneo. Pac Insects. 1961;3:203–22.
Phuphisut O, Nitatsukprasert C, Pathawong N, Jaichapor B, Pongsiri A, Adisakwattana P, et al. Sand fly identification and screening for Leishmania spp. in six provinces of Thailand. Parasit Vectors. 2021;14:352.
Sinton JA. Notes on some Indian species of the genus Phlebotomus. Part XXIV Phlebotomus barraudi n. sp. Ind J Med Res. 1929;156:716–24.
Sinton JA. Notes on some Indian species of the genus Phlebotomus. Part XXXIV. Phlebotomus iyengari n. sp. Ind J Med Res. 1933;22:221–4.
Raynal J, Gaschen HL. Sur les phlébotomes d’Indochine. VII. Présence de Phlebotomus iyengari, 1933 en Indochine-Nord et description des deux sexes. Bull Soc Pathol Exot. 1935;28:507–17.
Saini P, Shah HK, Jessu M, Sonia T, Anns T, Amju K. Morphological and molecular description of a new species of sandfly, Sergentomyia (Neophlebotomus) ashwanii sp. Nov. (Diptera: Psychodidae) from Western Ghats. India Asian Pac J Trop Med. 2024. https://doi.org/10.4103/apjtm.apjtm_732_23.
Phumee A, Tawatsin A, Thavara U, Pengsakul T, Thammapalo S, Depaquit J, et al. Detection of an unknown Trypanosoma DNA in a Phlebotomus stantoni (Diptera: Psychodidae) collected from Southern Thailand and records of new sand flies with reinstatement of Sergentomyia hivernus Raynal & Gaschen, 1935 (Diptera: Psychodidae). J Med Entomol. 2017;54:429–34.
Raynal J. Sur une nouvelle espèce de phlebotome du nord de la Chine: Phlebotomus khawi n. sp. Ann Parasitol Hum Comp. 1936;14:529–40.
Curler G. Records of phlebotomine sand flies (Diptera, Psychodidae, Phlebotominae) with a description of a new species of Sergentomyia França & Parrot from Khao Yai National Park Thailand. Zootaxa. 2011;2806:60–8.
Raynal J, Gaschen HL. Sur les phlébotomes d’Indochine. IX. Phlebotomus sylvaticus n. sp. Bull Soc Pathol Exot. 1935;28:592–601.
Manalang C. Three new sand-flies from the Philippines. Philipp J Sei. 1931;45:355–65.
Jariyapan N, Tippawangkosol P, Sor-Suwan S, Mano C, Yasanga T, Somboon P, et al. Significance of eggshell morphology as an additional tool to distinguish species of sand flies (Diptera: Psychodidae: Phlebotominae). PLoS ONE. 2022;17:e0263268.
Acknowledgements
We thank the staff of the IP-Laos, Vientiane, for assisting in field sand fly collection. We also thank the staff at the Vientiane Provincial Public Health Office, as well as the District Public Health Office, Lao Ministry of Health, who supported and assisted our field work in Laos.
Funding
Our study was supported by the National Geographic Society (grant number NGS-KOR-50729R-18) and the Institut Pasteur du Laos.
Author information
Authors and Affiliations
Contributions
KV, FJR, PB, and JD: idea conceptualization, study design, data analysis, drafting, critical review, final approval. KV, KL, NP, TP, VV, PL, and SC: field collection, sample preparation, laboratory work, sample identification and critical review. JD: drawing, describing new species. All authors have read, reviewed, and approved the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The study involved the collection of medically important insects from karstic areas of Laos. Local or regional ethics committee approval was not required for such work. Our field collections were performed with the authorization of the Lao Ministry of Health and the local authorities (no. 578).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
13071_2024_6444_MOESM3_ESM.xlsx
Additional file 3: Table S3: The pairwise distance of cytochrome b gene within species and between species found in this study.
13071_2024_6444_MOESM4_ESM.jpg
Additional file 4: Fig. S1: Shannon diversity indices (H) and species accumulation curves for the sand flies examined in this study. Shannon indices by district (A), by trapping position (B), and by season of collection (C). Species accumulation curves by district (D), by trapping position (E) and by season of collections (F).
13071_2024_6444_MOESM5_ESM.jpg
Additional file 5: Fig. S2: The relative abundance of sand flies by district (A) and sampling location (B). Principal coordinate analysis based on the Bray–Curtis index showing the species assemblage between districts (C) and sampling locations (D). PNK Tham Phanokkok cave, TN Tham Nam cave, TP Tham Pha cave, TY Tham Yao cave, PLS Tham Phaluesy cave, NOK Tham Nang Oau Khiem cave, and AL Angluang karstic areas.
13071_2024_6444_MOESM6_ESM.xlsx
Additional file 6: Table S4: Sand fly species composition from different sampling locations from three districts of Vientiane Province in 2019. PNK Tham Phanokkok cave, TN Tham Nam cave, TP Tham Pha cave, TY Tham Yao cave, PLS Tham Phaluesy cave, NOK Tham Nang Oau Khiem cave, and AL Angluang karstic areas.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

{kind=link}
{kind=link}
Cite this article
Vongphayloth, K., Randrianambinintsoa, F.J., Lakeomany, K. et al. A study on the diversity of phlebotomine sand flies (Diptera, Psychodidae) in karstic limestone areas in Vientiane Province, Laos, with a description of two new species of Sergentomyia França and & Parrot. Parasites Vectors 17, 385 (2024). https://doi.org/10.1186/s13071-024-06444-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-024-06444-w