
Abstract
In this paper, we study a predator–prey model with delay and harvesting on predator. We give the conditions for stability and Turing instability of coexisting equilibrium by analyzing the eigenvalue spectrum. By using delay as a bifurcation parameter we give conditions for occurrence of Hopf bifurcation. We investigate the property of bifurcating period solutions by calculating the normal form. We perform some numerical simulations to support our theoretical result. Our results show that diffusion and delay are two factors that should be considered in establishing the predator–prey model, since they can induced the Turing instability and spatially bifurcating period solutions.
Similar content being viewed by others
1 Introduction
Biological population dynamics is an important research area in biological mathematics. In biology there are various interactions between different populations, such as competitive relationship, dependency relationship, predation relationship, and so on. Among them, predation relationship is widespread and studied by many scholars [1,2,3].
Generally, an ordinary differential equation system describing the prey–predator model is
where \(u(t)\) and \(v(t)\) stand for the prey and predator densities. Without predator, the growth law of prey is represented by the function \(\phi (u)\), \(\varphi (u, v)\) is the functional response, α stands for the conversion rate, and θ is the death rate.
The functional response is essential for establishing the predator–prey model. It reflects the predator’s predation ability and can be affected by many factors, such as structure of the habitat structure, hunting ability, prey’s escape ability, and others. In predator–prey models, scholars have used different functional responses to model the interaction of predator and prey; they show that the functional response can enrich the model dynamics [4, 5]. One kind of commonly used functional response functions are Holling type I–III [6], usually called the prey-dependent functional response (with \(\varphi (u,v)\) denoted as \(\varphi (u)\), a function of prey u). Another kind of functional response functions are Beddington–DeAngelis type [7], Crowley–Martin type [8], Hassell–Varley type [9], usually called predator-dependent (with \(\varphi (u,v)\) a function of prey u and predator v).
The Crowley–Martin functional response is of the following form:
where E, S, and B stand for the capture rate of predator to prey, the handling time, and the magnitude of interference among predators, respectively. Cao and Jiang [10] studied a reaction–diffusion type predator–prey model with Crowley–Martin functional response, mainly focusing on Turing–Hopf bifurcation. In [11], the authors studied a predator–prey model with delay and Crowley–Martin functional response, mainly considering the stability and Hopf bifurcation. In [12], the authors considered a predator–prey model with Crowley–Martin functional response, mainly studying the flip bifurcation and Neimark–Sacker bifurcation. These works all suggest that the Crowley–Martin functional response can enrich the dynamics of predator–prey models. In this paper, we mainly study a predator–prey model with Crowley–Martin functional response.
To rationally develop the exploitation of biological resources, many scholars have considered predator–prey models with harvesting. The harvesting can be mainly divided into three types: (i) constant harvesting, (ii) proportional harvesting, and (iii) nonlinear-type harvesting (i.e., the harvesting is a nonlinear function). From a biological and economic perspective, more and more scholars recommend Michaelis–Menten type-harvesting [13,14,15,16], which has the following form:
where Q and E represent the catch ability coefficient and external effort, respectively, and η and β are suitable constants. Constant harvesting and proportional harvesting can be considered as two particular cases of the Michaelis–Menten-type harvesting. In [13], the authors studied the periodic solution of a prey–predator model with harvesting. Yuan et al. [15] studied bifurcation of a delayed predator–prey model with Michaelis–Menten-type prey harvesting. These works suggest that Michaelis–Menten-type harvesting performs well.
Moreover, time delay widely exists in population models. When the predator consumes the prey, it does not immediately increase the density of predator. There exists a gestation delay, and the density of predator increases after some time lag. This type time delay is often studied by scholars [17,18,19,20]. In general, predator–prey models with time delay are much more realistic, and they can exhibit much richer dynamics.
Motivated by these, we studied a diffusive delayed predator–prey model with the following form:
where \(u(x,t)\) and \(v(x,t)\) are the prey and predator densities, respectively, \(D_{1}\) and \(D_{2}\) are for diffusive coefficients, r and K are the growth rate of prey and the carrying capacity, C is the conversion rate of prey, and τ is for the gestation delay of predator. The harvesting term is a Michaelis–Menten-type harvesting on the predator. The main aim of this paper is to study the diffusion-driven Turing instability and delay-induced Hopf bifurcation.
The paper is organized as follows. In Sect. 2, we consider the existence of equilibria of the model. In Sect. 3, we study the stability of the coexisting equilibrium. In Sect. 4, we analyze the property of Hopf bifurcation. In Sect. 5, we give some numerical simulations. Finally, we end the paper with a brief conclusion in Sect. 6.
2 Equilibrium analysis
For convenience, we perform nondimensionalization of model (1.2). Denoting \(\tilde{u}=u/K\), \(\tilde{v}=Ev/r\), and \(\tilde{t}=\mathit{tr}\), system (1.2) becomes (after dropping tildes)
where \(d_{1}=\frac{D_{1}}{r}\), \(d_{2}=\frac{D_{2}}{r}\), \(a=SK\), \(b=\frac{Br}{E}\), \(c=\frac{CEK}{r}\), \(d=\frac{D}{r}\), \(e= \frac{\eta r}{Q}\), and \(q=\frac{\beta r^{2}}{QE^{2}}\). We assume that \(\varOmega =(0,l\pi )\), where \(l>0\).
Solving the equation system
we obtain that \((0,0)\), and \((1,0)\) are two boundary equilibria, and the coexisting equilibrium \((u_{*},v_{*})\) satisfies \(v_{*}=\frac{(1-u_{*}) (1+a u_{*})}{1-b+b(1-a) u_{*}+a b u_{*}^{2}}\) and \(h(u_{*})=0\), where
We just give a sufficient condition for the existence of coexisting equilibrium \((u_{*},v_{*})\):
Theorem 2.1
If the parameters satisfy condition (2.4), then model (2.1) has a coexisting equilibrium \((u_{*},v_{*})\), where \(u_{*}\) is the root of \(h(u_{*})=0\) in the region \((0,1)\), and \(v_{*}=\frac{(1-u_{*}) (1+a u_{*})}{1-b+b(1-a) u_{*}+a b u_{*}^{2}}\).
Proof
By direct calculation we have \(h(0)= (-1-b+b^{2} ) (-1-d (e+q(1-b)))>0\) and \(h(1)=(a+1) (b+1) (1+d (e+q))-c e-c q<0\) under condition (2.4). By the continuity of \(h(u)\) we obtain that \(h(u)=0\) has at least one root \(u_{*}\) in the \((0,1)\). Then \(v_{*}=\frac{(1-u_{*}) (1+a u_{*})}{1-b+b(1-a) u_{*}+a b u_{*}^{2}}>0\). □
3 Stability analysis
Linearize system (2.1) at \((u_{*},v_{*})\):
where
and
The characteristic equation is
where \(I=\operatorname{diag}\{1,1\}\) and \(M_{n}=-n^{2}/l^{2}\operatorname{diag}\{d _{1},d_{2}\}\), \(n \in \mathbb{N}_{0}\). Then we have
where
3.1 The case \(\tau =0\)
When \(\tau =0\), Eq. (3.3) reduces to the equation
where
and the eigenvalues are given by
We make the following hypothesis:
When \(d_{1}=d_{2}=0\) and \(\tau =0\), \((u_{*},v_{*})\) is locally asymptotically stable under hypothesis (\(\mathbf{H_{1}}\)).
Divide the parameters into the following three cases:
Denote
where \(\Delta _{k}\) is defined in (3.5).
Theorem 3.1
Suppose (\(\mathbf{H_{1}}\)) holds and \(\tau =0\).
-
(1)
In Case 1 (or Case 2), \((u_{*},v_{*})\) is locally asymptotically stable;
-
(2)
In Case 3, if \(\mathbb{K}_{1}= \varnothing \), then \((u_{*},v_{*})\) is locally asymptotically stable;
-
(3)
In Case 3, if \(\mathbb{K}_{2} \neq \varnothing \), then \((u_{*},v_{*})\) is Turing unstable.
Proof
Hypothesis (\(\mathbf{H_{1}}\)) implies that \(\mathit{tr}_{0}<0\) and \(\Delta _{0}>0\). For \(n \in \mathbb{N}_{0}\), we have \(\mathit{tr}_{n}<0\). In Case 1 (or Case 2), we have \(\Delta _{n}>0\) for \((n \in \mathbb{N}_{0})\), implying that all eigenvalues of (3.4) have negative real parts. This implies that statement \((1)\) holds. Similarly, statement \((2)\) holds. In Case 3, \(\Delta _{k}<0\) for \(k\in \mathbb{K} _{2}\). Then Eq. (3.4) has a positive real part root. Then statement (3) is true. □
3.2 The case \(\tau \neq 0\)
Next, we study the stability of \((u_{*},v_{*})\) when \(\tau >0\). Letting iω (\(\omega >0\)) be a solution of Eq. (3.3), we have
Then
leading to
Denoting \(z = \omega ^{2}\), we can change (3.8) to
and the roots of (3.9) are
Under condition (1) (or 2) of Theorem (3.1), we have
Denote
Define
and
Lemma 3.1
Assume that (\(\mathbf{H_{1}}\)) holds and the parameters satisfy the condition (1) (or 2) of Theorem 3.1.
-
(1)
For \(n\in \mathbb{S}_{1}\), Eq. (3.3) has a pair of purely imaginary roots \(\pm i\omega ^{+}_{n}\) at \(\tau ^{j,+}_{n}\), \(j \in \mathbb{N}_{0}\).
-
(2)
For \(n\in \mathbb{S}_{2}\), Eq. (3.3) has two pairs of purely imaginary roots \(\pm i\omega ^{\pm }_{n}\) at \(\tau ^{j,\pm }_{n}\), \(j \in \mathbb{N}_{0}\).
-
(3)
For \(n\in \mathbb{S}_{3}\), Eq. (3.3) has no purely imaginary root.
Proof
Equation (3.9) has a (two or no) positive root(s) \({z_{n} ^{+}}\) (or \({z_{n}^{\pm }}\)) when \(n\in \mathbb{S}_{1}\) (\(n\in \mathbb{S}_{2}\) or \(n\in \mathbb{S}_{3}\)). Then statements (1), (2), and (3) are true. □
Lemma 3.2
Assume that (\(\mathbf{H_{1}}\)) holds and the parameters satisfy condition (1) (or 2) of Theorem 3.1. Then \(\operatorname{Re}(\frac{d \lambda }{d \tau })|_{\tau =\tau ^{j,+}_{n}}>0\) and \(\operatorname{Re} ( \frac{d \lambda }{d \tau })|_{\tau =\tau ^{j,-}_{n}}<0\) for \(n \in \mathbb{S}_{1}\cup \mathbb{S}_{2}\) and \(j \in \mathbb{N}_{0}\).
Proof
Differentiating Eq. (3.3) with respect to τ, we obtain
Then
where \(\varLambda =\omega ^{4} b_{2}^{2} +C^{2}_{n} \omega ^{2}>0\). Therefore \(\operatorname{Re}(\frac{d \lambda }{d \tau })|_{\tau =\tau ^{j,+}_{n}}>0\) and \(\operatorname{Re} ( \frac{d \lambda }{d \tau })|_{\tau =\tau ^{j,-}_{n}}<0\). □
From (3.10) we have \(\tau ^{0,\pm }_{n}<\tau ^{j,\pm }_{n}\) \((j\in \mathbb{N})\). For \(n\in \mathbb{S}_{1}\cup \mathbb{S}_{2}\), define \(\tau _{*}=\mbox{min}\{\tau ^{0,\pm }_{n} \mbox{or} \tau ^{0,+} _{n} \mid n \in \mathbb{S}_{1}\cup \mathbb{S}_{2} \}\). By the preceding we obtain the following theorem.
Theorem 3.2
Assume that (\(\mathbf{H_{1}}\)) holds and the parameters satisfy condition (1) (or 2) of Theorem 3.1.
-
(1)
\((u_{*},v_{*})\) is locally asymptotically stable for all \(\tau \geq 0\) when \(\mathbb{S}_{1}\cup \mathbb{S}_{2}=\varnothing \).
-
(2)
\((u_{*},v_{*})\) is locally asymptotically stable for \(\tau \in [0,\tau _{*})\) when \(\mathbb{S}_{1}\cup \mathbb{S}_{2}\neq \varnothing \).
-
(3)
Hopf bifurcation occurs at \((u_{*},v_{*})\) when \(\tau =\tau ^{j,+}_{n}\) \((\tau =\tau ^{j,-}_{n})\), \(j\in \mathbb{N}_{0}\), \(n \in \mathbb{S}_{1}\cup \mathbb{S}_{2}\).
4 Property of Hopf bifurcation
Now, we will study the property of Hopf bifurcation by the method of [21, 22]. For a critical value \(\tau ^{j,+}_{n}\) (or \(\tau ^{j,-}_{n}\)), we denote it as τ̃. Let \(\tilde{u}(x,t)=u(x,\tau t)-u_{*}\) and \(\tilde{v}(x,t)=v(x, \tau t)-v_{*}\). Then system (2.1) is (dropping the tilde)
Denote \(\tau =\tilde{\tau }+\varepsilon \) and \(U=(u(x,t),v(x,t))^{T}\). In the phase space \(\mathscr{C}_{1}:=C([-1,0],X)\), (4.1) can be rewritten as
where \(L_{\varepsilon }(\varphi )\) and \(F(\varphi ,\varepsilon )\) are
and
with
for \(\varphi =(\varphi _{1}, \varphi _{2})^{T} \in \mathscr{C}_{1}\).
We know that \(\varLambda _{n}:=\{i \omega _{n} \tilde{\tau },-i \omega _{n} \tilde{\tau }\}\) are characteristic roots of
By the Riesz representation theorem there exists a \(2\times 2\) matrix function \(\eta ^{n}(s, \tilde{\tau })\) (\(-1\le s \le 0\)) with elements of bounded variation functions such that
for \(\varphi \in C([-1,0],\mathbb {R}^{2})\).
Choose
where
Define the bilinear paring
for \(\varphi \in C([-1,0],\mathbb {R}^{2})\) and \(\psi \in C([0,1], \mathbb {R}^{2})\); \(A(\tilde{\tau })\) has a pair of simple purely imaginary eigenvalues \(\pm i \omega _{n} \tilde{\tau }\), which are also eigenvalues of \(A^{*}\).
Define \(p_{1}(\theta )=(1,\zeta )^{T}e^{i\omega _{n} \tilde{\tau } s} (s \in [-1,0])\) and \(q_{1}(r)=(1,\vartheta )e^{-i\omega _{n} \tilde{\tau } r} (r \in [0,1]) \), where
Let \(\varPhi =(\varPhi _{1},\varPhi _{2})\) and \(\varUpsilon ^{*}=(\varUpsilon ^{*}_{1}, \varUpsilon ^{*}_{2})^{T}\) with
for \(\theta \in [-1,0]\) and
for \(r \in [0,1]\). Then by (4.8) we can compute
Define and construct a new basis ϒ for \(P^{*}\) by
Then \((\varUpsilon ,\varPhi )=I_{2}\). In addition, define \(f_{n}:=(\beta ^{1}_{n},\beta ^{2}_{n})\), where
We also define
and
for \(u=(u_{1},u_{2})\), \(v=(v_{1},v_{2})\), \(u,v\in X\), and \(\langle \varphi ,f _{0}\rangle =(\langle \varphi ,f^{1}_{0}\rangle , \langle \varphi ,f^{2}_{0}\rangle )^{T}\).
Rewrite Eq. (4.1) in the abstract form
where
The solution is
where
and
Then
Let \(z=x_{1}-i x_{2}\) and notice that \(p_{1}=\varPhi _{1}+i\varPhi _{2}\). Then
and
Equation (4.12) becomes
where
and
Let
Then
and
with
Hence
with
and
Denote
Notice that
We have
Then by (4.15), (4.17), and (4.23) we have \(g_{20}=g_{11}=g_{02}=0\) for \(n=1,2,3,\ldots \) . If \(n=0\), then we have
and for \(n\in \mathbb{N}_{0}\), we have \(g_{21}=\tilde{\tau }( \gamma _{1} \kappa _{1} +\gamma _{2} \kappa _{2})\).
From [21] we have
and
where
Hence we have
that is,
Then
Therefore
and
where
By the definition of \(A_{\tilde{\tau }}\) and (4.25) we have
that is,
where
By the definition of \(A_{\tilde{\tau }}\) and (4.25) we have, for \(-1\le \theta <0\),
As
and
we have
that is,
where
Similarly, from (4.26) we have
that is,
Similarly, we have
where
Thus we have:
Theorem 4.1
For any critical value \(\tau ^{j,+}_{n}\) (or \(\tau ^{j,-}_{n}\)), the bifurcating periodic solutions exist for \(\tau >\tau ^{j, \pm }_{n}\) (or \(\tau <\tau ^{j,\pm }_{n}\)) when \(\mu _{2}>0\) (or \(\mu _{2}<0\)) and are orbitally asymptotically stable (or unstable) when \(\beta _{2}<0\) (or \(\beta _{2}>0\)).
5 Numerical simulations
To verify our theoretical results, we give some numerical simulations. Fix the following parameters
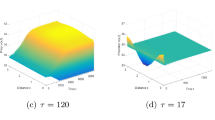
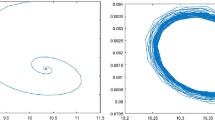
Then \((u_{*},v_{*})=(0.1606, 1.6143)\) is a unique coexisting equilibrium. Hypothesis (\(\mathbf{H_{1}}\)) is satisfied, and the parameters are in Case 1. By calculation we have \(\tau _{*}=\tau ^{0}_{0} \approx 2.3471\). By Theorem 3.1 we have that \((u_{*},v_{*})\) is stable when \(\tau \in [0,\tau _{*})\), which is shown in Fig. 1; \(\tau =\tau _{*}\) is the critical value. When τ crosses it, the stability of \((u_{*},v_{*})\) changes, and bifurcating solution occurs. By calculation we have
Hence the locally asymptotically stable bifurcating periodic solutions appears for \(\tau >2.3471\), which is shown in Fig. 2.

When \(\tau =2\), \((0.1606, 1.6143)\) is asymptotically stable

When \(\tau =2.5\), \((0.1606, 1.6143)\) is is unstable, and stable bifurcating periodic solutions appear
6 Conclusion
We have studied the impact of delay on the dynamics of a diffusive predator–prey model. In this model the functional response is of Crowley–Martin type, and the harvesting of predator is modeled by Michaelis–Menten-type harvesting. We give a sufficient condition (2.4) for coexisting equilibrium to exist. When time delay \(\tau =0\), the stability of coexisting equilibrium is investigated, and the conditions for stability and Turing instability are given in Theorem 3.1. When time delay τ increases, it can affect the stability of coexisting equilibrium and induce Hopf bifurcation. In addition, the property of Hopf bifurcation is considered, including the direction and stability of bifurcating period solutions. Our results suggest that diffusion and time delay are two factors that should be considered in establishing the predator–prey model, since they can induce the Turing instability and spatially bifurcating period solutions.
References
Yi, F., Wei, J., Shi, J.: Bifurcation and spatiotemporal patterns in a homogeneous diffusive predator–prey system. J. Differ. Equ. 246(5), 1944–1977 (2009)
Zhang, T., Meng, X., Song, Y., et al.: A stage-structured predator–prey SI model with disease in the prey and impulsive effects. Math. Model. Anal. 18(4), 505–528 (2013)
Wang, J., Shi, J., Wei, J.: Dynamics and pattern formation in a diffusive predator–prey system with strong Allee effect in prey. J. Differ. Equ. 251, 1276–1304 (2011)
Jana, D., Pathak, R., Agarwal, M.: On the stability and Hopf bifurcation of a prey–generalist predator system with independent age-selective harvesting. Chaos Solitons Fractals 83(83), 252–273 (2016)
Yuan, R., Jiang, W., Wang, Y.: Saddle-node-Hopf bifurcation in a modified Leslie–Gower predator–prey model with time-delay and prey harvesting. J. Math. Anal. Appl. 422(2), 1072–1090 (2015)
Holling, C.S.: The functional response of predators to prey density and its role in mimicry and population regulation. Mem. Entomol. Soc. Can. 97(45), 1–60 (1965)
Beddington, J.R.: Mutual interference between parasites or predators and its effect on searching efficiency. J. Anim. Ecol. 44(1), 331–340 (1975)
Crowley, P.H., Martin, E.K.: Functional responses and interference within and between year classes of a dragonfly population. J. North Am. Benthol. Soc. 8(3), 211–221 (1989)
Hassell, M.P., Varley, G.C.: New inductive population model for insect parasites and its bearing on biological control. Nature 223(5211), 1133–1137 (1969)
Cao, X., Jiang, W.: Turing–Hopf bifurcation and spatiotemporal patterns in a diffusive predator–prey system with Crowley–Martin functional response. Nonlinear Anal., Real World Appl. 43, 428–450 (2018)
Tripathi, J.P., Tyagi, S., Abbas, S.: Global analysis of a delayed density dependent predator–prey model with Crowley–Martin functional response. Commun. Nonlinear Sci. Numer. Simul. 30(1–3), 45–69 (2016)
Ren, J., Yu, L., Siegmund, S.: Bifurcations and chaos in a discrete predator–prey model with Crowley–Martin functional response. Nonlinear Dyn. 90, 19–41 (2017)
Wang, J., Cheng, H., Liu, H., et al.: Periodic solution and control optimization of a prey–predator model with two types of harvesting. Adv. Differ. Equ. 2018(1), 41 (2018)
Krishna, S.V., Srinivasu, P.D.N., Kaymakcalan, B.: Conservation of an ecosystem through optimal taxation. Bull. Math. Biol. 60(3), 569–584 (1998)
Yuan, R., Jiang, W., Wang, Y.: Saddle-node-Hopf bifurcation in a modified Leslie–Gower predator–prey model with time-delay and prey harvesting. J. Math. Anal. Appl. 422(2), 1072–1090 (2015)
Clark, C.W.: Mathematical models in the economics of renewable resources. SIAM Rev. 21(1), 81–99 (2006)
Jiang, Z., Global, W.L.: Hopf bifurcation for a predator–prey system with three delays. Int. J. Bifurc. Chaos 27(7), 1750108 (2017)
Wang, Z., Wang, X., Li, Y., et al.: Stability and Hopf bifurcation of fractional-order complex-valued single neuron model with time delay. Int. J. Bifurc. Chaos 27(13), 1750209 (2017)
Liu, G., Wang, X., Men, X., et al.: Extinction and persistence in mean of a novel delay impulsive stochastic infected predator–prey system with jumps. Complexity 2017(3), 1–15 (2017)
Li, L., Wang, Z., Li, Y., et al.: Hopf bifurcation analysis of a complex-valued neural network model with discrete and distributed delays. Appl. Math. Comput. 330, 152–169 (2018)
Wu, J.: Theory and Applications of Partial Functional Differential Equations. Springer, Berlin (1996)
Hassard, B.D., Kazarinoff, N.D., Wan, Y.H.: Theory and Applications of Hopf Bifurcation. Cambridge University Press, Cambridge (1981)
Acknowledgements
The authors wish to express their gratitude to the editors and the reviewers for the helpful comments.
Funding
This research is supported by the National Nature Science Foundation of China (No. 11601070), Fundamental Research Funds for the Central Universities (No. 2572019BC01), Heilongjiang Provincial Natural Science Foundation (No. A2018001), and Postdoctoral Science Foundation of China (No. 2019M651237).
Author information
Authors and Affiliations
Contributions
The idea of this research was introduced by WG and RY. All authors contributed to the main results and numerical simulations. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Gao, W., Tong, Y., Zhai, L. et al. Turing instability and Hopf bifurcation in a predator–prey model with delay and predator harvesting. Adv Differ Equ 2019, 270 (2019). https://doi.org/10.1186/s13662-019-2211-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-019-2211-4