
Abstract
This paper is related to some dynamical aspects of a class of predator–prey interactions incorporating cannibalism and Allee effects for non-overlapping generations. Cannibalism has been frequently observed in natural populations, and it has an ability to alter the functional response concerning prey–predator interactions. On the other hand, from dynamical point of view cannibalism is considered as a procedure of stabilization or destabilization within predator–prey models. Taking into account the cannibalism in prey population and with addition of Allee effects, a new discrete-time system is proposed and studied in this paper. Moreover, existence of fixed points and their local dynamics are carried out. It is verified that the proposed model undergoes transcritical bifurcation about its trivial fixed point and period-doubling bifurcation around its boundary fixed point. Furthermore, it is also proved that the proposed system undergoes both period-doubling and Neimark–Sacker bifurcations (NSB) around its interior fixed point. Our study demonstrates that outbreaks of periodic nature may appear due to implementation of cannibalism in prey population, and these periodic oscillations are limited to prey density only without leaving an influence on predation. To restrain this periodic disturbance in prey population density, and other fluctuating and bifurcating behaviors of the model, various chaos control methods are applied. At the end, numerical simulations are presented to illustrate the effectiveness of our theoretical findings.
Similar content being viewed by others


1 Introduction
The most interesting and fascinating topic of current research in mathematical biology is the inclusion of Allee effect as well as cannibalism in prey and predator population. The occurrence of Allee effect is the most essential phenomenon in the biological world, and it has been treated as the crucial and extremely significant factor in ecology and population dynamics [1]. Initially, in 1930, the famous ecologist Allee illustrated the Allee effect at low population densities, which has been acknowledged as a prominent factor of positive density dependence in low-density population [2, 3]. The existence of Allee effect represents that it is mandatory for a population to sustain at least a minimum size of population itself in the natural world. There exist numerous populations in the universe, in which Allee effect has been extensively investigated, including insects [4], birds and mammals [5], plants [6], and marine invertebrates [7]. Attention to the interaction among small size populations such as the Allee effect has developed quickly in recent decades [8–12], including predator–prey interaction models. Recently, the development in this era has proved that the inclusion of Allee effect factor in a predator–prey model affects the dynamics of the system and may be the cause of destabilization, but it depends upon where Allee effects are attached [13]. For more interesting dynamical results related to Allee effects, we refer to [14–18] and the references therein.
On the other hand, cannibalism is also an important and intriguing topic in the case of predator–prey interaction, and it plays a key role in the dynamics of such interaction. Among humans, the motivations for cannibalism factor can vary as in human populations it has been documented all around the world. It has been practiced as a social norm in various indigenous South American, African, and New Guinean tribes [19]. It has also been practiced in Northern India among a sect of ascetics or witch doctors “Aghoris” in the hope of achieving immortality. The emphasizing behavior of cannibalism has been observed in a substantial diversity of animals organized as noncarnivorous insects, flour beetles, spider, fish, and locusts [13, 20–23]. Generally, the cannibals and their sufferers are in various maturation stages of life such as adult and teenage, immature and mature, and diverse sorts of categories. This occurrence throws back a predator–prey interaction within the identical species, and the equivalent mathematical models are dissimilar structurally from the predator–prey models only for different species [24, 25]. Polis [25] has tremendous contribution related to cannibalism and has mentioned around thirteen hundred different species involving this factor. In the case of predator–prey interaction, the inclusion of cannibalism is considered as a mechanics of natural selection which is in fact a familiar phenomenon [26]. According to many ecologists and biologists, the behavior of population dynamics has been extremely affected in response to the impact of cannibalism, and these incorporate the lifeboat instrument, where the affected role of cannibalism precedes to perseverance in population destruction [27]. It can also be helpful to get stability in cycling populations [28]. In numerous species, cannibalism factor appears when minimal resources are available corresponding to high level population densities of the species [29]. In the beginning, cannibalism was stimulated as an impact of obstruction in the predator population only, and accordingly it is occasionally competition mediated [28, 30, 31]. Despite ecological support in experimental work as well as in field work, this phenomenon is often observable in prey population [32–34]. However, the experimental work immensely encourages researchers to formulate some innovative ideas in the present scenario. These experimental findings greatly inspire to develop new ideas in current research. A comprehensive study of the present and previous mathematical surveys related to cannibalism indicates that the appearance of cannibalism in population models has many applications. These models comprise ODE, PDE, and discrete models representing two and three species population models, where cannibalism is involved in both prey and predator populations, a ratio-dependent type functional response, and some recent emerging developed models incorporating diseased predators associated with cannibalism [27, 28, 35–39]. In [40] the authors studied a well-known Lotka–Volterra model involving cannibalism in a predator population and discussed the stability analysis of the proposed model. Additionally, these investigations reveal how stability has been affected by cannibalism. Zhang et al. [41] proposed a new method based on non-dimensionalization and applied it on a stage-structure model involving the predator cannibalism factor. The authors also studied the dynamics of the predator–prey model of stage structure including global stability analysis, subcritical and supercritical Hopf bifurcation along with biological meaning of parameters involved in the system.
For the purpose of investigating a class of population models related to non-overlapping generation, Danca et al. [42] discussed the following system:
where \(x_{n}\) and \(y_{n}\) represent prey and predator population respectively with nth generation. Furthermore,
-
r represents intrinsic growth rate of prey;
-
α indicates per capita searching efficiency;
-
d is the conversion rate of predator.
Taking into account the rate of natural death c for predator, system (1) takes the form [43]
Recently, Shabbir et al. [44] discussed the dynamical complexity of model (2) along with prey cannibalism. Moreover, Seval Işık [45] further modified (2) with addition of Allee effect in prey equation and stated it as follows:
where constant term m imposed on the prey equation is known as Allee constant.
At present, some remarkable continued work related to the modification of system (3) including asymptotic stability, bifurcation analysis, and chaos control study has been carried out. Liu [46] examined the existence of periodic solutions for a discrete semi-ratio-dependent predator–prey system. Moreover, the permanence and existence of unique uniformly asymptotic stability of positive almost periodic solutions in a discrete predator–prey system with time delays were determined in [47]. Din [48] considered a Leslie–Gower predator–prey model and studied bifurcation along with feedback control methodologies to control chaos and bifurcation. For further similar fascinating results related to discrete-time predator–prey models, we refer to [49–54] and the references therein.
In this article, our aim is to discuss the dynamics of our proposed model developed from the inclusion of cannibalism in prey population of discrete-time predator–prey model (3) and is expressed by
Clearly, the addition of the term \(\beta \times x\times \frac{x}{x+\gamma }\) is known as a generic cannibalism factor. The cannibalism rate is denoted by β, whereas prey cannibalism has Holling-II type functional response. The term bx represents the birth rate of prey, and the condition \(\beta >b\) is imposed because it takes depredation of prey. Note that the \(x(t)\) population is depredating on its own species.
The rest of this manuscript can be summarized as follows: Sect. 2 deals with the existence of biologically possible equilibria and the conditions of asymptotic stability. Section 3 is associated with the study of bifurcation analysis for system (4). OGY and hybrid control methods are utilized in Sect. 4. Finally, extensive numerical simulations are imposed in Sect. 5 to justify our analytical results.
2 Stability analysis of steady-states
This section is dedicated for the exploration of local stability analysis of system (4). To investigate the solution of system (4), we consider the following algebraic system:
Simple computations yield the following equilibria for system (4):
where
Moreover, trivial equilibrium \(E^{0}\) always exists, the boundary equilibrium \(E^{1}\) exists only for \(k>0\), that is, k is the solution of \(r x^{2} + ( 1+\beta +r\gamma -b-r ) x+\gamma -r\gamma -b\gamma =0\) with \(b+ r>1\), and unique positive equilibrium \(E^{\star }\) exists only for \(\gamma d^{2} + ( 1+c ) ( 1+\beta +\gamma r ) d+ (1+c)^{2} r< ( b+r ) ( 1+c+\gamma d ) d\). The variational matrix at any arbitrary point \((x, y)\) of system (4) is given by
Assume that \(R^{\star }\) is any arbitrary solution of system (4) with Jacobian
and the characteristic polynomial
where \(\mathrm{I} = \alpha _{11} + \alpha _{22}\) and \(\wp = \alpha _{11} \alpha _{22} - \alpha _{12} \alpha _{21}\), then Lemma 1 gives insight about the exploration of local stability analysis of system (4).
Lemma 1
([55])
Let\(\mathcal{P} ( \lambda ) = \lambda ^{2} - I \lambda + \wp \)and\(\mathcal{P} (1)>0\). Moreover, \(\lambda _{1}\), \(\lambda _{2}\)are roots of equation\(\mathcal{P}( \lambda )=0\), then:
-
(i)
\(\vert \lambda _{1} \vert <1\)and\(\vert \lambda _{2} \vert <1\)if and only if\(\mathcal{P}( - 1) >0\)and\(\wp < 1\);
-
(ii)
\(\vert \lambda _{1} \vert >1\)and\(\vert \lambda _{2} \vert >1\)if and only if\(\mathcal{P}( - 1) >0\)and\(\wp > 1\);
-
(iii)
\(\vert \lambda _{1} \vert <1\)and\(| \lambda _{2} | >1\)or (\(| \lambda _{1} | >1\)and\(\vert \lambda _{2} \vert <1\)) if and only if\(\mathcal{P} ( - 1 ) <0\);
-
(iv)
\(\lambda _{1} =-1\)and\(\vert \lambda _{2} \vert \neq 1\)if and only if\(\mathcal{P} ( - 1 ) =0\)and\(\mathrm{I}\neq 0, 2\);
-
(v)
\(\lambda _{1}\), \(\lambda _{2}\)are complex conjugates with\(| \lambda _{1} | =1= \vert \lambda _{2} \vert \)if and only if\(\mathrm{I}^{2} - 4 \wp < 0\)and\(\wp =1 \).
Since \(\lambda _{1}\) and \(\lambda _{2}\) are eigenvalues of system (4), we have elaborated the following topological findings interconnected to the stability of \(R^{\star } \). \(R^{\star }\) is known as sink if \(| \lambda _{1} | <1\) and \(| \lambda _{2} | <1\), as sink is the point of suction which is stable. The equilibrium point \(R^{\star }\) is recognized as source if \(| \lambda _{1} | >1\) and \(| \lambda _{2} | >1\), as it always remains unstable. The equilibrium \(R^{\star }\) is saddle if \(\vert \lambda _{1} \vert >1\) and \(\vert \lambda _{2} \vert <1\) or vice versa (\(\vert \lambda _{1} \vert <1\) and \(\vert \lambda _{2} \vert >1 \)), whereas it is non-hyperbolic if conditions (iv) and (v) from Lemma 1 are fulfilled.
It can be easily observed that the trivial equilibrium \(E^{0} =(0, 0)\) of system (4) has eigenvalues \(b+r\) and −c, then the following assumptions hold:
-
\((0, 0)\) is a sink if and only if \(b+r\in (0, 1)\) and \(c\in ( 0, 1 )\);
-
\((0, 0)\) is a source if and only if \(b+r>1\) and \(c>1\);
-
\((0, 0)\) represents a saddle point if and only if \(b+r>1\) and \(c<1\) and vice versa;
-
\((0, 0)\) is non-hyperbolic for \(b+r=1\) or \(c=1\).
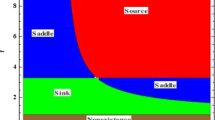
Also, for \(r=0.1\), the topological classification of trivial equilibrium in bc-plane is plotted in Fig. 1(a).

(a) Topological classification for trivial equilibrium for \(r=0.1\). (b) Topological classification of \(E^{1} \). (c) Topological classification for \(E^{\star }\)
Furthermore, at boundary equilibrium point \(E^{1} =(k, 0)\), \(\mathcal{V} ( k, 0 )\) is computed as follows:
Furthermore, the following topological results for boundary equilibrium are satisfied.
Theorem 1
Suppose that\(b+r>1\)and\(\varPsi = \frac{k\beta ( k+2\gamma )}{ ( k+\gamma )^{2}}\), then the following results hold:
-
\(E^{1}\)is a sink ⇔ \(2kr+ \varPsi < b+r+1<2 ( 1+ kr ) + \varPsi\) & \(0< dk<1+c\).
-
\(E^{1}\)is a source ⇔ \(1+ b+r>2 ( 1+ kr ) + \varPsi\) & \(dk>1+c\).
-
\(E^{1}\)is a saddle point ⇔ \(2kr+ \varPsi < b+r+1<2 ( 1+ kr ) + \varPsi \) & \(dk>1+c\).
-
\(E^{1}\)is non-hyperbolic ⇔ \(b+r=1+ 2kr+ \varPsi \)or\(dk = 1+c\).
Taking \(( \beta ,r,c,d ) = ( 1.27,0.5,0.82,1.27 )\), the topological classification for boundary equilibrium is depicted in Fig. 1(b) in bγ-plane.
Moreover, \(\mathcal{V} ( x, y )\) about interior equilibrium \(E^{\star }\) is expressed by
where
The characteristic equation of Jacobian \(\mathcal{V} ( E^{\star } )\) is given by
By performing simple algebraic calculations and letting \(b+r>1\) and \(d ( b+r ) > ( 1+c ) r+d ( 1+\beta +r\gamma )\), we get
From (6), we see that \(\mathcal{F} (1) >0\). Therefore, the following topological classification can be made by applying Lemma 1.
Theorem 2
Assume that\(( 1+c ) ( d ( 1+\beta +r\gamma ) + ( 1+c ) r ) + d^{2} \gamma < d ( b+r ) ( 1+c+d\gamma )\)and put\(1+c+dm=\eta \), \(1+c+d\gamma =\xi \), and\(1+c=\varOmega \)such that the equilibrium point\(E^{\star } = ( x^{\star }, y^{\star } )\)of map (4) exists, then the following findings remain accurate:
-
(a)
\(E^{\star }\)is asymptotically stable ⇔
$$ \left . \textstyle\begin{array}{l} d \varOmega ^{2} ( 1+mr+\xi \beta ( 2+\eta ) ) + d^{2} m ( 1+2 ( b+r ) ) + \xi ^{2} \varOmega ^{2} r ( \varOmega +2 ) \\ \quad < 2d \varOmega ^{2} \beta \eta+ \xi ^{2} \{ d^{2} m ( \varOmega ( 1+b+r ) +5 ) +d \varOmega ( \varOmega ( b+r ) +4 ) \} ; \end{array}\displaystyle \right \} $$and
$$ \left . \textstyle\begin{array}{l} d [ \beta \eta \varOmega ^{2} + \xi ^{2} \{ \varOmega ( 1+b \varOmega ) +(2+ ( b+r ) \varOmega )md \} ] \\ \quad < r( d^{2} m+ \varOmega ^{2} (1+d+ \varOmega ))+d\xi \{ \xi ( \eta +dm ( b+ \varOmega ) ) + \varOmega ^{2} ( 1+\xi +\eta \beta ) \} . \end{array}\displaystyle \right \} $$ -
(b)
\(E^{\star }\)is an unstable equilibrium point ⇔
$$ \left . \textstyle\begin{array}{l} d \varOmega ^{2} ( 1+mr+\xi \beta ( 2+\eta ) ) + d^{2} m ( 1+2 ( b+r ) ) + \xi ^{2} \varOmega ^{2} r ( \varOmega +2 )\\ \quad < 2d \varOmega ^{2} \beta \eta + \xi ^{2} \{ d^{2} m ( \varOmega ( 1+b+r ) +5 ) +d \varOmega ( \varOmega ( b+r ) +4 ) \} \end{array}\displaystyle \right \} $$and
$$ \left . \textstyle\begin{array}{l} d\xi \{ \xi ( \eta +dm ( b+ \varOmega ) ) + \varOmega ^{2} ( 1+\xi +\eta \beta ) \} +r ( d^{2} m+ \varOmega ^{2} ( 1+d+ \varOmega ) )\\ \quad < d [ \beta \eta \varOmega ^{2} + \xi ^{2} \{ \varOmega ( 1+b \varOmega ) +(2+ ( b+r ) \varOmega )md \} ]. \end{array}\displaystyle \right \} $$ -
(c)
\(E^{\star }\)is a saddle point if and only if
$$ \left . \textstyle\begin{array}{l} 2d \varOmega ^{2} \beta \eta + \xi ^{2} \{ d^{2} m ( \varOmega ( 1+b+r ) +5 ) +d \varOmega ( \varOmega ( b+r ) +4 ) \} \\ \quad < d \varOmega ^{2} ( 1+mr+\xi \beta ( 2+\eta ) ) + d^{2} m ( 1+2 ( b+r ) ) + \xi ^{2} \varOmega ^{2} r ( \varOmega +2 ). \end{array}\displaystyle \right \} $$ -
(d)
\(E^{\star }\)is non-hyperbolic ⇔ \(u_{11} + u_{22} \neq 0, 2\)and
$$ r:=- \frac{d\eta ( -5+c+ \frac{2 \varOmega }{\eta } +b ( 1-c- \frac{2 \varOmega }{\eta } ) - \frac{2 \varOmega ^{2} \beta }{\xi ^{2}} + \frac{\varOmega ^{2} (2+\eta )\beta }{\eta \xi } )}{(1+c)^{2} (3+c-d)+d(1+c(2+c-d)+d)m} $$or
$$\begin{aligned}& \vert u_{11} + u_{22} \vert \leq 2, \quad\textit{and} \\& \beta :=- \frac{ ( \varOmega ^{2} ( 2+c ) r-c d^{2} m ( b+r-1 ) - \varOmega ^{2} d ( b+r-mr-1 ) ) \xi ^{2}}{\varOmega ^{2} d ( 1+ ( 1+\eta ) ( c+d\gamma ) )}. \end{aligned}$$
Furthermore, taking \(( \beta ,b,c,d,m ) = ( 4.47,3.07,0.5,3.15,4.4 )\) topological classification for positive equilibrium is depicted in Fig. 1(c).
3 Bifurcation analysis
This section is devoted to the study of bifurcation in which three different types of bifurcations are investigated. We explore transcritical bifurcation, periodic-doubling bifurcation, and Neimark–Sacker bifurcation of system (4) at \(E^{0}\), \(E^{1}\), and \(E^{\star } \).
3.1 Transcritical bifurcation at \({E}^{{0}}\)
In this section, our claim is that fixed point \(E^{0}\) undergoes transcritical bifurcation. Hence we assume that
and consider the set
As \(( r^{0}, b, c, d,m,\alpha , \beta , \gamma ) \in \mathrm{B}_{\mathrm{T}}\), then (4) is alternatively described by the map
where parameter r̂ represents a very small purturbation in \(r^{0} \). Therefore, an application of the Taylor series expansion about \(( x, y, \hat{r} ) =(0, 0, 0)\) yields
where
The linear portion of map (7) is in a canonical form as \(r^{0}:=1-b\). Consequently, the implementation of center manifold \(\mathcal{W}^{c} (0, 0, 0)\) for map (7) is approximated by
where \(h_{1} = h_{2} = h_{3} =0\).
Additionally, we introduce the map restricted to the center manifold \(\mathcal{W}^{c} ( 0, 0, 0 )\):
where \(k_{1} =a-1- \frac{\beta }{\gamma }\), \(k_{2} =1\), \(k_{3} =0\).
Now, here we establish \(\mathcal{L}_{1} \neq 0\) and \(\mathcal{L}_{2} \neq 0\) as follows:
Thus, we can state the following theorem related to transcritical bifurcation.
Theorem 3
Suppose that\(r=1-b\)and\(b-1- \frac{\beta }{\gamma } \neq 0\), then (4) undergoes transcritical bifurcation at its trivial equilibrium\(E^{0} \).
3.2 Period-doubling bifurcation at \({E}^{{1}}\)
In the present section, we study period-doubling bifurcation about \(E^{1} =(k, 0)\). Therefore the Jacobian matrix of corresponding system (4) with respect to \(E^{1}\) is given by
Using Lemma 1 for the case of non-hyperbolic steady-state, when one eigenvalue \(\lambda _{1} =-1\) implies that
we consider the set
Moreover, suppose that \(( r, b, c, d,m,\alpha , \beta , \gamma ) \in \mathrm{B}_{\mathrm{P}_{\mathbb{B}}}\). Then the boundary fixed point \(E^{1}\) of system (4) sustains period-doubling bifurcation whenever β is chosen as a bifurcation parameter and it varies in a small neighborhood of \(\beta _{1} := \frac{(1+b+r-2kr) (k+\gamma )^{2}}{k(k+2\gamma )}\). Therefore, in terms of parameters \(( \beta _{1}, r, b, c, d,m,\alpha , \gamma )\), system (4) can be demonstrated as follows:
After a small perturbation β̃ in \(\beta _{1}\), map (8) can be rearranged as
where \(\vert \tilde{\beta } \vert \ll 1\). Suppose that \(x= \mathrm{H} -k\) and \(y= \mathrm{Z}\), then map (9) is reshaped as follows:
where
Additionally, we establish the following translation map:
where is a nonsingular matrix. Map (11) under translation (10) can be prepared as follows:
where
and \(x:= c_{12} ( u+v )\); \(y:= ( \lambda _{2} -c_{11} ) v- ( 1+ c_{11} ) u\).
In addition, we execute the center manifold \(w^{c} (0, 0, 0)\) for map (12) computed at \((0, 0)\) and within a small neighborhood of \(\tilde{\beta } =0\), then we have
Consequently, the restricted map to the center manifold \(w^{c} ( 0, 0, 0 )\) is given by
and
Now, here we establish once more that \(L_{1}\) and \(L_{2}\) both are nonzero:
The aforementioned analysis yields the following conclusion.
Theorem 4
If\(L_{1} \& L_{2} \neq 0\), then system (4) undergoes period-doubling bifurcation at\(E^{1}\)wheneverβvaries in a small neighborhood of\(\beta _{1} := \frac{(1+b+r-2kr) (k+\gamma )^{2}}{k(k+2\gamma )} \). Moreover, in the case of\(L_{2} >0\), there exist period-two orbits which bifurcate from equilibrium\(E^{1}\), which are stable orbits, whereas for\(L_{2} <0\)unstable orbits are generated.
3.3 Period-doubling bifurcation at \({E}^{{\star }}\)
In this section, we discuss the existence of period-doubling bifurcation in (4) at equilibrium \(E^{\star }\). In addition, the characteristic equation of the Jacobian matrix about \(E^{\star } = ( x^{\star }, y^{\star } )\) is expressed by
Suppose that
and \(\mathcal{F} ( - 1 ) = 0\), then it follows that
From Eq. (13), assume that \(\mathcal{F} ( \lambda ) =0\). Then one root is \(\lambda _{1} =-1\) and the condition \(\vert \lambda _{2} \vert \neq 1\) implies that
Again we consider the following set:
The fixed point \(E^{\star }\) of map (4) undergoes period-doubling bifurcation whenever the parameter r varies in a small neighborhood of the set \(\mathrm{B}_{\mathrm{P}_{\mathbb{U}}} \).
Let
and take the arbitrary parameters \(( r_{1}, b, c, d,m,\alpha , \beta , \gamma ) \in \mathrm{B}_{\mathrm{P}_{\mathbb{U}}}\), then map (4) can be expressed by
Assume a small perturbation r̃ bifurcation parameter. Then map (17) can be examined by
where \(\vert \tilde{r} \vert \ll 1\) is a small perturbation parameter.
Putting \(x=P- x^{*} \) and \(y=Q- y^{*}\), then system (18) is tranformed into the following form:
where
Now, we develop the following map:
where is an invertible matrix. Translation (20) under (19) can be written as follows:
where
and \(x:= a_{12} ( u+v )\); \(y:=- ( 1+ a_{11} ) u+ ( \lambda _{2} -a_{11} ) v\).
Now, considering the center manifold \(\mathbb{W}^{c} (0, 0, 0)\) of (21) in a small neighborhood of \(\tilde{r} =0\), then \(\mathbb{W}^{c} (0, 0, 0)\) can be embellished by
in which
Thus, the map restricted to \(\mathbb{W}^{c} ( 0, 0, 0 )\) is prescribed as follows:
where
Now, here we define two nonzero real numbers as follows:
Due to the aforementioned investigations, we state the following theorem related to period-doubling bifurcation.
Theorem 5
System (4) undergoes period-doubling bifurcation at\(E^{\star }\)when the parameterrchanges its values around a neighboring point of\(r_{1}\)whenever\(\mathbb{L}_{1} \neq 0\)and\(\mathbb{L}_{2} \neq 0\). Furthermore, the period-two orbits that bifurcate from interior equilibrium\(E^{\star }\)are stable whenever\(\mathbb{L}_{2} >0\)and unstable when\(\mathbb{L}_{2} <0\).
3.4 Neimark–Sacker bifurcation at \({E}^{{\star }}\)
This section consists of the existence criteria for Neimark–Sacker bifurcation around an interior fixed point by considering the rate of cannibalism β as a bifurcation parameter. For detailed analysis, we refer to the work done by the authors [56–59]. On the other hand, when Neimark–Sacker bifurcation exists, then as a result, dynamically closed curves appear and attracting steady-states are unstable as varied parameters move towards β. In return, we can discover some isolated orbits along with trajectories and with periodic behavior that thickly overlay these immutable closed curves [60]. In the case of non-hyperbolic fixed points, we have studied the conditions associated with system (4) and a pair of complex eigenvalues having unit modulus. For this, consider (13) and assume that \(\mathcal{F} ( \lambda ) =0\) has two roots which are complex conjugate and fulfill the following conditions:
and
Further, suppose that
Then equilibrium \(E^{\star }\) of (4) undergoes NSB for different parametric values belonging to the small neighborhood of the set \(\mathrm{B}_{\aleph } \).
Let us replace \(\beta _{1} = \beta \) in (22) and \(( r, b, c, d,\alpha , \beta _{1}, \gamma , m ) \in \mathrm{B}_{\aleph } \). Then the following modification in system (4) can be ensured:
Consider the perturbation of map (24) by selecting β̂ as a minimal perturbation parameter, then we have the following map:
where \(\vert \hat{\beta } \vert \ll 1\).
Now, we introduce the transformation and put \(x=P- x^{\star }\) and \(y=Q- y^{\star }\), where \(E^{\star } = ( x^{\star }, y^{\star } )\), then (24) can be rearranged as follows:
where
and \(m_{11}\), \(m_{12}\), \(m_{21}\), \(m_{22}\), \(m_{13}\), \(m_{14}\), \(m_{15}\), \(m_{16}\), and \(m_{23}\) are given in (19) by replacing \(m_{ij} = a_{ij}\), \(i=1, 2\); \(j=1,2,3,4,5,6\) and \(r_{1}\) by r, β by \((\beta _{1} + \hat{\beta } )\). The characteristic equation corresponding to the system of (25) evaluated at (0, 0) is expressed by
where
Since \(( r, b, c, d,\alpha , \beta _{1}, \gamma , m ) \in \mathrm{B}_{\aleph }\), the solutions of (26) are \(\lambda _{1}\) and \(\lambda _{2}\) along with \(\vert \lambda _{1} \vert = \vert \lambda _{2} \vert =1\); consequently one has
and
where \(1+c+dm=\eta \), \(1+c+d\gamma =\xi \), and \(1+c= \varOmega \).
Further, we assume that
Moreover, \(( \beta _{1},r, b, c, d,\alpha , \gamma , m ) \in \mathrm{B}_{\aleph }\) implies that \(-2< \mathcal{p} ( 0 ) <2\). Thus \(\mathcal{p} ( 0 ) \neq \pm 2, 0, -1\) gives \(\lambda _{1}^{n}, \lambda _{2}^{n} \neq 1\) for all \(n=1, 2, 3, 4\) at \(\hat{\beta } =0\). Consequently, the roots of (26) do not occur in the intersection of the unit circle with the coordinate axes when \(\hat{\beta } =0\) and if the following conditions hold:
Now we study the normal form of (26) at \(\hat{\beta } =0\). In order to acquire the normal form, we choose \(\mu = \frac{\mathcal{p} ( 0 )}{2}\), \(\zeta = \frac{1}{2} \sqrt{4 \mathcal{q} ( 0 ) - \mathcal{p}^{2} ( 0 )}\) is attained only if we elaborate the following transformation:
The desired form of (25) under conversion (28) can be reorganized as follows:
\(x= m_{12} u\) and \(y= ( \eta -m_{11} ) u-\zeta v\). Now, we define nonzero \(L \in R( \text{set of real numbers} )\) as follows:
where
and
Due to our aforementioned mathematical study, one can state the following theorem cf. [61–65].
Theorem 6
Assume that (27) holds and\(L\neq 0\), then (4) undergoes NSB at\(E^{\star }\), whenβchanges its values in a small neighborhood of\(\beta _{1} \). Additionally, if\(L<0\), then an attracting invariant closed curve bifurcates from\(E^{\star }\)for\(\beta _{1} < \beta \), and for the case of\(L>0\), a repelling invariant closed curve bifurcates from\(E^{\star }\)for\(\beta _{1} > \beta \).
4 Chaos control
The chaos control and theory of bifurcation is one of the most vital and developed areas of the current research. It has significant characteristics in population models especially models associated with biological species. Furthermore, discrete-time population models are more chaotic and complex as compared to their continuous counterparts. Hence, it is obvious to execute chaos control techniques to evade any uncertainty. The current section consists of the following two feedback control techniques:
-
(i)
OGY feedback control strategy;
-
(ii)
Hybrid feedback control strategy.
First, we apply the OGY method on system (4).
4.1 OGY control method
In this section, we execute the OGY method which was proposed by Ott et al., for details see also [66, 67]. Now, we implement the OGY method on system (4), then we have the following modified form of system (4):
where r is taken as a control parameter. Here, we restrict r to a small interval \(r\in ( r_{0} -\varsigma , r_{0} +\varsigma )\) to achieve the desired control by implementing small perturbations. Also \(r_{0}\) indicates any value from the chaotic region. Moreover, suppose that \(( x^{\star }, y^{\star } )\) is unstable equilibrium of system (4) in the chaotic region under the influence of period-doubling bifurcation. Then, by applying the following linear map, system (29) can be estimated in the neighborhood of \(( x^{\star }, y^{\star } )\) as follows:
where
and
Moreover, controllable system (29) yields that the following matrix
is of rank 2.
Moreover, taking \([ r- r_{0} ] =-K [ \tbinom{x_{n} - x^{\star }}{y_{n} - y^{\star }} ]\), where \(K= [ k_{1} \ k_{2} ]\), then system (30) can be written as
Furthermore, the equivalent controlled system of (4) is stated as follows:
Moreover, the equilibrium point \(( x^{\star }, y^{\star } )\) is asymptotically stable when both eigenvalues of ‘\(J-BK \)’ belong to an open unit disk. The variational matrix ‘\(J - BK \)’ of system (31) can be written as follows:
and the characteristic equation of ‘\(J-BK\)’ is given by
where
4.2 Hybrid control method
To control the chaos which develops due to appearance of bifurcation in system (4), we implement a hybrid control strategy [68]. This strategy was primarily developed for controlling the chaos which developed because of period-doubling bifurcation, but in [69] similar methodology is applied for Neimark–Sacker bifurcation. Moreover, assuming that (4) substantiates bifurcation at \(E^{\star }\), we get the modified controlled system as follows:
where \(\rho \in (0, 1)\) is a controlled parameter. For some applications of the method defined in (32), we refer to the following references [64, 65, 68–71].
Consider the Jacobian of (32) estimated at \(E^{\star }\) and prescribed as follows:
also the characteristic equation is \(\lambda ^{2} - ( \varGamma _{22} + \varGamma _{11} ) \lambda + \mathbb{D} =0\), where
Analogous to the above mathematical computation, we demonstrate the following lemma related to the stability of controlled system.
Lemma 2
The interior equilibrium\(E^{\star }\)of (32) is locally asymptotically stable whenever
5 Numerical simulation and discussion
Example 1
Choosing parameters \(\alpha =1.02\), \(\beta =0.2\), \(\gamma =0.1\), \(m=2.2\), \(b=2.5\), \(c=0.01\), \(d=0.5\), \(r\in [0.9, 1.4]\) and with initial condition \(( x_{0}, y_{0} ) = ( 2.01, 0.6 )\), system (4) undergoes period-doubling bifurcation when \(r\approx 0.9831708806347621 \). Bifurcation diagrams and maximum Lyapunov exponents (MLE) of the corresponding system are depicted in Fig. 2(a), (b), (c). Moreover, system (4) has the unique fixed point \(( x^{\star }, y^{\star } ) = ( 2.02, 0.6279609 )\), and the characteristic equation of the variational matrix calculated at this equilibrium is written as follows:
Furthermore, the roots of (33) are \(\lambda _{1} =-1\) and \(\lambda _{2} =0.8451671696715698\) with \(\vert \lambda _{2} \vert \neq 1\). Hence the parameters
Next, we implement the OGY control strategy to control chaos which appears due to period-doubling bifurcation. For this, taking \(r= 0.9832\), \(\beta =0.2\), \(b=2.5\), \(c=0.01\), \(d=0.5\), \(\alpha = 1.02\), \(\gamma =0.1\), \(m=2.2\), and \(( x^{\star }, y^{\star } ) =(2.02, 0.6279609)\), the equivalent controlled system is given by
Then the Jacobian ‘\(J - BK \)’ of (34) can be evaluated by
Moreover, \(\mathbb{{L}}_{1}\), \(\mathbb{{L}}_{2}\), and \(\mathbb{{L}}_{3}\) represent the marginal stability lines, which are given by
Region bounded by \(\mathbb{{L}}_{1}\), \(\mathbb{L}_{2}\), and \(\mathbb{{L}}_{3}\) represents the stability region and is shown in Fig. 2(d).

Bifurcation and MLE diagrams for system (4) along with parameters \(\alpha =1.02\), \(\beta =0.2\), \(\gamma =0.1\), \(m=2.2\), \(b=2.5\), \(c=0.01\), \(d=0.5\), \(r\in [ 0.9, 1.4 ]\), and \(( x_{0}, y_{0} ) = ( 2.01, 0.6 )\): (a) Bifurcation diagram for \(x_{n}\) (b) Bifurcation diagram for \(y_{n}\) (c) MLE (d) Region of stability for system (34)
Further, assume that \(\alpha =1.02\), \(\beta =0.2\), \(\gamma =0.1\), \(m=2.2\), \(b=2.5\), \(c=0.01\), \(d=0.5\), \(r\in [0.9, 1.4]\) and with initial conditions \(( x_{0}, y_{0} ) = ( 2.01, 0.6 )\), then system (4) undergoes period-doubling bifurcation. To control bifurcation, we stated the corresponding hybrid control system as follows:
The control interval of stability can be seen in Table 1 whenever \(0.9\leq r\leq 1.4\).
Furthermore, for system (35), the bifurcation diagrams are shown in Fig. 4(a) and (b).
Example 2
Let \(r=0.2\), \(b=2.5\), \(c=0.01\), \(d=0.5\), \(\alpha =1.02\), \(\gamma =0.1\), \(\beta \in [0.2, 0.5]\) and with initial conditions \(( x_{0}, y_{0} ) = ( 2.01, 1.7 )\), then in system (4) Neimark–Sacker bifurcation appears when \(\beta \approx 0.4491479133565015 \). On the other hand, the parallel bifurcation diagrams and MLE are dispatched in Fig. 3(a), (b), and (c). Furthermore, system (4) has an interior equilibrium point \(( x^{\star }, y^{\star } ) =(2.02, 1.7778691826151725)\) with characteristic equation calculated at \((2.02, 1.7778691826151725)\) and given by
Furthermore, the roots of (36) are \(\lambda _{1} =0.561640653969324 -0.8273812759598261i\) and \(\lambda _{2} =0.561640653969324 +0.8273812759598261i\) with \(\vert \lambda _{1,2} \vert =1\). Thus the parameters \(( \beta ,r, b, c, d, m,\alpha , \gamma ) = ( 0.4491479133565015, 0.2, 2.5, 0.01, 0.5, 2.2, 1.02, 0.1 ) \in \mathrm{B}_{\aleph } \).

Bifurcation and MLE diagrams of system (4) with parametric values \(r=0.2\), \(b=2.5\), \(c=0.01\), \(d=0.5\), \(\alpha =1.02\), \(\gamma =0.1\), \(\beta \in [ 0.2, 0.5 ]\), and \(( x_{0}, y_{0} ) = ( 2.01, 1.7 )\). (a) Bifurcation diagram for \(x_{n}\) (b) Bifurcation diagram for \(y_{n}\) (c) MLE (d) Stability region for controlled system (37)
Next, we implement the OGY control strategy to control chaos due to appearance of NSB. For this, taking \(r= 0.2\), \(\beta = 0.4491\), \(b=2.5\), \(c=0.01\), \(d=0.5\), \(m=2.2\), \(\alpha =1.02 \gamma =0.1\) and the unique positive equilibrium \(( x^{\star }, y^{\star } ) =(2.02, 1.7779)\), a modified controlled system is given by
Then, the Jacobian matrix ‘\(J - BK \)’ of updated controlled system (37) reduces to
Moreover, the marginal stability lines \(\mathbb{{L}}_{1}\), \(\mathbb{{L}}_{2}\), and \(\mathbb{{L}}_{3}\) are given as follows:
In addition, the triangular region of stability bounded by \(\mathbb{{L}}_{1}\), \(\mathbb{{L}}_{2}\), and \(\mathbb{{L}}_{3}\) is plotted in Fig. 3(d).
Furthermore, we again take \(r=0.2\), \(b=2.5\), \(c=0.01\), \(d=0.5\), \(\alpha =1.02\), \(\gamma =0.1\), \(\beta \in [0.2, 0.5]\) with initial conditions \(( x_{0}, y_{0} ) = ( 2.01, 1.7 ) \). For these values of parameters, system (4) exhibits Neimark–Sacker bifurcation. Now we perform the hybrid control strategy for the purpose of controlling chaos. For the above numeric values, controlled map (32) takes the form
It can be viewed from Table 2 that there exists the control interval of stability when \(0.2\leq \beta \leq 0.5\).
Figure 4(c) and (d) illustrates the bifurcation for controlled system (38).

Bifurcation diagrams for controlled systems (35) and (38). (a) Bifurcation diagrams of prey for controlled system (35) when \(\rho =0.77\). (b) Bifurcation diagrams of predator for controlled system (35) when \(\rho =0.77\). (c) Bifurcation diagrams of prey for controlled system (38) when \(\rho = 0.8860653 \). (d) Bifurcation diagrams of predator for controlled system (38) when \(\rho = 0.8860653 \)
Some phase portraits of system (4) confirm the chaotic behavior of a system where parameter β takes different values from the chaotic region with initial conditions \(( x_{0}, y_{0} ) = ( 2.01, 1.7 )\), while parameters \(( r, b, c, d, m,\alpha , \gamma ) = ( 0.2, 2.5, 0.01, 0.5, 2.2, 1.02, 0.1 )\) remain the same for each case. Finally, interesing local implication diagrams and the plot of the controlled system are plotted in Fig. 6.
6 Concluding remarks
Intra-specific predation or cannibalism is a significant natural process that controls population dynamics. It has immense complex consequences on population dynamics [24, 72]. Considering the Allee effect and cannibalism on prey population, a discrete-time system for predator–prey interaction is proposed and investigated. It is analyzed that model (4) has three steady-states \(E^{0}\), \(E^{1}\), \(E^{\star }\). The linearization technique is utilized to achieve the local stability of the steady-states. Furthermore, if \(\gamma d^{2} + ( 1+c ) ( 1+\beta +\gamma r ) d+ ( 1+c )^{2} r< ( b+r ) ( 1+c+\gamma d ) d\), then system (4) has a unique positive steady-state \(E^{\star }\). The topological classification of \(E^{0}\), \(E^{1}\), and \(E^{\star }\) is shown in Fig. 1(a), (b), and (c). It is proved that system (4) undergoes transcritical bifurcation at \(E^{0}\), period-doubling bifurcation at \(E^{1}\), and around \(E^{\star }\) there exist both period-doubling bifurcation and Neimark–Sacker bifurcation by using bifurcation theory. Also, our theoretical results are supported by some figures. In Fig. 2(a), a bifurcation diagram of system (4) is depicted for various parametric values of \(\alpha =1.02\), \(\beta =0.2\), \(\gamma =0.1\), \(m=2.2\), \(b=2.5\), \(c=0.01\), \(d=0.5\), \(r\in [0.9, 1.4]\) and the initial condition \(( x_{0}, y_{0} ) = ( 2.01, 0.6 ) \). We observe that \(E^{\star }\) is stable for \(r< 0.9831708806347621\) and loses its stability at \(r= 0.9831708806347621\), and the system undergoes period-doubling bifurcation when the growth rate of prey r exceeds the value 0.9831708806347621. In Figs. 3(a) and (b), the bifurcation diagrams of system (4) are shown. Now, by taking β (cannibalism rate of prey) as a bifurcation parameter with different values of parameters \(r=0.2\), \(b=2.5\), \(c=0.01\), \(d=0.5\), \(\alpha =1.02\), \(\gamma =0.1\), \(\beta \in [0.2, 0.5]\) and the initial condition \(( x_{0}, y_{0} ) = ( 2.01, 1.7 )\), the unique steady-state \(E^{\star } =(2.02, 1.7778691826151725)\) of system (4) is stable for \(\beta > 0.4491479133565015\) and loses its stability at \(\beta = 0.4491479133565015\), also an attracting invariant closed curves appear when \(\beta < 0.4491479133565015\). In Fig. 5, some phase portraits are plotted for different values of β, which shows the chaotic and complex behavior of the system. Moreover, Figs. 2(d), 3(d), and 4 ensure that our proposed control strategies successfully control the bifurcation. Ultimately, we can say that for predator–prey interaction, the growth rate of prey and the cannibalism rate of prey both have remarkable consequences for the stability of system (4) and for population models.

Phase portraits of system (4) for different values of bifurcation parameter β

Local amplification of system (4) and the plot for controlled map (35) when \(\beta =0.2112\) and \(\rho =0.89 \): (a) Local amplification of prey population (b) Local amplification of predator population (c) MLE for local amplification (d) Plot of \(x_{n}\) (e) Plot of \(y_{n}\) (f) Phase portrait for map (35)
References
Dennis, B.: Allee effects, population growth, critical density, and the chance of extinction. Nat. Resour. Model. 3(4), 481–538 (1989)
Allee, W.C.: Cooperation Among Animals. Henry Shuman, New York (1951)
Allee, W.C., Bowen, E.: Studies in animal aggregations mass protection against colloidal silver among goldfishes. J. Exp. Zool. 61(2), 185–207 (1932)
Kuussaari, M., Saccheri, I., Hanski, I.: Allee effect and population dynamics in the glanville fritillary butterfly. Oikos 82(2), 384–392 (1998)
Courchamp, F., Grenfell, B., Clutton-Brock, T.: Impact of natural enemies on obligately cooperatively breeders. Oikos 91(2), 311–322 (2000)
Ferdy, J.B., Austerlitz, F., Moret, J., Gouyon, P.H., Godelle, B.: Pollinator-induced density dependence in deceptive species. Oikos 87(3), 549–560 (1999)
Stoner, A., Ray-Culp, M.: Evidence for Allee effects in an over-harvested marine gastropod, density dependent mating and egg production. Mar. Ecol. Prog. Ser. 202, 297–302 (2000)
Allen, L., Fagan, J., Fagerholm, H.: Population extinction in discrete-time stochastic population models with an Allee effect. J. Differ. Equ. Appl. 11(4–5), 273–293 (2005)
Dennis, B.: Allee effects in stochastic populations. Oikos 96(3), 389–401 (2002)
Jang, S.R.J.: Allee effects in a discrete-time host–parasitoid model. J. Differ. Equ. Appl. 12(2), 165–181 (2006)
Morozov, A., Petrovskii, S., Li, B.L.: Bifurcations and chaos in a predator–prey system with the Allee effect. Proc. R. Soc. Lond. B 271(1546), 1407–1414 (2004)
Zhou, S., Liu, Y., Wang, G.: The stability of predator–prey systems subject to the Allee effects. Theor. Popul. Biol. 67(1), 23–31 (2005)
Wise, D.H.: Cannibalism, food limitation, intraspecific competition, and the regulation of spider populations. Annu. Rev. Entomol. 51, 441–465 (2006)
Berec, L., Angulo, E., Multiple, C.F.: Allee effects and population management. Trends Ecol. Evol. 22(4), 185–191 (2007)
Courchamp, F., Clutton-Brock, T., Grenfell, B.: Inverse density dependence and the Allee effect. Trends Ecol. Evol. 14(10), 405–410 (1999)
Courchamp, F., Berec, L., Gascoigne, J.: Allee Effects in Ecology and Conservation. Oxford University Press, London (2008)
Mooring, M.S., Fitzpatrick, T.A., Nishihira, T.T., Reisig, D.D.: Vigilance, predation risk and the Allee effect in desert bighorn sheep. J. Wildl. Manag. 68(3), 519–532 (2004)
Kangalgil, F.: Neimark–Sacker bifurcation and stability analysis of a discrete-time prey–predator model with Allee effect in prey. Adv. Differ. Equ. 2019(1), 92, 1–12 (2019)
Pennell, C.: Cannibalism in early modern North Africa. Br. J. Middle East. Stud. 18(2), 169–185 (1991)
Claessen, D., De Roos, A.M.: Bistability in a size-structured population model of cannibalistic fish a continuation study. Theor. Popul. Biol. 64(1), 49–65 (2003)
Guttal, V., Romanczuk, P., Simpson, S.J., Sword, G.A., Couzin, I.D.: Cannibalism can drive the evolution of behavioral phase polyphenism in locusts. Ecol. Lett. 15(10), 1158–1166 (2012)
Lioyd, M.: Self-regulation of adult numbers by cannibalism in two laboratory strains of flour beetles (Tribolium castaneum). Ecology 49(2), 245–259 (1968)
Richardson, M.L., Mitchell, R.F., Reagel, P.F., Hanks, L.M.: Causes and consequences of cannibalism in noncarnivorous insects. Annu. Rev. Entomol. 55, 39–53 (2010)
Fox, L.R.: Cannibalism in natural populations. Annu. Rev. Ecol. Syst. 6, 87–106 (1975)
Polis, G.A.: The evolution and dynamics of intraspecific predation. Annu. Rev. Ecol. Syst. 12, 225–251 (1981)
Claessen, D., De Roos, A.M., Persson, L.: Population dynamic theory of size-dependent cannibalism. Proc. R. Soc. Lond. B 271(1537), 333–340 (2004)
Getto, P., Diekmann, O., De Roos, A.: On the (dis)advantages of cannibalism. J. Math. Biol. 51(6), 695–712 (2005)
Kohlmeier, C., Ebenhoh, W.: The stabilizing role of cannibalism in a predator–prey system. Bull. Math. Biol. 57(3), 401–411 (1995)
Pizzatto, L., Shine, R.: The behavioral ecology of cannibalism in cane toads (Bufo marinus). Behav. Ecol. Sociobiol. 63(1), 123–133 (2008)
Fasani, S., Rinaldi, S.: Remarks on cannibalism and pattern formation in spatially extended prey–predator systems. Nonlinear Dyn. 67(4), 2543–2548 (2012)
Sun, G.Q., Zhang, G., Jin, Z., Li, L.: Predator cannibalism can give rise to regular spatial pattern in a predator–prey system. Nonlinear Dyn. 58, 75–84 (2009)
Rudolf, V.H.: Consequences of stage-structured predators: cannibalism, behavioral effects, and trophic cascades. Ecology 88(12), 2991–3003 (2007)
Rudolf, V.H.: The interaction of cannibalism and omnivory: consequences for community dynamics. Ecology 88(11), 2697–2705 (2007)
Rudolf, V.H.: The impact of cannibalism in the prey on predator–prey systems. Ecology 89(6), 3116–3127 (2008)
Biswas, S., Chatterjee, S., Chattopadhyay, J.: Cannibalism may control disease in predator population: result drawn from a model based study. Math. Methods Appl. Sci. 38(11), 2272–2290 (2015)
Buonomo, B., Lacitignola, D., Rionero, S.: Effect of prey growth and predator cannibalism rate on the stability of a structured population model. Nonlinear Anal., Real World Appl. 11, 1170–1181 (2010)
Buonomo, B., Lacitignola, D.: On the stabilizing effect of cannibalism in stage-structured population models. Math. Biosci. Eng. 3(4), 717–731 (2006)
Basheer, A., Quansah, E., Bhowmick, S., Parshad, R.D.: Prey cannibalism alters the dynamics of Holling–Tanner-type predator–prey models. Nonlinear Dyn. 85(4), 2549–2567 (2016)
Basheer, A., Parshad, R.D., Quansah, E., Yu, S., Upadhyay, R.K.: Exploring the dynamics of a Holling–Tanner model with cannibalism in both predator and prey population. Int. J. Biomath. 11(1), 1850010 (2018)
Deng, H., Chen, F., Zhu, Z., Li, Z.: Dynamic behaviors of Lotka–Volterra predator–prey model incorporating predator cannibalism. Adv. Differ. Equ. 2019, 359, 1–17 (2019)
Zhang, F., Chen, Y., Li, J.: Dynamical analysis of a stage-structured predator–prey model with cannibalism. Math. Biosci. 307, 33–41 (2019)
Danca, M., Codreanu, S., Bako, B.: Detailed analysis of a nonlinear prey–predator model. J. Biol. Phys. 23, 11–20 (1997)
Rana, S.M.S.: Bifurcation and complex dynamics of a discrete-time predator–prey system. Comput. Ecol. Softw. 5(2), 187–200 (2015)
Shabbir, M.S., Din, Q., Alabdan, R., Tassaddiq, A., Ahmad, K.: Dynamical complexity in a class of novel discrete-time predator–prey interaction with cannibalism. IEEE Access 8, 100226–100240 (2020)
Seval, I.: A study of stability and bifurcation analysis in discrete-time predator–prey system involving the Allee effect. Int. J. Biomath. 12(1), 1950011 (2019)
Liu, X.: A note on the existence of periodic solutions in discrete predator–prey models. Appl. Math. Model. 34(9), 2477–2483 (2010)
Li, Y., Zhang, T., Ye, Y.: On the existence and stability of a unique almost periodic sequence solution in discrete predator–prey models with time delays. Appl. Math. Model. 35(11), 5448–5459 (2011)
Din, Q.: Complexity and chaos control in a discrete-time prey–predator model. Commun. Nonlinear Sci. Numer. Simul. 49, 113–134 (2017)
Gámez, M., Lopez, I., Rodrıguez, C., Varga, Z., Garay, J.: Ecological monitoring in a discrete-time prey–predator model. J. Theor. Biol. 429, 52–60 (2017)
Huang, J., Liu, S., Ruan, S., Xiao, D.: Bifurcations in a discrete predator–prey model with nonmonotonic functional response. J. Math. Anal. Appl. 464, 201–230 (2018)
Weide, V., Varriale, M.C., Hilker, F.M.: Hydra effect and paradox of enrichment in discrete-time predator–prey models. Math. Biosci. 310, 120–127 (2019)
Shabbir, M.S., Din, Q., Safeer, M., Khan, M.A., Ahmad, K.: A dynamically consistent nonstandard finite difference scheme for a predator–prey model. Adv. Differ. Equ. 2019(1), 381, 1–17 (2019)
Din, Q., Shabbir, M.S., Khan, M.A., Ahmad, K.: Bifurcation analysis and chaos control for a plant–herbivore model with weak predator functional response. J. Biol. Dyn. 13(1), 481–501 (2019)
Chow, Y., Jang, S.R.: Cannibalism in discrete-time predator–prey systems. J. Biol. Dyn. 6, 38–62 (2012)
Liu, X., Xiao, D.: Complex dynamic behaviors of a discrete-time predator–prey system. Chaos Solitons Fractals 32(1), 80–94 (2007)
Din, Q.: Qualitative analysis and chaos control in a density-dependent host–parasitoid system. Int. J. Dyn. Control 6(3), 778–798 (2018)
Din, D., Hussain, M.: Controlling chaos and Neimark–Sacker bifurcation in a host–parasitoid model. Asian J. Control 21(3), 1202–1215 (2019)
He, Z., Lai, X.: Bifurcation and chaotic behavior of a discrete-time predator–prey system. Nonlinear Anal., Real World Appl. 12(1), 403–417 (2011)
Jing, Z., Yang, J.: Bifurcation and chaos in discrete-time predator–prey system. Chaos Solitons Fractals 27, 259–277 (2006)
Din, Q.: Bifurcation analysis and chaos control in discrete-time glycolysis models. J. Math. Chem. 56(3), 904–931 (2018)
Guckenheimer, J., Holmes, P.: Nonlinear Oscillations, Dynamical Systems, and Bifurcations of Vector Fields. Springer, New York (1983)
Robinson, C.: Dynamical Systems: Stability, Symbolic Dynamics and Chaos. CRC Press, Boca Raton (1999)
Wiggins, S.: Introduction to Applied Nonlinear Dynamical Systems and Chaos. Springer, New York (2003)
Wan, Y.H.: Computation of the stability condition for the Hopf bifurcation of diffeomorphism on \(\mathrm{R}^{2}\). SIAM J. Appl. Math. 34(1), 167–175 (1978)
Kuznetsov, Y.A.: Elements of Applied Bifurcation Theory. Springer, New York (1997)
Ott, E., Grebogi, C., Yorke, J.A.: Controlling chaos. Phys. Rev. Lett. 64, 1196–1199 (1990)
Lynch, S.: Dynamical Systems with Applications Using Mathematica. Birkhäuser, Boston (2007)
Luo, X.S., Chen, G., Wang, B.H., Fang, J.Q.: Hybrid control of period-doubling bifurcation and chaos in discrete nonlinear dynamical systems. Chaos Solitons Fractals 18(4), 775–783 (2003)
Yuan, L.G., Yang, Q.G.: Bifurcation, invariant curve and hybrid control in a discrete-time predator–prey system. Appl. Math. Model. 39, 2345–2362 (2015)
Khan, M.A., Shabbir, M.S., Din, Q., Ahmad, K.: Chaotic behavior of harvesting Leslie–Gower predator–prey model. Comput. Ecol. Softw. 9(3), 67–88 (2019)
Khan, M.S., Khan, M.A., Shabbir, M.S., Din, Q.: Stability, bifurcation and chaos control in a discrete-time prey–predator model with Holling type-II response. Netw. Biol. 9(3), 58–77 (2019)
Magnusson, K.: Destabilizing effect of cannibalism on a structured predator–prey system. Math. Biosci. 155, 61–75 (1999)
Acknowledgements
We would appreciate the editors and referees for their valuable comments and suggestions to improve our paper.
Availability of data and materials
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
All authors contributed equally to the writing of this paper. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
It is declared that none of the authors have any competing interests in this manuscript.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Shabbir, M.S., Din, Q., Ahmad, K. et al. Stability, bifurcation, and chaos control of a novel discrete-time model involving Allee effect and cannibalism. Adv Differ Equ 2020, 379 (2020). https://doi.org/10.1186/s13662-020-02838-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-020-02838-z