
Abstract
A stochastic two-prey one-predator system with distributed delays is proposed in this paper. Firstly, applying the linear chain technique, we transform the predator–prey system with distributed delays to an equivalent system with no delays. Then, by use of the comparison method and the inequality technique, we investigate the stability in mean and extinction of species. Further, by constructing some suitable functionals, using M-matrix theory and three important lemmas, we establish sufficient conditions assuring the existence of distribution and the attractivity of solutions. Finally, some numerical simulations are given to illustrate the main results.
Similar content being viewed by others

1 Introduction
Predator–prey system is very popular in the world. In order to reveal the dynamical relationship between predator and prey, a lot of predator–prey systems have been widely investigated and many good results have been obtained in the last decades, which has long been one of the hot topics in ecology [1–3]. Since two-species ecological models cannot describe the natural phenomena accurately and many vital behaviors can only be exhibited by systems with three or more species, for example, in the natural world, the predator often feeds on some competing prey, and hence, a three- or multi-species population system attracts more and more attention [4–7].
On the other hand, all species are inevitably affected by environmental noise. To better describe ecological phenomena, the white noise is introduced into a predator–prey model to reveal richer and more complex dynamics [8–15]. There are many kinds of stochastic perturbation. Considering the stochastic influence on the intrinsic growth rates of populations, we have \(a_{i}\rightarrow a_{i} +\xi _{i} \,d{\omega } (t)\), where \(\omega (t)\) is a standard Brownian motion defined on a complete probability space \((\Omega , \mathcal{{F} },\mathcal{{P})}\) with a filtration \(\{{\mathcal{{F}}}_{t}\}_{t\geq 0} \), \(\xi ^{2}\) is the intensity of white noise. For example, Liu [6] proposed the following three-species predator–prey model:
where \(N_{1}(t)\) and \(N_{2}(t)\) are the population sizes of prey species, \(N_{3}(t)\) is the population size of predator species, \(a_{i}>0\) (\(i=1,2\)) are the intrinsic rates of increase, \(a_{3}<0\) is the intrinsic rate of decrease, \(d_{12}>0\) and \(d_{21}>0\) are the parameters representing competitive effects between two prey, \(d_{13}>0\) and \(d_{23}>0\) are the coefficients of decrease of prey species due to predation, \(d_{31}<0\) and \(d_{32}<0\) are the predation rate of predator, \(d_{ii}>0\) (\(i=1,2,3\)) are the rate of competition within the same species.
As we know, predator–prey interaction is a frequently observed phenomenon. Almost all species should exhibit some delays. Considering the inevitability, more and more researchers have taken delay into an ecological model and obtained some nice results [16–19]. Recently, infinite delay has been widely introduced into the ecological model since the works of Volterra to translate the cumulative effect of the past history of a system [20–24]. Chen [22] et al. proposed the following model with distributed delays:
where the kernel \(K:[0,\infty )\rightarrow [0,\infty )\) is a normalized \(L^{1}\) function such that \(\int _{0}^{\infty }K(s)\,ds=1\). For distributed delay, MacDonald [25] initially proposed that it was reasonable to use gamma distribution as a kernel function, that is, \(f(t)=\frac{t^{n}\sigma ^{n+1}e^{-\sigma t}}{n!}\), where \(\sigma >0, n\) is a nonnegative integer. If \(n=0\), then the kernel \(f(t)=\sigma e^{-\sigma t}\) is called a weak kernel, otherwise it is called a strong kernel.
Motivated by the above discussion, in this paper, we consider a stochastic two-prey one-predator system with distributed delays. For convenience, we mainly consider the weak kernel case, i.e., \(f(t)=\sigma e^{-\sigma t}\). Our model is as follows:
with the initial data
where \(C((-\infty ,0],R_{+})\) is the set of all continuous functions from \((-\infty ,0)\) to \(R_{+}=(0,\infty )\), \(\omega _{i}(t)\) (\(i=1,2,3\)) is a standard and independent Brownian motion defined as above. All parameters are positive constants and their biological meanings refer to [6].
Define
Computing the derivative of \(y_{i}(t)\), then \(dy_{i}(t)=\sigma _{i}(N_{i}(t)-y_{i}(t))\,dt\), \(i=1,2,3\). Using the linear chain technique to (1.1) yields
According to the equivalent property of (1.1) and (1.2), in what follows, we mainly consider (1.2) to reveal the dynamical properties of (1.1). Our main aims are as follows.
Firstly, we study the stability in mean and extinction of all species of (1.2), which have long been and will still be two important topics for the study of stochastic population systems.
Secondly, for a stochastic population system, instead of the positive equilibrium state of the determinate system, it is interesting and important to study the existence and uniqueness of the distribution of (1.2).
The rest work of this paper is organized as follows. Section 2 begins with some notations, definitions, and important lemmas. Section 3 focuses on the stability in mean and extinction of species of (1.2). Section 4 is devoted to the existence and uniqueness of distribution. Some numerical simulations are given in Sect. 5. Finally, we conclude the paper with a brief conclusion and discussion in Sect. 6.
2 Preliminaries
For simplicity, we give the following notations.
Throughout this paper, we denote the complement minor of \(a_{ij}\) in determinant A by \(A_{ij}\) (\(i,j=1,2,3\)), and assume that \(A>0\), \(A_{i}>0\), i.e., when there is no stochastic perturbation, a positive equilibrium state exists for model (1.1). Further, for convenience, we always assume that K stands for a generic positive constant whose value may be different at different places. And for any function \(x(t)\), \(t>0\), we denote
Now we give assumptions, definitions, and some important lemmas, which are used in our main proof.
Assumption 2.1
\(A_{13}>0\), \(A_{23}<0\), \(A_{33}>0\), \(A_{31}<0\), \(A_{32}>0 \).
Assumption 2.2
\(a_{ii}>\sum_{j=1,j\neq i}^{3}a_{ji}\), \(i,j=1,2,3\).
Remark 2.1
Assumption 2.2 means that the intra-specific competitive rates are stronger than the interaction competitive rates or predation rates among different species.
Definition 2.1
Let \(P(t)=(N_{1}(t),N_{2}(t),N_{3}(t),y_{1}(t),y_{2}(t),y_{3}(t))^{T} \in C((-\infty ,0],R_{+}^{6})\) be a solution of system (1.2), then
-
(I)
The population \(P(t)\) is said to be extinct if \(\lim_{t\rightarrow \infty }P(t)=0\);
-
(II)
The population \(P(t)\) is said to be stable in mean if \(\lim_{t\rightarrow \infty }\langle P(t)\rangle =K\), a.s., where K is a constant.
Definition 2.2
Let \(P(t)=(N_{1}(t),N_{2}(t),N_{3}(t),y_{1}(t),y_{2}(t),y_{3}(t))^{T}\in C((-\infty ,0], R_{+}^{6})\) and \(\bar{P}(t)=(\bar{N}_{1}(t),\bar{N}_{2}(t),\bar{N}_{3}(t),\bar{y}_{1}(t), \bar{y}_{2}(t),\bar{y}_{3}(t))^{T} \in C((-\infty ,0],R_{+}^{6})\) be any two positive solutions of (1.2) with the initial value \(P(0)>0\), \(\bar{P}(0)>0\), then system (1.2) is said to be globally attractive if
Lemma 2.1
([26])
Suppose that \(Z(t)\in C[\Omega \times [0,+\infty ), R_{+}] \) and \(\lim_{t\rightarrow \infty }F(t)/t=0\), a.s.
-
(a)
If there exist two positive constants \(T>0\), \(\lambda _{0}>0\) such that, for all \(t>T\),
$$\begin{aligned}& \ln Z(t)\leq \lambda t-\lambda _{0} \int _{0}^{t}z(s)\,ds+F(t), \quad \textit{a.s.}, \\& \textit{then}\quad \textstyle\begin{cases} \langle Z\rangle ^{*}\leq \lambda /\lambda _{0}, &\textit{a.s.}, \textit{if } \lambda \geq 0, \\ \lim_{t\rightarrow +\infty } Z(t)= 0, & \textit{a.s.}, \textit{if } \lambda < 0. \end{cases}\displaystyle \end{aligned}$$ -
(b)
If there exist some constants \(T>0\), \(\lambda _{0}>0,\lambda \) such that, for all \(t>T\),
$$\begin{aligned}& \ln Z(t)\geq \lambda t-\lambda _{0} \int _{0}^{t}z(s)\,ds+F(t), \quad \textit{a.s.}, \\& \textit{then} \quad \langle Z\rangle _{*}\geq \lambda /\lambda _{0}, \quad \textit{a.s.} \end{aligned}$$
Lemma 2.2
System (1.2) has a unique solution \(P(t)=(N_{1}(t),N_{2}(t),N_{3}(t),y_{1}(t), y_{2}(t), y_{3}(t))^{T}\in C((-\infty ,0],R_{+}^{6})\) for any given initial data \(P(t_{0})\in C((-\infty ,0],R_{+}^{6})\), almost surely.
Proof
The proof is standard. For the readers’ convenience, we give the proof in Appendix A.1.
As to the expectation boundedness and asymptotical properties of the solution of (1.2), we have the following lemma. The proof is similar to that of references [20, 27, 28] and is presented in Appendix A.2. □
Lemma 2.3
Let \(P(t)=(N_{1}(t),N_{2}(t),N_{3}(t),y_{1}(t),y_{2}(t),y_{3}(t))^{T}\) be the solution of (1.2), then for any initial data \(P(t_{0})\in C((-\infty ,0],R_{+}^{6})\), there exists a positive constant \(K(p)\) such that
further,
For the following integral equation
there is a result as follows.
Lemma 2.4
([29])
Suppose that the coefficients of (2.1) are independent of t and satisfy:
in \(U_{R}\) for any \(R>0\), and there exists a nonnegative \(C^{2}\) function \(V(x)\) in \(R^{l}\) such that
outside some compact set, then system (2.1) has a solution, which is a stationary Markov process.
Lemma 2.5
([30])
Let \(f(t)\) be a nonnegative function defined on \([0,+\infty )\) such that \(f(t)\) is integrable on \([0,+\infty )\) and is uniformly continuous on \([0,+\infty )\), then \(\lim_{t\rightarrow \infty }f(t)=0\).
Lemma 2.6
Let \(P(t)=(N_{1}(t),N_{2}(t),N_{3}(t),y_{1}(t),y_{2}(t),y_{3}(t))^{T}\) be a solution of (1.2) with the initial value \(P(0)>0\), then almost every sample path of \(P(t)\) is uniformly continuous on \(t\geq 0\).
Proof
For the first equation of (1.2), it is equivalent to the following stochastic integral equation:
By computation, we have
Using the moment inequality for stochastic integrals, for any \(0\leq t_{1}\leq t_{2}\), \(p>2\), we have
In the same manner, we can discuss the following five equations of (1.2) and obtain similar inequalities as above. Therefore, by Lemma 2.4 of Refs. [31], we conclude that almost every sample path of \(P(t)\) is uniformly continuous. The proof is completed. □
3 Stability in mean and extinction of species
Firstly, we give the following result on stability in mean and extinction of species of model (1.2).
Theorem 3.1
If Assumptions 2.1and 2.2hold, then for system (1.2), we have:
-
(i)
If \(b_{1}<0\), \(b_{2}<0\), then \(\lim_{t\rightarrow \infty }N_{i}(t)=0\), \(i=1,2,3\);
-
(ii)
If \(b_{1}<0\), \(b_{2}>0\), \(\Delta _{1}<\tilde{\Delta }_{1}\), then
$$ \lim_{t\rightarrow \infty }N_{1}(t)=0, \qquad \lim_{t \rightarrow \infty } \bigl\langle N_{2}(t) \bigr\rangle =\frac{b_{2}}{a_{22}}, \qquad \lim _{t\rightarrow \infty } N_{3}(t)=0; $$If \(b_{1}<0\), \(b_{2}>0\), \(\Delta _{1}>\tilde{\Delta }_{1}\), then
$$ \lim_{t\rightarrow \infty }N_{1}(t)=0,\qquad \lim_{t \rightarrow \infty } \bigl\langle N_{2}(t) \bigr\rangle = \frac{\Delta _{1}^{*}-\tilde{\Delta }_{1}^{*}}{A_{11}},\qquad \lim _{t\rightarrow \infty } \bigl\langle N_{3}(t) \bigr\rangle = \frac{\Delta _{1}-\tilde{\Delta }_{1}}{A_{11}}; $$ -
(iii)
If \(b_{1}>0\), \(b_{2}<0\), \(\Delta _{2}<\tilde{\Delta }_{2}\), then
$$ \lim_{t\rightarrow \infty } \bigl\langle N_{1}(t) \bigr\rangle = \frac{b_{1}}{a_{11}},\qquad \lim_{t\rightarrow \infty } N_{2}(t)=0,\qquad \lim _{t\rightarrow \infty } \bigl\langle N_{3}(t) \bigr\rangle =0; $$If \(b_{1}>0\), \(b_{2}<0\), \(\Delta _{2}>\tilde{\Delta }_{2}\), then
$$ \lim_{t\rightarrow \infty }N_{2}(t)=0, \qquad \lim_{t \rightarrow \infty } \bigl\langle N_{1}(t) \bigr\rangle = \frac{\Delta _{2}^{*}-\tilde{\Delta }_{2}^{*}}{A_{22}},\qquad \lim _{t\rightarrow \infty } \bigl\langle N_{3}(t) \bigr\rangle = \frac{\Delta _{2}-\tilde{\Delta }_{2}}{A_{22}}; $$ -
(iv)
If \(b_{1}>0\), \(b_{2}>0\), \(A_{i}>\tilde{A}_{i}\) (\(i=1,2,3\)), then
$$ \lim_{t\rightarrow \infty } \bigl\langle N_{1}(t) \bigr\rangle = \frac{A_{1}-\tilde{A}_{1}}{A},\qquad \lim_{t\rightarrow \infty } \bigl\langle N_{2}(t) \bigr\rangle =\frac{A_{2}-\tilde{A}_{2}}{A},\qquad \lim_{t\rightarrow \infty } \bigl\langle N_{3}(t) \bigr\rangle = \frac{A_{3}-\tilde{A}_{3}}{A}; $$If \(b_{1}>0\), \(b_{2}>0\), \(A_{3}<\tilde{A}_{3}\), \(\Delta _{3}>\tilde{\Delta }_{3}\), then
$$ \lim_{t\rightarrow \infty } \bigl\langle N_{3}(t) \bigr\rangle =0,\qquad \lim_{t\rightarrow \infty } \bigl\langle N_{1}(t) \bigr\rangle = \frac{\Delta _{3}^{*}-\tilde{\Delta }_{3}^{*}}{A_{33}},\qquad \lim_{t\rightarrow \infty } \bigl\langle N_{2}(t) \bigr\rangle = \frac{\Delta _{3}-\tilde{\Delta }_{3}}{A_{33}}; $$
Proof
For (1.2), integrating the forth to the sixth equations from 0 to t leads to
Taking the limit as \(t\rightarrow \infty \), combining with Lemma 2.3, we have
By utilizing Itô’s formula to \(\ln N_{i}(t)\) (\(i=1,2,3\)) and integrating both sides of the first three equations of (1.2) from 0 to t, we obtain
Denote \(\xi _{i}(t)\omega _{i}(t)=\vartheta _{i}(t)\), then
and
We begin with the proof of (i).
It follows from (3.2) and (3.3) that
and
Using Lemma 2.1 to (3.5) and (3.6), then
Since \(b_{3}<0\), (3.4) implies \(\lim_{t\rightarrow \infty }N_{3}(t)=0\), and hence
Now we prove (ii).
By the proof of (i), if \(b_{1}<0\), then \(\lim_{t\rightarrow \infty }N_{1}(t)=0\), and hence (3.3) and (3.4) imply that
and
By the elimination method, adding (3.7) applied by \(a_{33}\) and (3.8) applied by \(-a_{23}\) gives
Applying Lemma 2.1 and Lemma 2.3 to (3.9), we get
Substituting \(\langle N_{2}(t)\rangle ^{*}\) into (3.8), we obtain
and
If \(b_{1}<0\), \(b_{2}>0\), \(\Delta _{1}<\tilde{\Delta }_{1}\), then (3.10) implies \(\lim_{t\rightarrow \infty } N_{3}(t) =0\). By use of (3.7) again, we have
Applying Lemma 2.1 to (3.11) gives
Similarly, we have
and \(\langle N_{2}(t)\rangle _{*}\geq \frac{b_{2}}{a_{22}}\), and hence
If \(b_{1}<0\), \(b_{2}>0\), \(\Delta _{1}>\tilde{\Delta }_{1}\), then we can derive from (3.7) and (3.8) that
and
Using Lemma 2.1 to (3.12) yields
Applying Lemma 2.1 to (3.15) yields
Then we have
Therefore, case (ii) is proved. The proof of case (iii) is similar to case (ii) and we omit it here.
Next we enter the proof of case (iv). We begin to eliminate \(\langle N_{1}(t)\rangle \), \(\langle N_{2}(t)\rangle \) from (3.2)–(3.4) by the elimination method. By analysis, there exist positive constants \(p=A_{13}/A_{33}>0\), \(q=-A_{23}/A_{33}>0\), multiplying both sides of (3.2)–(3.4) by p, q, and 1, respectively, adding the three inequalities yields
Similarly, by the elimination method, there exist constants
such that
and
Using Lemma 2.3 in equality (3.16), for arbitrarily \(\varepsilon >0\), there exists \(T>0\), for all \(t>T\), we have
Substituting (3.19) into (3.16) leads to
Since \(A_{3}>\tilde{A}_{3}\), letting \(\varepsilon >0\) be small enough such that \(A_{3}-\tilde{A}_{3}-\varepsilon >0\), then by Lemma 2.1, we have
Similarly, we derive from (3.2) and (3.3) that
and
Applying Lemma 2.1 to (3.21) and (3.22) again, for sufficiently large t, we obtain
By the definition of sup limit, we deduce from (3.4) that
Therefore, Lemma 2.1 implies
By the same way, from (3.2) and (3.3), we obtain
and
Substituting \(\langle N_{1}(t)\rangle ^{*}\leq \frac{A_{1}-\tilde{A}_{1}}{A}\), \(\langle N_{2}(t)\rangle ^{*}\leq \frac{A_{2}-\tilde{A}_{2}}{A}\), \(\langle N_{3}(t)\rangle ^{*} \leq \frac{A_{3}-\tilde{A}_{3}}{A}\) into (3.23) and (3.24) and using Lemma 2.1, we have
Therefore,
which is the required assertion.
If \(b_{1}>0\), \(b_{2}>0\), \(A_{3}<\tilde{A}_{3}\), then the proof is similar to case (iii), and we omit it here. The proof is completed. □
Remark 3.1
By the process of our proof, if considering the effect of Lévy jumps, one can also establish sufficient conditions preserving the stability in mean and extinction of all species. Here we move some restricting conditions like \(R>0\) and \(b_{1}>b_{2}\), which appeared in [6].
4 Stability in distribution
Theorem 4.1
The solution of model (1.2) is a stationary Markov process, that is, there exists a stationary distribution for system (1.2) if Assumption 2.2holds.
Proof
Define
where \(R_{1}\), \(R_{2}\), \(R_{3}\) are positive constants defined later. By Itô’s formula, we have
By Assumption 2.2, there exist positive constants
such that
Define \(\breve{V}=\hat{V}+\sum_{i=1}^{3}\frac{y_{i}^{p+1}}{2\sigma _{i}}\), then
Let
Applying Itô’s formula to \(\tilde{V}(t)\) yields
Define \(V(t)=\breve{V}(t)+\tilde{V}(t)\), then
where \(\sigma =\sigma _{1}+\sigma _{2}+\sigma _{3}\), and
Choose \(\varepsilon >0\) small enough such that
Define the following bounded closed set:
and for \(i=1,2,3\), denote
Denote the complement of \(D_{\varepsilon }\) by \(D_{\varepsilon }^{C}\), then it is easy to get \(D_{\varepsilon }^{C}=\bigcup_{j=1}^{12} D_{\varepsilon }^{j}\). For all \((N_{1},N_{2},N_{3},y_{1},y_{2},y_{3})\in D_{\varepsilon }^{C}\), we discuss as follows.
-
(i)
If \((N_{1},N_{2},N_{3},y_{1},y_{2},y_{3})\in D_{\varepsilon }^{i}\), \(i=1,2,3\), then
$$ LV\leq \sigma +M-\frac{1}{4}N_{i}^{p}\leq \sigma +M- \frac{1}{4\varepsilon ^{p+1}}< -1; $$ -
(ii)
If \((N_{1},N_{2},N_{3},y_{1},y_{2},y_{3})\in D_{\varepsilon }^{i+3}\), \(i=1,2,3\), then
$$ LV\leq \sigma +M-\frac{1}{4}y_{i}^{p}\leq \sigma +M- \frac{1}{4\varepsilon ^{p+1}}< -1; $$ -
(iii)
If \((N_{1},N_{2},N_{3},y_{1},y_{2},y_{3})\in D_{\varepsilon }^{i+6}\), \(i=1,2,3\), then
$$ LV\leq \sigma +M-\iota \frac{r_{i}-\frac{\xi _{i}^{2}}{2}(\iota +1)}{N_{i}^{\iota }}\leq \sigma +M-\iota \frac{r_{i}-\frac{\xi _{i}^{2}}{2}(\iota +1)}{{\varepsilon }^{\iota }} < -1; $$ -
(iv)
If \((N_{1},N_{2},N_{3},y_{1},y_{2},y_{3})\in D_{\varepsilon }^{i+9}\), \(i=1,2,3\), then
$$ LV\leq \sigma +M-\sigma _{i} N_{i}/y_{i}\leq \sigma +M-\sigma _{i} \varepsilon /\varepsilon ^{2} < -1. $$
Consequently, for any \((N_{1},N_{2},N_{3},y_{1},y_{2},y_{3})\in D_{\varepsilon }^{C}\), we have
Therefore, it follows from Lemma 2.4 that there exists a stationary distribution for system (1.2). The proof is completed. □
Theorem 4.2
Under Assumption 2.2, solutions of model (1.2) are globally attractive.
Proof
Firstly, let \(N(t)=N(t,N(\phi ))\) and \(\bar{N}(t)=\bar{N}(t,\bar{N}(\phi ))\) be any two solutions of model (1.1) with the initial data \(N(\phi ), \bar{N}(\phi )\in C([-\tau ,0],R_{+}^{3})\). We only need to prove \(\lim_{t\rightarrow \infty }\mathbb{ E}|N_{i}(t)-\bar{N}_{i}(t)|=0\) for \(i=1,2,3\).
Define
where \(D_{i}\) (\(i=1,2,3\)) is defined later in the proof. By computing the upper right derivative of \(V(t)\), then
On the other hand, by (1.2), we have
that is,
Therefore,
Integrating two sides of the above inequality from 0 to t, we have
Integrating both sides of (4.1) from 0 to t and taking expectations give
For the following equations,
under Assumption 2.2, the coefficient matrix of \(D_{1}\), \(D_{2}\), and \(D_{3}\) is a nonsingular M-matrix, then by M-matrix theory, there exists \(D_{i}>0\) (\(i=1,2,3\)) satisfying the equation. Therefore,
which means \(|N_{i}(t)-\bar{N}_{i}(t)|\in \mathtt{L}_{1}[0,+\infty )\). Consequently, we can derive from Lemma 2.5 and Lemma 2.6 that
The proof is completed. □
Remark 4.1
Combining the existence of distribution and the global attractivity of solutions of (1.2), we conclude that system (1.2) has a unique distribution, which is stable.
5 Numerical simulations
In this section, we give some numerical simulations to validate our theoretical results. By the Milstein higher order method proposed by Higham [32], we numerically simulate the solutions of system (1.2). Using discretization Brownian path over \([0,T]\) and writing efficient Matlab codes, we can obtain the corresponding simulation figures one by one.
Let
In the following, without special mention, we only change the parameter of white noise and keep the rest of parameters unchanged so as to clearly see the dynamical effect of white noise.
Case (i) \(b_{1}<0\), \(b_{2}<0\).
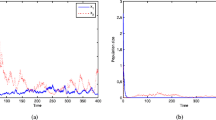
Let \(\xi _{1}=0.8\), \(\xi _{2}=0.8\), \(\xi _{3}=0.5292\), then an easy computation yields \(b1=-0.01\), \(b_{2}=-0.01\), \(b_{3}=-0.15\). It follows from Theorem 3.1 that all species are extinct, illustrated in Fig. 1.

Extinction of all species of system (1.2) with \(\xi _{1}=0.8\), \(\xi _{2}=0.8\), \(\xi _{3}=0.5292\). (a) Extinction of \(N_{1}\), \(N_{2}\), \(N_{3}\). (b) Extinction of \(y_{1}\), \(y_{2}\), \(y_{3}\)
Case (ii) \(b_{1}<0\), \(b_{2}>0\).
Let \(\xi _{1}=0.8\), \(\xi _{2}=0.4\), \(\xi _{3}=0.01\), then \(b1=-0.01\), \(b_{2}=0.23\), \(b_{3}=-0.01\), and \(\Delta _{1}^{*}=0.019\), \(\tilde{\Delta }_{1}^{*}=0.093\), \(A_{11}=0.17\), \(\frac{\Delta _{1}-\tilde{\Delta }_{1}}{A_{11}}=0.019/0.17=0.1118\), \(\frac{\Delta _{1}^{*}-\tilde{\Delta }_{1}^{*}}{A_{11}}=0.093/0.17=0.5471\). By Theorem 3.1, then
which is illustrated in Fig. 2(a).

Dynamical behavior of (1.2). (a) is for (1.2) with \(\xi _{1}=0.8\), \(\xi _{2}=0.4\), \(\xi _{3}=0.01\), then \(\lim_{t\rightarrow \infty }N_{1}(t)=0\), \(\lim_{t \rightarrow \infty }\langle N_{2}(t)\rangle =0.5471\), \(\lim_{t \rightarrow \infty }\langle N_{3}(t)\rangle =0.1118\), (b) is for (1.2) with \(r_{3}=0.1\), \(\xi _{1}=0.8\), \(\xi _{2}=0.469\), \(\xi _{3}=0.01\), then \(N_{1}(t)\), \(N_{3}(t) \) are both extinct, \(\lim_{t\rightarrow \infty }\langle N_{2}(t)\rangle =0.5\)
If \(r_{3}=0.1\), \(\xi _{1}=0.8\), \(\xi _{2}=0.469\), \(\xi _{3}=0.01\), then \(b1=-0.01\), \(b_{2}=0.2\), \(b_{3}=-0.1\), \(b_{2}a_{32}+b_{3}a_{22} =-0.02<0\). Hence, Theorem 3.1 implies \(N_{1}(t)\), \(N_{3}(t) \) are both extinct, and species \(N_{2}(t)\) is stable in mean, and \(\lim_{t\rightarrow \infty }\langle N_{2}(t)\rangle =0.5\), see Fig. 2(b).
Case (iii) \(b_{1}>0\), \(b_{2}<0\).
Let \(\xi _{1}=0.4\), \(\xi _{2}=0.8\), \(\xi _{3}=0.01\), then \(b1=0.23\), \(b_{2}=-0.01\), \(b_{3}=-0.01\). By computation, \(\Delta _{2}-\tilde{\Delta }_{2} =0.0305\), \(\Delta _{2}^{*}-\tilde{\Delta }_{2}^{*}=0.0935\), \(A_{22}=0.1825\). Hence, it follows from Theorem 3.1 that \(N_{2}(t)\) is extinct, and
Figure 3(a) verifies it correctly.

Dynamical behavior of (1.2). (a) is for system (1.2) with \(\xi _{1}=0.4\), \(\xi _{2}=0.8\), \(\xi _{3}=0.01\), then \(N_{2}(t)\) is extinct, and \(\lim_{t\rightarrow \infty }\langle N_{1}(t)\rangle =0.1671\), \(\lim_{t\rightarrow \infty }\langle N_{3}(t)\rangle =0.5123\). (b) is for system (1.2) with \(r_{3}=0.1\), \(\xi _{1}=0.469\), \(\xi _{2}=0.8\), \(\xi _{3}=0.01\), then \(N_{2}(t)\), \(N_{3}(t)\) are both extinct and \(\lim_{t\rightarrow \infty }\langle N_{1}(t)\rangle =0.5\)
If \(r_{3}=0.1\), \(\xi _{1}=0.469\), \(\xi _{2}=0.8\), \(\xi _{3}=0.01\), then \(b1=0.2\), \(b_{2}=-0.01\), \(b_{3}=-0.1\), and \(b_{1}a_{31}+b_{3}a_{11}-0.01\). Theorem 3.1 indicates that \(N_{2}(t)\), \(N_{3}(t)\) are both extinct and \(\lim_{t\rightarrow \infty }\langle N_{1}(t)\rangle =0.2/0.4=0.5\), see Fig. 3(b).
Case (iv) \(b_{1}>0\), \(b_{2}>0\).
We choose \(\xi _{1}=0.1414\), \(\xi _{2}=0.1414\), \(\xi _{3}=0.2\) such that \(b1=0.3\), \(b_{2}=0.3\), \(b_{3}=-0.03\). By computation, then \(A_{1}-\tilde{A}_{1} =0.036\), \(A_{2}-\tilde{A}_{2} =0.0266\), \(A_{3}- \tilde{A}_{3}=0.0153 \). Therefore, by Theorem 3.1, we have
which is illustrated in Fig. 4.

Stable case of (1.2) with \(\xi _{1}=0.1414\), \(\xi _{2}=0.1414\), \(\xi _{3}=0.2\). Theorem 3.1 shows that \(\lim_{t\rightarrow \infty }\langle N_{1}(t)\rangle = \frac{A_{1}-\tilde{A}_{1}}{A}=0.5581\), \(\lim_{t\rightarrow \infty }\langle N_{2}(t)\rangle =\frac{A_{2}-\tilde{A}_{2}}{A}=0.4124\), \(\lim_{t\rightarrow \infty }\langle N_{3}(t)\rangle = \frac{A_{3}-\tilde{A}_{3}}{A}=0.2372\)
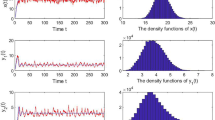
By use of Theorems 4.1 and 4.2, we know that system (1.2) has a unique distribution, which is revealed in Figs. 5 and 6. Figure 5 is the probability density function of preys \(N_{1}(t)\), \(N_{2}(t)\) and predator \(N_{3}(t)\), respectively. Figure 6 shows the attractivity of the solutions. They both indicate the existence and stability of stationary distribution function. The simulation results verify that when the condition is satisfied, that is, the white noise is relatively small, system (1.2) is stable.

Probability density function of preys \(N_{1}(t)\), \(N_{2}(t)\) and predator \(N_{3}(t)\) of (1.2) with \(\xi _{1}= 0.1414\), \(\xi _{2}=0.1414\), \(\xi _{3}=0.2\)

Attractivity of solutions of system (1.2)
If \(\xi _{1}=0.1414\), \(\xi _{2}=0.1414\), \(\xi _{3}=0.5292\), then \(b1=0.3\), \(b_{2}=0.3\), \(b_{3}=-0.15\). By computation, we have \(A_{3}-\tilde{A}_{3}=-0.0015<0 \), \(\Delta _{3}-\tilde{\Delta }_{3}=0.06\), \(\Delta _{3}^{*}-\tilde{\Delta }_{3}^{*}=0.09\), \(A_{33}=0.14 \), which guarantees that the condition of case (iv) holds, and hence Theorem 3.1 implies the two-prey are stable in mean and the predator \(N_{3}\) is extinct, further,
and
Figure 7 indicates the result is true.

Dynamical behavior of (1.2) with \(\xi _{1}=0.1414\), \(\xi _{2}=0.1414\), \(\xi _{3}=0.5292\). Theorem 3.1 implies the two prey are stable in mean and the predator \(N_{3}\) is extinct, and \(\lim_{t\rightarrow \infty }\langle N_{1}(t)\rangle = \frac{\Delta _{3}-\tilde{\Delta }_{3}}{A_{33}}=0.4286\), \(\lim_{t \rightarrow \infty }\langle N_{2}(t)\rangle = \frac{\Delta _{3}^{*}-\tilde{\Delta }_{3}^{*}}{A_{33}}=0.6429\), \(\lim_{t\rightarrow \infty }\langle N_{3}(t)\rangle =0\)
6 Conclusion and discussion
In this paper, we consider a three-species stochastic predator–prey system with distributed delays. Theorem 3.1 gives sufficient conditions of the stability in mean and extinction of each species. Theorems 4.1 and 4.2 give the existence and uniqueness of distribution of each species. Finally, by numerical simulations, we illustrate the validity of our theoretical results.
Theorem 3.1 implies that stochastic parameter \(\xi _{i}\) (\(i=1,2,3\)) has some important influences to the extinction, stability in mean of all species of (1.2), which is illustrated by our simulations clearly. Simulations reveal that small intensity of white noise strengthens the stability of (1.2), while large intensity of white noise will bring serious influence to the dynamical behavior.
Recently, regime switching appears in a biological system frequently, and many nice results have been obtained by many researchers. How about the white noise affecting the dynamical behavior of a predator–prey system with regime switching? We believe it is very interesting and leave it for our future work.
Availability of data and materials
Data sharing not applicable to this article as no datasets were generated or analysed during the current study.
References
Rudnicki, R., Pichór, K.: Influence of stochastic perturbation on prey-predator systems. Math. Biosci. 206, 108–119 (2007)
Shao, Y.: Globally asymptotical stability and periodicity for a nonautonomous two-species system with diffusion and impulses. Appl. Math. Model. 36, 288–300 (2012)
Shao, Y., Li, Y.: Dynamical analysis of a stage structured predator-prey system with impulsive diffusion and generic functional response. Appl. Math. Comput. 220, 472–481 (2013)
Wu, J.: Stability of a three-species stochastic delay predator-prey system with Lévy noise. Physica A 502, 492–505 (2018)
Hastings, A., Powell, T.: Chaos in a three-species food chain. Ecology 72, 896–903 (1991)
Liu, M., Bai, C., Deng, M., Du, B.: Analysis of stochastic two-prey one-predator model with Lévy jumps. Physica A 445, 176–188 (2016)
Liu, M.: Population dynamical behaviour of a two-predator one-prey stochastic model with time delays. Discrete Contin. Dyn. Syst. 37, 2513–2538 (2017)
Qi, H., Meng, X.: Threshold behavior of a stochastic predator-prey system with prey refuge and fear effect. Appl. Math. Lett. 113, 106846 (2021)
Liu, G., Qi, H., Chang, Z., Meng, X.: Asymptotic stability of a stochastic May mutualism system. Comput. Math. Appl. 79(3), 735–745 (2020)
Feng, T., Meng, X., Zhang, T., Qiu, Z.: Analysis of the predator-prey interactions: a stochastic model incorporating disease invasion. Qual. Theory Dyn. Syst. 19(2), 55 (2020)
Zhao, Y., Yuan, S., Zhang, T.H.: Stochastic periodic solution of a non-autonomous toxic-producing phytoplankton allelopathy model with environmental fluctuation. Commun. Nonlinear Sci. Numer. Simul. 44, 266–276 (2017)
Tran, K., Yin, G.: Stochastic competitive Lotka–Volterra ecosystems under partial observation: feedback controls for permanence and extinction. J. Franklin Inst. 351, 4039–4064 (2014)
Li, X., Mao, X.: Population dynamical behavior of non-autonomous Lotka–Volterra competitive system with random perturbation. Discrete Contin. Dyn. Syst. 24(2), 523–545 (2009)
Du, N.H., Sam, V.H.: Dynamics of a stochastic Lotka–Volterra model perturbed by white noise. J. Math. Anal. Appl. 324, 82–97 (2006)
Mao, X.: Stationary distribution of stochastic population systems. Syst. Control Lett. 60, 398–405 (2011)
Kuang, Y.: Delay Differential Equations with Applications in Population Dynamics. Academic Press, New York (1993)
Liu, Q.: The effects of time-dependent delays on global stability of stochastic Lotka–Volterra competitive model. Physica A 420, 108–115 (2015)
Hu, Y., Wu, F., Huang, C.: Stochastic Lotka–Volterra models with multiple delays. J. Math. Anal. Appl. 375, 42–57 (2011)
Liu, M., Bai, C.: Optimal harvesting of a stochastic logistic model with time delay. J. Nonlinear Sci. 25, 277–289 (2015)
Jiang, D., Hayat, T., Zuo, W., Alsaedib, A., Sun, X.: Long-time behaviors of a stochastic cooperative Lotka–Volterra system with distributed delay. Physica A 506, 542–559 (2018)
Chen, L.J., Chen, F.D.: Dynamic behaviors of the periodic predator-prey system with distributed time delays and impulsive effect. Nonlinear Anal., Real World Appl. 12, 2467–2473 (2011)
Chen, L., Song, X., Lu, Z.: Mathematical Ecology Models and Research Methods. Sichuan Science and Technology Press, Chengdu (2003)
Macdonald, N.: Time Lags in Biological Models. Lecture Notes in Biomathematics. Springer, Heidelberg (1978)
Ruan, S.: Delay differential equations in single species dynamics. In: Arino, O., et al.(eds.) Delay Differential Equations and Applications, pp. 477–517. Springer, New York (2006)
Blyuss, K.B., Kyrychko, Y.N.: Stability and bifurcations in an epidemic model with varying immunity period. Bull. Math. Biol. 72, 490–505 (2010)
Liu, M., Wang, K., Wu, Q.: Survival analysis of stochastic competitive models in polluted environment and stochastic competitive exclusion principle. Bull. Math. Biol. 73, 1969–2012 (2011)
Zhang, X.H., Li, W.X., Li, M., Wang, K.: Dynamics of a stochastic Holling II one-predator two-prey system with jumps. Physica A 421, 571–582 (2015)
Huang, L.C.: Stochastic delay population systems. Appl. Anal. 88, 1303–1320 (2009)
Khas’minskii, R.Z.: Stochastic Stability of Differential Equations. Sijthoff Noordhoff, Alphen aan den Rijn, Netherlands (1980)
Barbalat, I.: Systems d’equations differential d’oscillations nonlinearies. Rev. Roum. Math. Pures Appl. 4, 267–270 (1959)
Xu, D.Y., Huang, Y.M., Yang, Z.G.: Existence theorem for periodic Markov process and stochastic functional differential equations. Discrete Contin. Dyn. Syst. 24, 1005–1023 (2009)
Higham, D.J.: An algorithmic introduction to numerical simulation of stochastic differential equations. SIAM Rev. 43, 525–546 (2001)
Mao, X., Marion, G., Renshaw, E.: Environmental Brownian noise suppresses explosions in populations dynamics. Stoch. Process. Appl. 97, 95–110 (2002)
Acknowledgements
We are very grateful to the anonymous referees for their careful reading and very valuable comments, which led to improvement of our paper.
Funding
This work is supported partially by the Natural Science Foundation of China (Grant No. 11861027) and Inner Mongolia Natural Science Foundation (Grant No. 2018MS01017 and No. 2021MS01005).
Author information
Authors and Affiliations
Contributions
JZ carried out all studies and drafted the manuscript. YS conceived of the study and participated in its design and coordination and helped to draft the manuscript, YS performed the simulation analysis. All authors read and approved the final manuscript. All authors contributed equally to the writing of this paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Appendix
Appendix
1.1 1.1 Proof of Lemma 2.2
Let \(N_{i}(t)=e^{x_{i}(t)}\), \(y_{i}(t)=e^{M_{i}(t)}\), then (1.2) is transformed to the following equivalent system:
Clearly the coefficients of (A.1) obey the local Lipschitz condition, then it has a unique local solution on \([0, \tau _{e})\), where \(\tau _{e}\) is the explosion time. According to Itô’s formula, we can see that \(N_{i}(t)=e^{x_{i}(t)}\), \(y_{i}(t)=e^{M_{i}(t)}\) (\(i=1,2,3\)) is the unique positive local solution of (1.2). So we only need to prove \(\tau _{e}=\infty \). To this end, we employ the method of Theorem 3.1 Mao et al. [33] and Zuo et al. [20]. The key step is to construct a nonnegative \(C^{2}\)-function \(V: R_{+}^{6}\rightarrow R_{+}\) such that
and
where \(U_{k}=(\frac{1}{k},k)\times (\frac{1}{k},k)\times (\frac{1}{k},k) \times (\frac{1}{k},k)\times (\frac{1}{k},k)\times (\frac{1}{k},k)\) and M is a positive constant.
Define
where \(p>1\), \(R_{1}\), \(R_{2}\), \(R_{3}\) are positive constants defined in Theorem 4.1.
Obviously,
since
On the other hand, by the proof of Theorem 4.1, we have
By Itô’s formula, we have
Therefore,
where \(\tilde{M}=\max_{N_{i},y_{i}\in (0,\infty )} \{- \frac{1}{2} (N_{1}^{p+1}(t) +N_{2}^{p+1}(t)+N_{3}^{p+1}(t)+y_{1}^{p+1}(t)+y_{2}^{p+1}(t)+y_{3}^{p+1}(t) ) +R_{1}N_{1}(t)^{p} (r_{1}+\frac{p-1}{2}\xi _{1}^{2} ) +R_{2}N_{2}(t)^{p} (r_{2}+\frac{p-1}{2}\xi _{2}^{2} ) +R_{3}N_{3}(t)^{p} (-r_{3}+ \frac{p-1}{2}\xi _{1}^{2} )+\sum_{i=1}^{3}a_{ii}N_{i} +a_{12}y_{2}+a_{13}y_{3}+a_{21}y_{1}+ a_{23}y_{3} \}\). Let \(M=r_{3}+\sum_{i=1}^{3}\sigma _{i}+\sum_{i=1}^{3} \frac{\xi _{i}^{2}}{2}+ \tilde{M}\), then \(L V(N_{1},N_{2},N_{3},y_{1},y_{2},y_{3})\leq M\). The proof is completed.
1.2 1.2 Proof of Lemma 2.3
The methods applied here are motivated by [20]. Let
as defined in Theorem 4.1, where
Then, by the proof of Theorem 4.1, we have
For any \(k>0\), we compute
Choosing k sufficiently small such that
we have
where
Applying the same method of Lemma 5.1 in [17] and integrating both sides of \(L(e^{kt}\breve{V}(t))\) and taking expectation lead to
where H is a constant. By the monotonicity of the expectation, we can derive that
By the Cauchy–Schwarz inequality, there exists \(\varrho (p)\) such that
Denote \(K(p)=\max \{\frac{pH}{R_{i}},\varrho (p)(2\sigma _{i}H)^{ \frac{p}{p+1}},i=1,2,3\}\), then
Next computing the derivative of \(\breve{V}(t)\) reads
For small \(\tau >0\) enough and \(n=1,2,\ldots \) , we integrate both sides of \(d\breve{V}(t)\) from nτ to t and take expectation, then
Again using a similar proof of Lemma 5.1 in [17], for any positive constant ϵ and any finitely many n, one can derive that
Letting \(\epsilon \rightarrow 0\) leads to
which implies
Fixing \(\epsilon _{0}=\frac{p}{2(p+1)}\), then there exists \(T>0\) such that \(\ln y_{i}(t)\leq (\frac{1}{p+1}+\epsilon _{0})\ln t\) for all \(t>T\). Therefore,
Together with the positivity of \(y_{i}(t)\), we have
Similarly, we can derive that
that is, \(\limsup_{t\rightarrow \infty }\frac{\ln N_{i}(t)}{\ln t} \leq \frac{1}{p}\), a.s. By the same deduction, we have
The proof is confirmed.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhao, J., Shao, Y. Dynamical analysis of a stochastic three-species predator–prey system with distributed delays. Adv Differ Equ 2021, 344 (2021). https://doi.org/10.1186/s13662-021-03493-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-021-03493-8