
Abstract
Obesity is a prevalent chronic disease that has significant negative impacts on humans and our companion animals, including dogs and cats. Obesity occurs with multiple comorbidities, such as diabetes, hypertension, heart disease and osteoarthritis in dogs and cats. A direct link between lipid metabolism dysregulation and obesity-associated diseases has been implicated. However, the understanding of such pathophysiology in companion animals is limited. This review aims to address the role of lipid metabolism in various metabolic disorders associated with obesity, emphasizing the involvement of the gut microbiota. Furthermore, we also discuss the management of obesity, including approaches like nutritional interventions, thus providing novel insights into obesity prevention and treatment for canines and felines.
Similar content being viewed by others

Introduction
Having cats and dogs as companions is believed to contribute to good physical health and psychosocial well-being of humans; hence, the importance of pets in our lives is becoming obvious [1]. Meanwhile, the prevalence of obesity and associated metabolic diseases is increasing in companion animals. This is closely related to the dysregulation of lipid metabolism, including the biosynthesis and degradation of lipids such as fatty acids (FAs), triglycerides, cholesterol, etc. [2]. While emerging evidence suggests that gut microbiota plays a major role in regulating the host’s lipid metabolism, by producing short-chain fatty acids (SCFAs), which serve as both substrates and modulators. On the one hand, SCFAs especially acetate and butyrate can be converted into acetyl-CoA, thereafter enter the tricarboxylic acid cycle and/or synthesize palmitic acid and other lipids. On the other hand, it is shown in mice and humans that butyrate could promote FA oxidation and stimulate the metabolic activity of brown adipose tissue, subsequently preventing diet-induced obesity and hypertriglyceridemia [3]; gut microbiota can also reshape the host-derived bile acids (BAs) pool by chemically modify them into secondary BAs; as well as regulating host satiety, affecting gene expression, maintaining intestinal barrier function, modulating immune responses, and producing specific enzymes etc. These various processes collectively impact the host's energy expenditure, lipid absorption and metabolic pathways, potentially contributing to the development of metabolic diseases such as obesity [4, 5]. The causal role of gut microbiota in host lipid metabolism dysregulation is best demonstrated in germ-free mice where they develop obesity when colonized with “obese microbiota” [6]. Moreover, recent studies have revealed that the gut microbiome may regulate intestinal epithelial lipid absorption and white adipose tissue lipid accumulation through an array of non-coding RNAs [7, 8]. However, how these lipid metabolism and gut microbiota interact in companion animals is still poorly understood.
In this review, we provide an overview of the occurrence, development, and outcomes of obesity in dogs and cats. Specifically, we aim to elucidate the mechanisms of lipid dysregulation, its connection with obesity-related complications in dogs and cats, and the differences between the two species. Furthermore, we illustrate the potential interaction of lipid metabolism and gut microbiota. In addition, we also discuss obesity management in dogs and cats from a nutritional perspective, opening up new avenues for obesity prevention and intervention in dogs and cats.
Epidemiological and etiological studies of obesity in dogs and cats
Prevalence of obesity in dogs and cats
Similar to humans, obesity is also widespread in domestic cats and dogs. Early studies conducted in the U.S. and other countries have demonstrated that the prevalence of obesity among dogs is between 25% and 40.5%, whereas among cats, it ranges from 6% to 52% [9,10,11,12,13]. This incidence has increased sharply in recent decades [14,15,16,17]. Moreover, the prevalence of comorbidities related to obesity has risen steadily over the past few years. This trend has even surpassed that observed in humans, posing a serious concern within the veterinary community [5, 13, 18]. Therefore, understanding the diagnostic criteria and causative factors of obesity in dogs and cats is crucial for effective management.
Obesity assessment criteria for dogs and cats
In the clinical context, dogs and cats are classified as being overweight or obese when their body weight surpasses the ideal body weight for their body size by 15% or 30%, respectively [19]. However, due to body type, age, and breed variations, this method is highly error-prone when assessing whether pets are obese. Currently, the most accurate method for determining obesity in dogs and cats is measuring their fat and muscle mass using dual-energy X-ray absorptiometry (DEXA) [20]. However, DEXA requires sedation or anesthesia, making it uneconomical and impractical for daily assessment of obesity in dogs and cats. The most widely used method for diagnosing obesity in dogs and cats is the 9-point body condition score (BCS), developed and validated by researchers at Purdue University in 1997 [21]. Compared to body mass index (BMI), which is used for assessing human weight, BCS is used to quantify excess body mass and can give an estimation of excess body fat without measuring weight and height. The ideal BCS is 5, while a BCS of 6 or 7 correlates to overweight and obesity may be identified based on a BCS of 8 out of 9 [22]. Although the evaluation criteria are subjective and depend on factors such as visual and tactile perception, the accuracy of the scoring system is not compromised. Some studies have shown that BCS strongly correlates with DEXA measurement of body fat content [23, 24]. Besides, this scoring system has been incorporated into determining the nutritional requirements of dogs and cats by the National Research Council of the National Academies for calculating their energy needs [25]. Therefore, BCS offers an accurate and convenient approach to assess obesity in cats and dogs.
Major contributing factors to obesity in dogs and cats
To effectively treat obesity in dogs and cats, it is necessary to establish criteria to determine the condition and understand the underlying factors contributing to its onset. Various factors can be classified broadly into two categories: extrinsic/environmental factors and intrinsic factors. For dogs, the primary risk factors for obesity include breed predisposition, age, gender, neutering, and ad libitum feeding [26,27,28]. For cats, in addition to these factors, physical inactivity is also one of the causes of feline obesity [15, 16, 29, 30]. Nowadays, not only diet but also its quality is considered relevant in the development of obesity, i.e., the overall nutritional composition and balance of a diet, including the presence and proportions of essential nutrients. Although it is hard to measure, the concept of diet quality has garnered substantial attention in nutritional research and is closely related to the quality of life. Specifically, a poor-quality diet characterized by excessive caloric intake, imbalanced nutrient composition, and low nutrient density can cause weight gain and obesity in pets [31,32,33,34]. Finally, neutering is of importance for pet obesity since the procedure lowers the metabolic rate of dogs and cats; an increase in food intake will lead to greater susceptibility to obesity in these animals, particularly those who lack exercise [28, 35, 36]. Although pet owners have the means to effectively address these two factors (exercise and ad libitum feeding), some owners may consider a plump appearance endearing, failing to recognize the seriousness of pet obesity as a significant concern [37, 38]. Thus, it is necessary for us to clarify the issue so pet owners can comprehend the detrimental outcomes that obesity can have on their pets.
Outcomes of obesity in dogs and cats
Obesity is a major chronic disease in humans with multiple comorbidities, such as diabetes, hypertension, and coronary heart disease [39]. Similarly, it significantly impacts the health of dogs and cats, increasing their mortality (Fig. 1).

Obesity complications in obese dogs and cats. Overweight dogs are prone to a higher incidence of heart and lung diseases, hypertension, reproductive disorders, and surgical complications. Similarly, obesity in cats is associated with an increased incidence of endocrine and metabolic disorders such as diabetes, hepatic lipidosis, and lameness
Overweight and obese dogs are also prone to have a higher incidence of heart and lung diseases, hypertension, reproductive disorders, and surgical complications [27]. Obesity in cats is associated with an increased incidence of endocrine and metabolic disorders such as diabetes, hepatic lipidosis, and lameness [40].
Diabetes mellitus
Diabetes mellitus (DM) in canine and feline is one of the metabolic diseases caused by genetic predisposition and comes along with obesity, where the animals exhibit insufficient insulin secretion and resistance [41]. Animals with DM could suffer from polydipsia, polyphagia, and polyuria, and start to show weight loss, physical decline, and proteinuria. The accumulation of excess fat and lipid metabolism disorders from obesity leads to insufficient insulin secretion or decreased insulin sensitivity, resulting in increased blood glucose levels in animals and ultimately leading to the occurrence and development of DM [41]. DM in dogs and cats shares several resemblances with human DM. However, it is essential to highlight that there are differences between these species. The effects of insulin resistance in dogs and cats are different. In obese dogs, the impact of a large amount of fat is more insulin resistance, resulting in increased insulin secretion in the first stage. In obese cats, the first stage of insulin secretion will be missing or reduced due to poor glucose tolerance caused by a large amount of fat [42, 43]. However, the final result is type 2 diabetes. Moreover, while obese dogs may display insulin resistance, this does not necessarily lead to the onset of DM [44]. It is worth noting that diabetes in dogs typically presents as a type 1 diabetes-like syndrome. The etiology underlying β-cell destruction in diabetic canines is often elusive, albeit evidence suggests that immunity-mediated destruction of β-cells akin to type 1 diabetes in humans frequently contributes to the explication of pathogenesis [45, 46]. In contrast, overweight cats often develop diabetes mellitus, which shares similarities with human type 2 diabetes [47]. Concurrent with the insulin resistance in DM, dysfunctions in amylin secretion and particularly in the processing of amylin that results in its deposition as amyloid in the islets could potentially account for the gradual demise of β-cells [48]. As a naturally occurring amyloidogenic protein in several animal species, feline islet amyloid polypeptide plays a critical role in β-cell apoptosis [26]. Unlike obese humans and cats, insulin resistance in obese dogs does not spontaneously progress to type 2 DM, because the amylin protein in dogs does not aggregate and form pancreatic islet amyloid [49, 50]. Together, obesity-induced DM is one of the critical metabolic diseases in dogs and cats.
Hepatic lipidosis
Obesity not only leads to diabetes but also increases the risk of hepatic lipidosis. Hepatic lipidosis is identified by the atypical buildup of triglycerides in over 80% of hepatocytes [51, 52]. This accumulation leads to a secondary impairment of liver function and intrahepatic cholestasis [53, 54]. Low-density lipoprotein (LDL) content is significantly higher in cats with hepatic lipidosis than in healthy cats [55, 56].
Hepatic lipidosis is the most common hepatobiliary disease and has been more widely studied in cats than dogs [54, 57, 58]. Meanwhile, in dogs, researchers generally use the term “diffuse vacuolar hepatopathy” to refer to this type of disease rather than "hepatic lipidosis" [59]. Hence, the present section only focuses on feline hepatic lipidosis. Some studies have observed that the progression of hepatic lipidosis is most prompt and consistent when felines exhibit elevated BCS [57]. Feline hepatic lipidosis is more prevalent and often results from an underlying disease, including DM, pancreatitis, or renal failure [60]. The pathophysiology of feline hepatic lipidosis is complex and not yet fully illustrated. Targeting lipid metabolism will help to ameliorate obesity and its related hepatic lipidosis.
Other complications of obesity in cats and dogs
In dogs and cats, aside from the development of diabetes and hepatic lipidosis, obesity is also associated with the incidence of other diseases, impacting cardiac function negatively [61]. Compared with lean canines, corpulent canines exhibit an elevation in heart rate and a mild to moderate augmentation in blood pressure [62, 63]. It is well-established that obese cats tend to display a higher heart rate and greater cardiac muscle contraction compared to their average-sized counterparts [64]. Additionally, obesity leads to the accumulation of large amounts of adipose tissue, infiltrating the atrial tissue [65].
Obese felines and canines have been demonstrated to have an increased vulnerability to urolithiasis, which is correlated with elevated food consumption, adipose tissue accumulation, and a heightened excretion of minerals within the urinary system [66]. Urolithiasis, commonly called urinary stone disease, is characterized by calculi formation within the urinary system [67]. Based on their location within the urinary tract, the calculi are categorized as nephroliths (in the kidneys), ureteroliths (in the ureters), urocystoliths (in the bladder), and urethroliths (in the urethra) [66]. Osteoarthritis is a debilitating condition marked by the gradual deterioration of articular cartilage, resulting in the loss of both proteoglycans and collagen [68]. Additionally, subchondral bone sclerosis, periarticular osteophyte formation, and chronic synovial membrane inflammation further exacerbate the disease's progression [68]. Although various factors such as age, gender, and breed of animals can contribute to the development of osteoarthritis, obesity stands out as a particularly influential risk factor [69]. Obesity increases mechanical stress on the cartilage and leads to a higher incidence of non-weight-bearing joint involvement in osteoarthritis [70]. Obesity poses various risks to dogs and cats, suggesting that its treatment is a tremendous priority in pet health.
Association between obesity and lipid metabolism dysregulation in pets
Comparison of lipid metabolism in dogs and cats versus other animals
De novo lipogenesis
De novo lipogenesis (DNL) or de novo FA synthesis is a metabolic pathway that generates fatty acids from surplus carbohydrates. These fatty acids can be incorporated into triglycerides (TGs) for energy storage [71]. Under normal conditions, DNL occurs primarily in the liver and adipose tissue and is believed to play a major role in maintaining serum TG homeostasis [72]. In canines and felines, DNL predominantly occurs in adipose tissues, whereas in humans and rodents, the liver is the major site [73] (Fig. 2). Acetate is considered to be the primary substrate for DNL in feline [74]. Furthermore, it has previously been shown in organoids derived from the livers of mice, humans, dogs, and cats when administered with FA, lipid accumulation can be detected in all species. Notably, feline liver organoids exhibited a greater propensity for lipid droplet accumulation than their human counterparts [75]. It is possible that the liver of cats has a relatively weak capacity for lipid metabolism. Some researchers speculate that the other metabolic pathways handling excess FFA (β-oxidation, very low-density lipoprotein secretion) are quickly saturated in feline hepatocytes, leading to extensive lipid-droplet formation [75]. Owing to the inefficiency of the feline liver in metabolizing FA originating from abundant peripheral lipolysis, a buildup of lipids transpires in the liver [55]. In addition, dogs, being descendants of wolves, have evolved to adeptly manage the fluctuating nutrient availability associated with feast and famine eating patterns. This adaptation has endowed them with superior resilience in coping with nutritional extremes [76]. Therefore, compared to cats, dogs are generally less susceptible to developing "hepatic lipidosis".

Differences in lipid metabolism underlying diabetes mellitus in obese dogs versus cats. In canine, the lack of CETP enzyme causes a perpetual acquisition of cholesteryl esters by HDL2 molecules, ultimately leading to the generation of distinctive HDL1 molecules. Next, HDL1 facilitates the transfer of cholesteryl esters from tissues to the liver for either disposal or reuse rather than to LDL or VLDL molecules that, in humans, transport cholesterol to peripheral tissues. TG: triglyceride, VLDL: very low-density lipoproteins, LDL: low-density lipoproteins, HDL: high-density lipoproteins, CETP: cholesterol ester transfer protein, FFA: free fatty acid
Lipoprotein metabolism
Obesity is correlated with dysregulation of lipid metabolism. Studies have demonstrated that overweight neutered cats exhibit elevated serum non-esterified fatty acids (NEFAs), triglycerides, and cholesterol compared to their lean counterparts during fasting [77]. These aqueous insoluble lipids are transported through plasma in particular particles called lipoproteins. Chylomicrons, which are large lipoproteins rich in triglycerides, are assembled around apoB48 in enterocytes of the small intestine and transport dietary lipids [78]. In contrast, very low-density lipoproteins (VLDL), which are smaller in size, contain apoB100 and are produced by hepatocytes [79]. High-density lipoproteins (HDLs) are primarily synthesized in the liver and play a crucial role as donors and recipients of apolipoproteins C, apo E, and various lipids from other circulating lipoproteins [80]. HDLs also have a critical function in the reverse cholesterol transport pathway by transferring cholesterol from peripheral tissues to small circulating discoid HDL molecules, which leads to the conversion of these molecules into nascent HDL3 molecules [81, 82]. In canine, the lack of cholesterol ester transfer protein (CETP) enzyme, which is present in humans, causes a perpetual acquisition of cholesteryl esters by HDL2 molecules, ultimately leading to the generation of distinctive HDL1 molecules [83] (Fig. 2). Next, HDL1 facilitates the transfer of cholesteryl esters from tissues to the liver for either disposal or reuse rather than to LDL or VLDL molecules that, in humans, transport cholesterol to peripheral tissues [59]. It has been proposed that this role of HDL1 contributes to the lower prevalence of atherosclerotic disorders in dogs relative to humans [84]. Although CETP enzyme deficiency is also present in cats, it does not lead to the formation of HDL1 particles as observed in dogs [80, 85]. It is also observed that atherogenesis and coronary artery disease are not a feature of feline obesity or diabetes [80, 86]. The precise mechanism underlying this phenomenon has not yet been comprehensively investigated.
Adipokines
Obesity in cats and dogs has been associated with alterations in adipokines [87, 88]. Currently, the most studied adipokine in cats and dogs is adiponectin. Fully differentiated adipocytes exclusively synthesize adiponectin and circulate in the plasma as multimers, with the high molecular weight isoform exhibiting the most significant biological activity [26]. In felines, the expression of the adiponectin gene is markedly elevated in visceral adipose depots compared to other depots [89, 90]. Some findings have indicated that high molecular weight (HMW) multimers constitute approximately 80% of total adiponectin in cats, whereas humans exhibit only 30% HMW adiponectin [91, 92]. Similarly, dogs possess a higher concentration of HMW adiponectin than humans [93]. HMW multimers, strongly related to insulin sensitivity and body fat mass, show more excellent activity compared to total adiponectin [92]. In addition, adiponectin has profound effects on increasing insulin sensitivity and decreasing lipid concentrations, as well as anti-inflammatory and anti-atherosclerotic properties [94,95,96]. This could be a potential explanation for the lower susceptibility of dogs and cats to atherosclerosis. In humans, reduced levels of circulating total or HMW adiponectin are associated with insulin resistance and are indicative of the onset of type 2 diabetes or other metabolic syndrome [97]. Some studies have failed to find a correlation between obesity and adiponectin concentrations in dogs despite significant evidence of obesity’s effects on insulin resistance [88, 98, 99]. Unlike in humans, obese dogs do not experience a decrease in HMW adiponectin levels, nor do the levels change after losing weight [93, 99]. It has been suggested that this species’ difference in adiponectin may be a protective factor against developing type 2 diabetes in dogs since this condition does not occur naturally in this species [93].
The gut microbiota associated with obesity and its regulatory mechanism
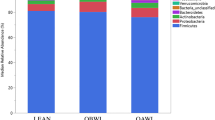
Recent studies have consistently established a strong association between gut microbiota dysbiosis and the onset and progression of obesity [100]. Evidence of significant differences in the composition and biodiversity of gut microbiota between obese and healthy pet animals is also accumulating (Fig. 3). In dogs and cats, five major phyla have been identified: Firmicutes, Fusobacteria, Bacteroidetes, Proteobacteria, and Actinobacteria [5, 13].

Altered gut microbiota in obese dogs and cats
Gut microbiota alteration in obese dogs
A study of 20 dogs reveals that the relative abundance of Firmicutes was lower in obese dogs when compared to the lean ones, including a decrease of Clostridiaceae (affiliated with Firmicutes). Meanwhile, the number of Bacteroidetes was higher in obese dogs, mainly due to the increase of Bacteroidaceae. Interestingly, the relative abundance of both Fusobacteria and Proteobacteria was increased in the gut of obese dogs, while these bacteria were reduced when obese animals lost weight. Additionally, it is also found that the abundance of Coriobacteriaceae, which belongs to the Actinobacteria phylum, was reduced in obese dogs, whereas in obese dogs lost weight, its abundance was then increased to the level in the gut of lean dogs [101]. Another study also shows a significant increase of Proteobacteria and Fusobacteria [102] in the gut microbiota of obese dogs compared to the lean ones. While the latter gut microbiota is predominantly dominated by Firmicutes [103]. Consistently, a study of 21 obese and 22 lean dogs shows a higher relative abundance of Actinobacteria and Roseburia in lean dogs’ gut microbiota compared to obese dogs [104]. In contrast, an analysis of 66 clinically healthy dogs’ fecal samples using 16S rRNA gene sequencing reveals a higher proportion of Bifidobacteriaceae (affiliated with Actinobacteria) in obese dogs. At the genus level, Blautia, Lachnospiraceae, and Eubacterium biforme are increased in the normal weight dogs, while the abundance of Ruminococcus was decreased in obese dogs [105]. Together, these studies suggest significant alterations in the gut microbiota of obese dogs compared to the healthy lean ones. However, due to the heterogeneity of these studies, including variations in factors such as age, sex, breed, nutrition level, and genetic factors, the conclusions drawn are inconsistent.
Gut microbiota alteration in obese cats
Similarly, the gut microbiota of obese cats is also dysbiotic. A metagenomic study of the gut microbiota of 8 obese cats and 8 healthy cats has found that the microbial diversity and the Firmicutes-to-Bacteroidetes ratio in obese cats are significantly reduced, while the abundance of Bifidobacterium sp., Dialister sp., Olsenella provencensis and Campylobacter upsaliensis are increased compared to the healthy control. Additionally, the quantity of Fusobacteria, Erysipelotrichaceae bacterium, and Phascolarctobacterium succinatutens is decreased in the obese cats’ microbiota [106]. Furthermore, a microbial study of 82 domestic cats has also shown an increase in Bifidobacterium abundance and a decrease in Fusobacteria abundance in obese cats, whereas the abundance of Coriobacteriales belonging to the Actinobacteria phylum was decreased compared to the lean ones [107]. Another study on the effects of dietary intervention on obese cats has shown that the abundance of Actinobacteria increases in obese cats during weight loss, primarily due to changes in Bifidobacterium sp. and Collinsella sp. Meanwhile, there is a decrease in bacterial infections in obese cats when they lose weight, mainly driven by the Prevotella spp. reduction. Within the Firmicute phylum, Blautia, Dorea, Eubacterium, Oscillospira, Peptococcus, and Ruminococcus increased whereas Lactobacillus, Butyricicoccus, and Phascolarctobacterium decreased overtime [108]. A 16S rRNA gene sequencing study on the feces of 24 cats (including eight obese cats) shows a significant decrease in the Firmicutes-to-Bacteroidetes ratio in obese cats. Interestingly, there are substantial increases in Prevotella while the proportions of Blautia and Clostridium decline [109]. A study involving 14 lean neutered cats and 17 obese neutered cats has found that the gut microbiota of obese cats is enriched with Firmicutes, Actinobacteria, Proteobacteria, and Planococcaceae incertae sedis. However, this study found no significant differences in the biodiversity index of microbiota between obese and lean cats [110]. Overall, in the gut microbiota of obese cats, there is an increase in the abundance of Proteobacteria and a decrease in the abundance of Actinobacteria. Additionally, unlike obese dogs, there is a decrease in the Firmicutes-to-Bacteroidetes ratio and a decline in the abundance of Fusobacteria in the gut microbiota of obese cats. The difference may be related to the variations in the intestinal structure and dietary habits of the two animals.
Taken together, gut microbiota plays vital roles in pet lipid metabolism. Bacteroidetes, Firmicutes, and Actinobacteria produce SCFAs by fermenting non-digestible carbohydrates (such as starch, inulin, cellulose, hemicellulose, pectin, and gum). These SCFAs provide energy to the intestinal epithelium and other tissues and support the intestinal defense mechanism by lowering the intestinal pH value. Furthermore, they influence the capacity for intestinal absorption and digestion, ultimately affecting obesity [111]. However, there are currently limited studies on the mechanism of intestinal microbiota regulating animal fat deposition. Metagenomics, metabolomics, and transcriptomics offer potential explanations for the interaction between these two factors. By utilizing these multi-omics research methods, we can further explore the mechanisms of intestinal microorganisms in pet lipid metabolism. Thus, we may identify potential signaling pathways and regulatory factors and discover new therapeutic and intervention strategies for controlling and regulating animal fat deposition.
The mechanisms of gut microbiota in regulating lipid metabolism and the potential role of the microbiota-gut-brain axis
The gut microbiota plays a crucial role in regulating the host’s lipid metabolism through various mechanisms such as the production of SCFAs, BA metabolism, hunger regulation, maintenance of intestinal barrier function, immune response modulation, and specific enzyme production. These processes impact energy expenditure and lipid absorption, potentially affecting the onset of metabolic diseases like obesity. Moreover, the dysregulation of the gut microbiota can impair the host's ability to process polysaccharides and absorb dietary lipids, leading to disruptions of liver and adipose tissue functions and initiating a cascade of metabolic disorders, including obesity, fatty liver, type 2 diabetes, and cardiovascular diseases [6]. However, the precise mechanisms underlying the interplay between gut microbiota and lipid metabolism require further investigation. Existing evidence has demonstrated that the gut microbiome can regulate lipid metabolism and subsequently impact body weight via the modulation of specific microbial metabolites, such as BAs, SCFAs, TMAO, tryptophan and indole derivatives [112,113,114]. For instance, TMAO can induce a high liver score of fatty liver hemorrhage syndrome by reducing the total BA content in the liver and increasing the total cholesterol [113]. These microbial metabolites engage in a cross-talk with the intestinal barrier and endocrine hormones, consequently regulating lipid metabolism through the microbiota-gut-brain axis.
Recently, it has also been suggested that the gut microbiota communicates directly with the brain through neurological pathways, such as the vagus nerve, and affects brain function via hormonal and immune pathways. Microbiota-derived SCFAs can regulate the host’s energy balance and mood through specific receptors, impacting the synthesis of key neurotransmitters like serotonin [115]. Moreover, the gut microbiota potentially modulates the expression of brain-derived neurotrophic factor (BDNF), which is crucial for neuronal growth and survival. In return, the brain influences the composition and function of the gut microbiota through neuroendocrine stress responses and regulates gastrointestinal motility and secretion via the autonomic nervous system [115,116,117]. The brain also modulates the gut microbiota by affecting the activity of immune cells and immune responses, while controlled behaviors and dietary habits directly impact these microbes [118]. These physiological processes described are all normal physiological processes for animals within the normal range. However, when they are imbalanced or excessive, they can all potentially induce metabolic and obesity issues in animals, which is why gut microbiota is so crucial for the development of obesity in animals (Fig. 4).

Why the gut microbiota plays a crucial role in the development of obesity. Gut flora in obese dogs and cats ① can affect lipid metabolism in several ways: ② LPS is increased in the intestines of obese dogs and cats and causes inflammation and fatty liver when it enters the bloodstream through a weakened intestinal barrier. ③ TMA produced by intestinal flora through the metabolism of methylamine-containing nutrients is converted to make TMAO, which promotes hepatic fat formation by affecting bile acids. ④ The gut flora of obese dogs and cats produces more SCFAS, such as acetate and butyrate, that promote de novo fat synthesis. ⑤ Tryptophan is metabolized by the intestinal microbiota to indole and its derivatives that can stimulate the production of GLP-1 by enteroendocrine L-cells and insulin secretion by pancreatic β-cells and serve as molecular signals to regulate food intake and appetite. ⑥ Gut flora alters GLP-1, GLP-2, and PYY secretion by regulating enteroendocrine cells (L cells). ⑦ Gut flora communicates with the brain through metabolic byproducts and neural pathways, influencing mood and behavior. ⑧ Brain modulates the composition and function of the gut flora through neuroendocrine responses and immune system alterations. LPS: lipopolysaccharide, GLP-1: glucagon-like peptide-1, GLP-2: glucagon-like peptide-2, PYY: peptide tyrosine tyrosine, Trp: tryptophan, I3CA: indole-3-carboxylic acid
The significance of diverse nutritional requirements in the management of obesity in dogs and cats
Variations in nutrient requirements between dogs and cats
In the wild, cats consume prey that is rich in proteins and contains moderate amounts of fat and minimal quantities of carbohydrates [119]. Consequently, they have evolved as a carnivore species, possessing a metabolic adaptation that allows for a higher rate of protein metabolism and lower utilization of carbohydrates compared to dogs and other omnivores [120]. In contrast, in canines, approximately 40% to 70% of ingested glucose during a meal is absorbed by the liver via the portal circulation [121]. Whereas the feline hepatic glucose extraction is comparatively low due to the absence of the hepatic enzyme glucokinase [122]. The lack of this enzyme reflects, to a certain extent, the relatively limited contribution of dietary carbohydrates in maintaining blood glucose levels in feline physiology. The evolutionary divergence in energy metabolism determines that feline species refrain from down-regulating amino acid catabolic enzymes, even when dietary protein is limited. It implies that cats may have to engage in the breakdown of lean mass to generate amino acids for hepatic gluconeogenesis, thereby supporting the maintenance of circulating glucose levels [123]. The significant differences in protein requirements between cats and omnivorous species, such as dogs, further emphasize this important metabolic distinction. For instance, while the protein requirement for kittens is 1.5 times higher than that of young individuals from other species, adult cats require 2 to 3 times more protein in their diet than mature individuals from omnivorous taxa [124].
Nutritional and health risks of restricted feeding
By implementing a restricted diet, it is possible to maintain a healthy weight for both dogs and cats, preventing obesity. However, it should be noted that limited food intake in cats can potentially lead to lipid deposition disorder [57]. Energy restriction may prompt the mobilization of FA. Still, insufficient choline intake resulting from the limitation can compromise FA transport from the liver and reduce FA oxidation, leading to fat accumulation and eventually hepatic lipid deposition [57]. In clinical practice, it is generally considered safe for cats to lose weight at a rate of 0.5%–2% of their initial body weight per week [25]. Rapid weight loss beyond this recommended range can increase the risk of feline hepatic lipidosis [125]. However, currently there is no standardized formula in the pet industry for calculating how to restrict energy intake [25, 126,127,128]. Therefore, the sector must unify and establish a scientifically-based program for energy restriction in the future.
Nutritional interventions targeting gut microbiome
Dietary intervention
Variations in macronutrient ratios have been reported to significantly impact the gut microbiota composition of animals, including cats [129]. In dogs, the composition and function of the gut microbiota can be selectively altered by diets with varying macronutrient ratios, especially during obesity [130, 131]. Weight-loss foods are often characterized to have a high protein, low carbohydrate, and low-fat content. Numerous studies conducted on humans, mice, dogs, and cats have demonstrated the potential benefits of these diets, including increased satiety, decreased body fat, and maintenance of lean body weight [132,133,134,135]. Notably, the reason for the elevated protein inclusion in the weight loss formulations is not solely for the satiating effect but rather to ensure animals continue to receive adequate protein intake while the total energy consumption is reduced, thus maintaining a high protein-to-energy ratio (PER) [136, 137]. Since pets depend on their owners for food, maintaining a high PER is critical, especially during weight management programs. This approach allows pets to consume significantly less energy while obtaining sufficient protein. In addition, the provision of protein can also facilitate fat reduction and aid in preventing weight rebound [130]. Despite the potential benefits, excess protein intake has been associated with adverse effects on the gut microbiota, altering the metabolic byproducts such as causing SCFAs reduction or inducing TMAO production [138,139,140] and promoting intestinal inflammation [141]. A study of dogs found that in contrast to increased Bacteroides uniformis and Clostridium butyricum promoted by a low-protein, high-carbohydrate diet, the growth of a high-protein, low-carbohydrate diet increased the abundance of Clostridium hiranonis, Clostridium perfringens, and Ruminococcus gnavus, all of which are potential pathogens. Additionally, the high-protein, low-carbohydrate diet resulted in a decrease in the Bacteroidetes-to-Firmicutes ratio and an increase in the Bacteroides-to-Prevotella ratio [130].
High-carbohydrate diets may contribute to obesity. Excessive consumption of carbohydrates and exceptionally high simple sugar intake can rapidly increase blood glucose levels, stimulating insulin secretion and promoting fat production. Furthermore, a high-sugar diet can induce insulin resistance, resulting in hyperinsulinemia and high blood pressure [142, 143]. However, current research cannot establish a causal relationship between a high-carbohydrate diet and pet obesity. A study involving neutered cats reveals that high-fat diets resulted in more significant weight gain than a high-carbohydrate diet [144]. This implies that providing low-energy density feed may offer a viable strategy for preventing obesity.
Indeed, fat is one of the three macronutrients, characterized by its high energy density and compact size. A high-fat diet can modify the composition of the gut microbiota and impair the intestinal mucosa, contributing to the development of obesity [145]. A study of healthy adult dogs shows that high-fat, low-carbohydrate diets with different percentages of canola oil inclusion resulted in linear increases of Tenericutes, Spirochaetes, Fibrobacteres, and Planctomycetes according to fat levels. In contrast, Prevotellaceae, Catenibacterium, Alloprevotella, and bacteria belong to the Lachnospiraceae family, and the genus Bacteroides showed a linear decrease in abundance with increasing fat content in the diet. It is speculated that bacterial populations that increase dietary fat levels may utilize fat or bile as a key metabolic substrate and vice versa [146]. In addition, studies have shown that specific gut microbes, such as Bifidobacterium, can directly or indirectly regulate the absorption, bioavailability, and biotransformation of omega-3 polyunsaturated fatty acids (PUFAs), further affecting FA intake and their functions. PUFA-derived metabolites produced by the gut microbiota may be a new type of active metabolite, capable of exerting anti-obesity and anti-inflammatory effects through various mechanisms [147]. Another study has also shown that the gut microbiota can convert dietary PUFAs, such as 10-hydroxy-cis-12-octadecenoic acid (HYA) derived from linoleic acid, into a variety of metabolites. Furthermore, supplementation with HYA can alleviate obesity induced by a high-fat diet in mice. This effect is independent of arachidonic acid (AA)-mediated adipose inflammation and improves metabolic conditions through free fatty acid receptors. By producing metabolites such as HYA, the gut microbiota helps to regulate the host's energy balance and insulin sensitivity, thereby enhancing resistance to metabolic disorders [148].
Probiotics
In addition to macronutrient inclusion, supplementation of probiotics, prebiotics, synbiotics and/or postbiotics may also be approaches of interest in managing obesity in pets (Table 1). Probiotics are defined as "living microorganisms that, when ingested in adequate amounts, confer health benefits to the host" [149]. Recent studies have demonstrated the significant role of probiotics in improving gut health. In a study of mice fed a high-fat diet, Lactobacillus plantarum K50 administration reduced fat content in adipose tissue and liver and improved gut microbial composition [150]. Furthermore, studies have demonstrated the ability of probiotic supplementation, particularly with Lactobacillus and Bifidobacterium, to effectively lower weight gain induced by a high-fat diet in obese mice and their blood glucose and lipid levels [151]. Moreover, Lactobacillus fermentum CECT5716 has ameliorated high-fat diet-induced obesity in mice by restoring the gut microbiota to its original normal state [152]. Studies suggest that probiotics exert a regulatory effect on gut microbiota, potentially through immune interactions, achieving weight loss goals in obese patients. Therefore, using probiotics may also improve the body fitness of pets [149].
Indeed, a study of high-fat-induced obesity in canines has demonstrated that Enterococcus faecium IDCC 2102 and Bifidobacterium lactis IDCC 4301 effectively promoted the proliferation of beneficial commensal bacteria in the intestinal tract. These probiotics activated glycolysis and pyruvate metabolism, enhancing energy metabolism and curbing weight gain in obese animals [154]. Marcináková et al. [155] also reported that oral administration of probiotic Enterococcus faecium strain EE38 decreased the total lipids and protein levels in blood from 8 out of 11 healthy dogs, where cholesterol levels reached physiological levels. Enterococcus faecium SF68 is an EU-approved pet probiotic commonly used in research on pet diseases [160, 161]. However, a study conducted by Kathrani et al. [153] on naturally occurring obesity in cats, free from specific pathogens, shows that this probiotic did not influence animal body weight, composition, or metabolic parameters. Notably, this study's intervention duration was short. Probiotic products for dogs and cats are trendy nowadays, where optimal effects are often found with host-derived probiotic preparations [162]. Indeed, it is suggested that probiotics of host origin may exhibit higher colonization efficacy in the intestines of the corresponding species [163]. Thus, using beneficial microorganisms isolated from dogs and cats may better address the gastrointestinal needs of obese animals.
Prebiotics/synbiotic
Prebiotics are substances that can be selectively utilized by intestinal microorganisms and benefit the host’s health. They encompass functional oligosaccharides, non-starch polysaccharides, resistant starch, proteins, plant extracts etc. [164]. A study involving nine overweight beagles reveals that the commercial prebiotic inulin-type fructans increased the SCFA production and the relative abundance of Eubacterium (affiliated with Firmicutes), and decreasing the relative abundance of some Proteobacteria, ultimately improving the glucose and insulin incremental area under the curve of the overweight animals [156]. There have been reports on the positive effects of short-chain fructo-oligosaccharides on reducing fat deposits and regulating specific bacteria in animals [165]. A study on high-fat feeding in mice found that galactooligosaccharides addition restored the intestinal microbial dysbiosis by increasing the reduction of beneficial microbes such as Akkermansia muciniphila, Bacteroides acidifies, Lactobacillus gasseri and Bifidobacterium pseudopodium, which was closely related to the improved body weight gain, insulin resistance, and lipid metabolism [166]. Research has also found that bamboo shoot fiber effectively prevents obesity in mice by modulating the intestinal microbial community [167]. Ingesting Ganoderma lucidum water extract can reduce weight and inflammation and alleviate insulin resistance in mice on a high-fat diet [156]. These studies imply that prebiotics may also be used in managing pet obesity. However, most prebiotic studies were conducted with healthy cats and dogs [14]. One study of 12 healthy adult domestic shorthair cats found that resistant starch promoted the intestinal growth of Actinobacteria and butyrate production while reducing the blood cholesterol levels in cats [168]. Similarly in canines, prebiotic fibers are also reported to enrich fiber-fermenting microbes and increase SCFA production [14, 169].
Synbiotics are mixtures comprising live microorganisms and substrate(s) selectively utilized by intestinal microorganisms that confer health benefits on the host [170]. It is summarized that certain combinations of probiotics and prebiotics exhibit superior beneficial effects compared to products containing either probiotics or prebiotics alone in managing obesity and the associated gut microbiota dysbiosis [171,172,173,174]. In obese dogs, it is found that a synbiotic containing Lactobacillus gasseri BNR17 and a non-digestible maltodextrin, galacto-oligosaccharides, fructo-oligosaccharides, and polydextrose mixture can decrease body weight and subcutaneous fat levels at the third lumbar vertebra, enhance microbial diversity, and improve carbohydrate and lipid metabolism [157]. There is a lack of primary experimental data on probiotics and prebiotics compatibility, which warrants further investigations.
Postbiotics
Postbiotics are defined as non-living microorganisms and their components that could confer health benefits to the host [175]. Effective postbiotics must contain inactive microbial cells or cell components, with or without metabolites, contributing to observed health benefits. A study of 48 healthy dogs reveals that feeding fiber and polyphenolic components resulted in a shift of gut microbiota in the colon from primarily digesting proteins to primarily digesting carbohydrates, thus producing potential postbiotic compounds [158]. Similarly, another study on postbiotics in cats also indirectly confirmed the potential of certain positive postbiotics to alleviate obesity in pets [176]. Conversely, dietary fats significantly affect the gut microbiota metabolism and potential postbiotic production. Specific FA, such as medium-chain triglycerides and long-chain polyunsaturated FA, can regulate the metabolic activities of the microbial community, leading to changes in the levels of various lipid classes. Furthermore, the intake of specific long-chain polyunsaturated FA can alter the concentrations of gut microbiome-derived postbiotics, such as indoles/indolic sulfates and phenols/phenolic sulfates, thereby further influencing health, obesity and related metabolic diseases [176, 177].
Fecal microbiota transplantation (FMT)
Fecal microbiota transplantation has emerged as a technique in medicine in recent years and has undergone clinical applications in treating gut microbiota-related diseases in humans and dogs [178, 179]. This therapy involves transferring a donor's healthy microbial community using a fecal preparation into the recipient's gut, thereby reconstructing their gut microbiota composition and achieving therapeutic goals. Symbiotic bacteria are considered the primary practical components of FMT. However, other elements such as viruses, bacterial fragments, proteins, antimicrobial compounds, metabolites, oligonucleotides, and shed cells from the donor also play essential roles [180]. Previous research has investigated the indications of FMT in dogs, showing positive regulatory effects on disease treatment. For instance, dogs treated with FMT recovered quicker from acute diarrhea than those treated with metronidazole. The FMT-treated dogs exhibited a lower abundance of E. coli and Streptococcus spp. in their gut microbiota and an overall more diverse community [178]. FMT has also been demonstrated to be effective in treating diseases such as Clostridium difficile infection (CDI) [181], inflammatory bowel disease (IBD) [182,183,184], canine parvovirus (CPV) enteritis [185], and acute hemorrhagic diarrhea syndrome (AHDS) [186, 187]. However, research using FMT in cats is relatively limited compared to dogs. Furmanski et al. [188] reported a case study of a cat with ulcerative gastroenteritis unresponsive to conventional treatments. After undergoing two FMT procedures, the cat’s condition was gradually stabilized, and its bowel movements were normalized [188]. Another case study found FMT improved vomiting and diarrhea in a 6-year-old cat [189]. Furthermore, a study evaluating the changes in fecal microbiota after FMT in cats confirmed the efficacy of orally administered FMT capsules in improving chronic colitis [190].
Unfortunately, there is a lack of research exploring the effects of FMT on obesity in pets. Nevertheless, considering the findings from human FMT studies, it is reasonable to speculate that FMT may play a positive role in managing pet obesity. A study in mice reveals that FMT had mitigated obesity induced by a high-fat diet [191]. The effects may be attributed to the significant alteration of the composition and structure of the gut microbiota following FMT, including an increased abundance of Romboutsia and Lactobacillus. Interestingly, changes in the composition of the gut microbiota would not only affect the process of intestinal lipid metabolism but also impact the levels of m6A, a methylation modification found on RNA molecules that plays a crucial role in regulating gene expression. Variations in the gut microbiota, by modulating m6A levels, may alter the absorption and excretion of fats, thereby reducing the intestinal uptake of lipids, enhancing lipid excretion, and ultimately boosting the host’s resistance to obesity [192, 193]. In a study by Ridaura et al. [194], they cohabitated mice carrying the gut microbiota from obese twins with mice carrying the gut microbiota from lean twins. The researchers observed that mice with ‘obese microbiota’ exhibited inhibition of fat growth, body weight, and the development of obesity-related metabolic phenotypes. Additionally, the microbiota metabolic profile of the obese mice was eventually transformed into a state similar to that of lean mice. These promising studies suggest that FMT may have become a valuable approach for testing pet obesity.
Although FMT therapy has emerged as a potential alternative to conventional drug treatments in veterinary clinics, it has also encountered several challenges. One of the critical issues is the stringent screening process required for donor feces to prevent infectious disease transmission. Moreover, a notable obstacle is the absence of a dedicated “fecal bank” for pets, which adds complexity and expense to the screening process. While trying to establish such banks, they would face some difficulties. Firstly, the infrequent use of FMT in the current veterinary clinics could result in the long-term storage of fecal material, potentially compromising its efficacy. Additionally, research on pet FMT is limited, failing to account for the potential impacts of breed, size, age, and physiological status on its effectiveness. Further research on pet FMT is essential to understand its mechanisms and applications [178] comprehensively.
In general, significant gaps remain in understanding the mechanisms of gut microbiota in controlling obesity and identifying potential targets for obesity prevention. Various factors influence obesity and the complex interactions among gut microbiomes and host metabolism. Interventions like fecal microbiota transplantation, probiotics, and prebiotics could quickly address gut microbiota imbalance. However, a drawback is that once the intervention ceases and the patient resumes a normal diet, the gradual disappearance of the acquired beneficial microbiota may occur. Therefore, the combination of multiple strategies is vital for sustaining a stable gut microbiota and could have a synergistic effect in regulating the body’s metabolism. This entails reducing the consumption of high-energy-density foods, modifying the macronutrient composition (e.g., high-protein, low-fat, and high-fiber diets), and incorporating prebiotics and probiotics [153, 195,196,197]. Nevertheless, current research indicates that approximately half of obese cats and dogs regain weight following a weight loss plan. This emphasizes the need for personalized nutrition plans to effectively control weight and achieve precise nutritional effects.
Conclusion
Over the past decades, a high prevalence of obesity has been found in dogs and cats. It is clear that obesity is associated with several related chronic diseases in these animals. While previous studies have shed light on the role of lipid metabolism in obesity, the process of lipid metabolism in companion animals is still poorly understood. This study clarifies the importance of lipid metabolism and its relationship with gut microbiota. Additionally, this review discusses nutritional interventions for managing obesity in dogs and cats, specifically focusing on the gut microbiome. These interventions involve dietary modifications, probiotics, prebiotics/synbiotics, postbiotics, and FMT. These interventions show potential for enhancing lipid metabolism in obese dogs and cats and offer valuable scientific evidence and practical nutritional strategies for clinical practice.
Availability of data and materials
Not applicable.
Abbreviations
- AHDS:
-
Acute hemorrhagic diarrhea syndrome
- BA:
-
Bile acids
- BCS:
-
Body condition score
- BDNF:
-
Brain-derived neurotrophic factor
- CETP:
-
Cholesterol ester transfer protein
- CHO:
-
Cholesterol
- DEXA:
-
Dual-energy X-ray absorptiometry
- DM:
-
Diabetes mellitus
- DNL:
-
De novo lipogenesis
- FA:
-
Fatty acid
- FFA:
-
Free fatty acid
- FMT:
-
Fecal microbiota transplantation
- HDL:
-
High-density lipoproteins
- HMW:
-
High molecular weight
- HYA:
-
10-Hydroxy-cis-12-octadecenoic acid
- LDL:
-
Low-density lipoproteins
- NEFA:
-
Non-esterified fatty acid
- PUFA:
-
Polyunsaturated fatty acid
- SCFA:
-
Short-chain fatty acid
- TG:
-
Triglyceride
- TMAO:
-
Trimethylamine oxide
- VLDL:
-
Very low-density lipoproteins
References
Kete K. Pets in America: a history. Am Hist Rev. 2007;112(3):899–900.
Upadhyay J, Farr O, Perakakis N, Ghaly W, Mantzoros C. Obesity as a disease. Med Clin. 2018;102(1):13–33.
Li Z, Yi CX, Katiraei S, Kooijman S, Zhou E, Chung CK. Butyrate reduces appetite and activates brown adipose tissue via the gut-brain neural circuit. Gut. 2018;67(7):1269–79.
Wang Y, Han T, Liu T, Sun L, Dou B, Xin J, et al. New insights into starch, lipid, and protein interactions - Colon microbiota fermentation. Carbohydr Polym. 2024;335:122113.
Pilla R, Suchodolski JS. The role of the canine gut microbiome and metabolome in health and gastrointestinal disease. Front Vet Sci. 2020;6:498.
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444(7122):1027–31.
Wang YH, Wang M, Chen JX, Li Y, Kuang Z, Dende C, et al. The gut microbiota reprograms intestinal lipid metabolism through long noncoding RNA. Science. 2023;381(6660):851–6.
Virtue AT, McCright SJ, Wright JM, Jimenez MT, Mowel WK, Kotzin JJ, et al. The gut microbiota regulates white adipose tissue inflammation and obesity via a family of microRNAs. Sci Transl Med. 2019;11(496):eaav1892.
Robertson I. The association of exercise, diet and other factors with owner-perceived obesity in privately owned dogs from metropolitan Perth. WA Prev Vet Med. 2003;58(1–2):75–83.
McGreevy P, Thomson P, Pride C, Fawcett A, Grassi T, Jones B. Prevalence of obesity in dogs examined by Australian veterinary practices and the risk factors involved. Vet Rec. 2005;156(22):695–702.
Colliard L, Paragon B-M, Lemuet B, Bénet J-J, Blanchard G. Prevalence and risk factors of obesity in an urban population of healthy cats. J Feline Med Surg. 2009;11(2):135–40.
Lund EM, Armstrong P, Kirk CA, Klausner J. Prevalence and risk factors for obesity in adult cats from private US veterinary practices. Intern J Appl Res Vet Med. 2005;3(2):88–96.
Chiang CF, Villaverde C, Chang WC, Fascetti AJ, Larsen JA. Prevalence, risk factors, and disease associations of overweight and obesity in cats that visited the Veterinary Medical Teaching Hospital at the University of California, Davis from January 2006 to December 2015. Top Companion Anim M. 2022;47:100620.
Pilla R, Suchodolski JS. The gut microbiome of dogs and cats, and the influence of diet. Vet Clin North Am Small Anim Pract. 2021;51(3):605–21.
Öhlund M, Palmgren M, Holst BS. Overweight in adult cats: a cross-sectional study. Acta Vet Scand. 2018;60(1):1–10.
Teng KT, McGreevy PD, Toribio J-AL, Raubenheimer D, Kendall K, Dhand NK. Risk factors for underweight and overweight in cats in metropolitan Sydney, Australia. Prev Vet Med. 2017;144:102–11.
Vandendriessche V, Picavet P, Hesta M. First detailed nutritional survey in a referral companion animal population. J Anim Physiol An N. 2017;101:4–14.
Pantalone KM, Hobbs TM, Chagin KM, Kong SX, Wells BJ, Kattan MW, et al. Prevalence and recognition of obesity and its associated comorbidities: cross-sectional analysis of electronic health record data from a large US integrated health system. BMJ Open. 2017;7(11):e017583.
De Godoy M, Swanson K. Companion animals symposium: nutrigenomics: using gene expression and molecular biology data to understand pet obesity. J Anim Sci. 2013;91(6):2949–64.
Speakman JR, Booles D, Butterwick R. Validation of dual energy X-ray absorptiometry (DXA) by comparison with chemical analysis of dogs and cats. Int J Obesity. 2001;25(3):439–47.
Laflamme D. Development and validation of a body condition score system for dogs. Canine Practice. 1997;25(5/6):13–8.
Santarossa A, Parr JM, Verbrugghe A. The importance of assessing body composition of dogs and cats and methods available for use in clinical practice. J Am Vet Med A. 2017;251(5):521–9.
Mawby DI, Bartges JW, d’Avignon A, Laflamme DP, Moyers TD, Cottrell T. Comparison of various methods for estimating body fat in dogs. J Am Anim Hosp Assoc. 2004;40(2):109–14.
German AJ, Holden SL, Moxham GL, Holmes KL, Hackett RM, Rawlings JM. A simple, reliable tool for owners to assess the body condition of their dog or cat. J Nutr. 2006;136(7):2031S-S2033.
National Research Council. Nutrient requirements of dogs and cats. Washington, DC: The National Academies Press; 2006.
Osto M, Lutz TA. Translational value of animal models of obesity—Focus on dogs and cats. Eur J Pharmacol. 2015;759:240–52.
German AJ. The growing problem of obesity in dogs and cats. J Nutr. 2006;136(7):1940S-S1946.
Jeusette I, Detilleux J, Cuvelier C, Istasse L, Diez M. Ad libitum feeding following ovariectomy in female Beagle dogs: effect on maintenance energy requirement and on blood metabolites. J Anim Physiol An N. 2004;88(3–4):117–21.
Rand JS, Fleeman LM, Farrow HA, Appleton DJ, Lederer R. Canine and feline diabetes mellitus: nature or nurture? J Nutr. 2004;134(8):2072S-S2080.
Wei A, Fascetti AJ, Kim K, Lee A, Graham JL, Havel PJ, et al. Early effects of neutering on energy expenditure in adult male cats. PLoS ONE. 2014;9(2):e89557.
Aa A. Diet quality concept. Nutrition. 2014;30(6):613–8.
Keller C, Liesegang A, Frey D, Wichert B. Metabolic response to three different diets in lean cats and cats predisposed to overweight. Bmc Vet Res. 2017;13:1–10.
Case LP, Daristotle L, Hayek MG, Raasch MF. Canine and feline nutrition: a resource for companion animal professionals. St Louis: Mosby Inc.; 2010.
Forrest R, Awawdeh L, Esam F, Pearson M, Waran N. Potential owner-related risk factors that may contribute to obesity in companion dogs in Aotearoa New Zealand. Animals. 2022;12(3):267.
Belsito K, Vester B, Keel T, Graves T, Swanson K. Impact of ovariohysterectomy and food intake on body composition, physical activity, and adipose gene expression in cats. J Anim Sci. 2009;87(2):594–602.
Machado B, Bruno C, Silva D, Barth J, Santos L, Alves M, et al. An overweight/obesity survey among dogs and cats attended at a veterinary teaching hospital during the second year of the COVID-19 pandemic. Arq Bras Med Vet Zootec. 2023;74:999–1006.
Taylor A. Obesity in felines. BSAVA Congress Proceedings 2020; Gloucester: BSAVA Library; 2020.
Millington HJ. Obesity: why is it a welfare issue? VNJ. 2020;36(3):101–3.
Cercato C, Fonseca FA. Cardiovascular risk and obesity. Diabetol Metab Syndr. 2019;11(1):74.
Raffan E. The big problem: battling companion animal obesity. Vet Rec. 2013;173(12):287–91.
Niaz K, Maqbool F, Khan F, Hassan FI, Momtaz S, Abdollahi M. Comparative occurrence of diabetes in canine, feline, and few wild animals and their association with pancreatic diseases and ketoacidosis with therapeutic approach. Vet World. 2018;11(4):410.
Verkest KR, Fleeman LM, Rand JS, Morton JM. Evaluation of beta-cell sensitivity to glucose and first-phase insulin secretion in obese dogs. Am J Vet Res. 2011;72(3):357–66.
Hoenig M, Alexander S, Holson J, Ferguson D. Influence of glucose dosage on interpretation of intravenous glucose tolerance tests in lean and obese cats. J Vet Intern Med. 2002;16(5):529–32.
Gayet C, Bailhache E, Dumon H, Martin L, Siliart B, Nguyen P. Insulin resistance and changes in plasma concentration of TNFα, IGF1, and NEFA in dogs during weight gain and obesity. J Anim Physiol An N. 2004;88(3–4):157–65.
Davison L, Herrtage M, Steiner J, Williams D, Catchpole B. Evidence of anti-insulin autoreactivity and pancreatic inflammation in newly diagnosed diabetic dogs. J Vet Intern Med. 2003;17:395.
Catchpole B, Kennedy L, Davison L, Ollier W. Canine diabetes mellitus: from phenotype to genotype. J Small Anim Pract. 2008;49(1):4–10.
Nelson R, Himsel C, Feldman E, Bottoms G. Glucose tolerance and insulin response in normal-weight and obese cats. Am J Vet Res. 1990;51(9):1357–62.
Ma Z, Westermark P, Westermark GT. Amyloid in human islets of Langerhans: immunologic evidence that islet amyloid polypeptide is modified in amyloidogenesis. Pancreas. 2000;21(2):212–8.
Verkest K, Rand J, Fleeman L, Morton J. Spontaneously obese dogs exhibit greater postprandial glucose, triglyceride, and insulin concentrations than lean dogs. Domest Anim Endocrin. 2012;42(2):103–12.
Pöppl AG, de Carvalho GLC, Vivian IF, Corbellini LG, González FHD. Canine diabetes mellitus risk factors: A matched case-control study. Res Vet Sci. 2017;114:469–73.
Center SA, Crawford MA, Guida L, Erb HN, King J. A retrospective study of 77 cats with severe hepatic lipidosis: 1975–1990. J Vet Intern Med. 1993;7(6):349–59.
Biourge V, Massat B, Groff J, Morris J, Rogers Q. Effects of protein, lipid, or carbohydrate supplementation on hepatic lipid accumulation during rapid weight loss in obese cats. Am J Vet Res. 1994;55(10):1406–15.
Postic C, Girard J. The role of the lipogenic pathway in the development of hepatic steatosis. Diabetes Metab J. 2008;34(6):643–8.
Center SA. Feline hepatic lipidosis. Vet Clin N Am-Small. 2005;35(1):225–69.
Minamoto T, Walzem RL, Hamilton AJ, Hill SL, Payne HR, Lidbury JA, et al. Altered lipoprotein profiles in cats with hepatic lipidosis. J Feline Med Surg. 2019;21(4):363–72.
Minamoto T. Lipoprotein profiling in dogs and cats with gastrointestinal disease or hepatic disease. Doctoral dissertation, Texas A & M University. 2018. Available electronically from https://hdl.handle.net/1969.1/173446.
Armstrong PJ, Blanchard G. Hepatic lipidosis in cats. Vet Clin N Am-Small. 2009;39(3):599–616.
Dimski DS. Feline hepatic lipidosis. Seminars in veterinary medicine and surgery (small animal); 1997. p. 28–33.
Xenoulis PG, Steiner JM. Lipid metabolism and hyperlipidemia in dogs. Vet J. 2010;183(1):12–21.
Valtolina C, Favier RP. Feline hepatic lipidosis. Vet Clin N Am-Small. 2017;47(3):683–702.
Chandler ML. Impact of obesity on cardiopulmonary disease. Vet Clin N Am-Small. 2016;46(5):817–30.
Bouthegourd J-C, Kelly M, Clety N, Tardif S, Smeets D. Effects of weight loss on heart rate normalization and increase in spontaneous activity in moderately exercised overweight dogs. Int J Appl Res Vet Med. 2009;7(4):153.
Mehlman E, Bright J, Jeckel K, Porsche C, Veeramachaneni D, Frye M. Echocardiographic evidence of left ventricular hypertrophy in obese dogs. J Vet Intern Med. 2013;27(1):62–8.
Ware WA, Bonagura JD, Scansen BA. Myocardial diseases of the cat. In: Ware WA, Bonagura JD, editors. cardiovascular disease in companion animals: dog, cat, and horse. 2nd ed. Boca Raton: CRC Press; 2021. p. 649–94.
Hatem SN, Redheuil A, Gandjbakhch E. Cardiac adipose tissue and atrial fibrillation: the perils of adiposity. Cardiovasc Res. 2016;109(4):502–9.
Gomes VdR, Ariza PC, Borges NC, Schulz FJ, Fioravanti MCS. Risk factors associated with feline urolithiasis. Vet Res Commun. 2018;42:87–94.
Burggraaf ND, Westgeest DB, Corbee RJ. Analysis of 7866 feline and canine uroliths submitted between 2014 and 2020 in the Netherlands. Res Vet Sci. 2021;137:86–93.
Fritsch D, Allen T, Dodd C, Jewell D, Sixby K, Leventhal P, et al. Dose-titration effects of fish oil in osteoarthritic dogs. J Vet Intern Med. 2010;24(5):1020–6.
Johnson KA, Lee AH, Swanson KS. Nutrition and nutraceuticals in the changing management of osteoarthritis for dogs and cats. J Am Vet Med A. 2020;256(12):1335–41.
Wang T, He C. Pro-inflammatory cytokines: the link between obesity and osteoarthritis. Cytokine Growth Factor Rev. 2018;44:38–50.
Song Z, Xiaoli AM, Yang F. Regulation and metabolic significance of de novo lipogenesis in adipose tissues. Nutrients. 2018;10(10):1383.
Ameer F, Scandiuzzi L, Hasnain S, Kalbacher H, Zaidi N. De novo lipogenesis in health and disease. Metabolism. 2014;63(7):895–902.
Bergen WG, Mersmann HJ. Comparative aspects of lipid metabolism: impact on contemporary research and use of animal models. J Nutr. 2005;135(11):2499–502.
Richard MJ, Holck JT, Beitz DC. Lipogenesis in liver and adipose tissue of the domestic cat (Felis domestica). Comp Biochem Physiol. 1989;93(3):561–4.
Kruitwagen HS, Oosterhoff LA, Vernooij IG, Schrall IM, van Wolferen ME, Bannink F, et al. Long-term adult feline liver organoid cultures for disease modeling of hepatic steatosis. Stem Cell Rep. 2017;8(4):822–30.
Corbee RJ, Van Everdingen DL, Kooistra HS, Penning LC. Fibroblast growth factor-21 (FGF21) analogs as possible treatment options for diabetes mellitus in veterinary patients. Front Vet Sci. 2023;9:1086987.
Wilkins C, Long RC, Waldron M, Ferguson DC, Hoenig M. Assessment of the influence of fatty acids on indices of insulin sensitivity and myocellular lipid content by use of magnetic resonance spectroscopy in cats. Am J Vet Res. 2004;65(8):1090–9.
Dash S, Xiao C, Morgantini C, Lewis GF. New insights into the regulation of chylomicron production. Annu Rev Nutr. 2015;35:265–94.
Choi SH, Ginsberg HN. Increased very low density lipoprotein (VLDL) secretion, hepatic steatosis, and insulin resistance. Trends Endocrin Met. 2011;22(9):353–63.
Bauer JE. Lipoprotein-mediated transport of dietary and synthesized lipids and lipid abnormalities of dogs and cats. J Am Vet Med A. 2004;224(5):668–75.
Watson T, Barrie J. Lipoprotein metabolism and hyperlipidaemia in the clog and cat: A review. J Small Anim Pract. 1993;34(10):479–87.
Fielding CJ, Fielding PE. Molecular physiology of reverse cholesterol transport. J Lipid Res. 1995;36(2):211–28.
Xenoulis P, Steiner J. Canine hyperlipidaemia. J Small Anim Pract. 2015;56(10):595–605.
Johnson MC. Hyperlipidemia disorders in dogs. Compendium. 2005;27:361–70.
Hoenig M. The cat as a model for human nutrition and disease. Curr Opin Clin Nutr. 2006;9(5):584–8.
Sennello KA, Schulman RL, Prosek R, Siegel AM. Systolic blood pressure in cats with diabetes mellitus. J Am Vet Med A. 2003;223(2):198–201.
Hoenig M, Thomaseth K, Waldron M, Ferguson D. Insulin sensitivity, fat distribution, and adipocytokine response to different diets in lean and obese cats before and after weight loss. Am J Physiol-Reg I. 2007;292(1):R227–34.
German A, Hervera M, Hunter L, Holden S, Morris P, Biourge V, et al. Improvement in insulin resistance and reduction in plasma inflammatory adipokines after weight loss in obese dogs. Domest Anim Endocrin. 2009;37(4):214–26.
Ishioka K, Omachi A, Sasaki N, Kimura K, Saito M. Feline adiponectin: molecular structures and plasma concentrations in obese cats. J Vet Med Sci. 2009;71(2):189–94.
Zini E, Linscheid P, Franchini M, Kaufmann K, Monnais E, Kutter AP, et al. Partial sequencing and expression of genes involved in glucose metabolism in adipose tissues and skeletal muscle of healthy cats. Vet J. 2009;180(1):66–70.
Kaser S, Tatarczyk T, Stadlmayr A, Ciardi C, Ress C, Tschoner A, et al. Effect of obesity and insulin sensitivity on adiponectin isoform distribution. Eur J Clin Invest. 2008;38(11):827–34.
Lusby AL, Kirk CA, Bartges JW. The role of key adipokines in obesity and insulin resistance in cats. J Am Vet Med A. 2009;235(5):518–22.
Verkest K, Rand J, Fleeman L, Morton J, Richards A, Rose F, et al. Distinct adiponectin profiles might contribute to differences in susceptibility to type 2 diabetes in dogs and humans. Domest Anim Endocrin. 2011;41(2):67–73.
Karbowska J, Kochan Z. Role of adiponectin in the regulation of carbohydrate and lipid metabolism. J Physiol Pharmacol. 2006;57:103.
Ouchi N, Kihara S, Arita Y, Okamoto Y, Maeda K, Kuriyama H, et al. Adiponectin, an adipocyte-derived plasma protein, inhibits endothelial NF-κB signaling through a cAMP-dependent pathway. Circulation. 2000;102(11):1296–301.
Kadowaki T, Yamauchi T. Adiponectin and adiponectin receptors. Endocr Rev. 2005;26(3):439–51.
Li S, Shin HJ, Ding EL, van Dam RM. Adiponectin levels and risk of type 2 diabetes: a systematic review and meta-analysis. JAMA. 2009;302(2):179–88.
Verkest K, Fleeman L, Morton J, Ishioka K, Rand J. Compensation for obesity-induced insulin resistance in dogs: assessment of the effects of leptin, adiponectin, and glucagon-like peptide-1 using path analysis. Domest Anim Endocrin. 2011;41(1):24–34.
Wakshlag JJ, Struble AM, Levine CB, Bushey JJ, Laflamme DP, Long GM. The effects of weight loss on adipokines and markers of inflammation in dogs. Brit J Nutr. 2011;106(S1):S11–4.
Patra D, Banerjee D, Ramprasad P, Roy S, Pal D, Dasgupta S. Recent insights of obesity-induced gut and adipose tissue dysbiosis in type 2 diabetes. Front Mol Biosci. 2023;10:1224982.
Macedo HT, Rentas MF, Vendramini THA, Macegoza MV, Amaral AR, Jeremias JT, et al. Weight-loss in obese dogs promotes important shifts in fecal microbiota profile to the extent of resembling microbiota of lean dogs. Anim Microbiome. 2022;4:6.
Chun JL, Ji SY, Lee SD, Lee YK, Kim B, Kim KH. Difference of gut microbiota composition based on the body condition scores in dogs. J Anim Sci and technology. 2020;62(2):239–46.
Park HJ, Lee SE, Kim HB, Isaacson RE, Seo KW, Song KH. Association of obesity with serum leptin, adiponectin, and serotonin and gut microflora in beagle dogs. J Vet Intern Med. 2015;29(1):43–50.
Handl S, German AJ, Holden SL, Dowd SE, Steiner JM, Heilmann RM, et al. Faecal microbiota in lean and obese dogs. Fems Microbiol Ecol. 2013;84(2):332–43.
Forster GM, Stockman J, Noyes N, Heuberger AL, Broeckling CD, Bantle CM, et al. A comparative study of serum biochemistry, metabolome and microbiome parameters of clinically healthy, normal weight, overweight, and obese companion dogs. Top Companion Anim M. 2018;33(4):126–35.
Ma X, Brinker E, Graff EC, Cao W, Gross AL, Johnson AK, et al. Whole-genome shotgun metagenomic sequencing reveals distinct gut microbiome signatures of obese cats. Microbiol Spectr. 2022;10(3):e0083722.
Kieler IN, Osto M, Hugentobler L, Puetz L, Gilbert MTP, Hansen T, et al. Diabetic cats have decreased gut microbial diversity and a lack of butyrate producing bacteria. Sci Rep-Uk. 2019;9(1):4822.
Pallotto MR, de Godoy MRC, Holscher HD, Buff PR, Swanson KS. Effects of weight loss with a moderate-protein, high-fiber diet on body composition, voluntary physical activity, and fecal microbiota of obese cats. Am J Vet Res. 2018;79(2):181–90.
Fischer MM, Kessler AM, Kieffer DA, Knotts TA, Kim K, Wei A, et al. Effects of obesity, energy restriction and neutering on the faecal microbiota of cats. The Brit J Nutr. 2017;118(7):513–24.
Tal M, Weese JS, Gomez DE, Hesta M, Steiner JM, Verbrugghe A. Bacterial fecal microbiota is only minimally affected by a standardized weight loss plan in obese cats. BMC Vet Res. 2020;16:112.
Mondo E, Marliani G, Accorsi PA, Cocchi M, Di Leone A. Role of gut microbiota in dog and cat’s health and diseases. Open Vet J. 2019;9(3):253–8.
Vourakis M, Mayer G, Rousseau G. The role of gut microbiota on cholesterol metabolism in atherosclerosis. Int J Mol Sci. 2021;22(15):8074.
Zhu L, Wang J, Ding X, Bai S, Zeng Q, Xuan Y, et al. Serum trimethylamine-N-oxide and gut microbiome alterations are associated with cholesterol deposition in the liver of laying hens fed with rapeseed meal. Anim Nutr. 2021;7(4):1258–70.
Agus A, Clement K, Sokol H. Gut microbiota-derived metabolites as central regulators in metabolic disorders. Gut. 2021;70(6):1174–82.
Silva YP, Bernardi A, Frozza RL. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front Endocrinol (Lausanne). 2020;11:25.
Fröhlich EE, Farzi A, Mayerhofer R, Reichmann F, Jačan A, Wagner B, et al. Cognitive impairment by antibiotic-induced gut dysbiosis: Analysis of gut microbiota-brain communication. Brain Behav Immun. 2016;56:140–55.
Loh JS, Mak WQ, Tan LKS, Ng CX, Chan HH, Yeow SH, et al. Microbiota-gut-brain axis and its therapeutic applications in neurodegenerative diseases. Signal Transduct Tar. 2024;9(1):37.
Grenham S, Clarke G, Cryan JF, Dinan TG. Brain-gut-microbe communication in health and disease. Front Physiol. 2011;2:94.
Hand MS, Lewis LD. Small animal clinical nutrition. Topeka, Kansas: Mark Morris Institute; 2000.
Zoran DL. The carnivore connection to nutrition in cats. J Am Vet Med A. 2002;221(11):1559–67.
Barrett EJ, Ferrannini E, Gusberg R, Bevilacqua S, DeFronzo RA. Hepatic and extrahepatic splanchnic glucose metabolism in the postabsorptive and glucose fed dog. Metabolism. 1985;34(5):410–20.
Bagavan Reddy P, Namdeo S, Vaani Shreeya T, Das T, Kumar M, Katam D. Nutritional management of liver diseases in dogs and cats. Pharma Innov J. 2023;12(1):1352–9.
Rogers Q, Morris J, Freeland R. Lack of hepatic enzymatic adaptation to low and high levels of dietary protein in the adult cat. Enzyme. 1977;22(5):348–56.
Morris JG. Idiosyncratic nutrient requirements of cats appear to be diet-induced evolutionary adaptations. Nutr Res Rev. 2002;15(1):153–68.
Brooks D, Churchill J, Fein K, Linder D, Michel KE, Tudor K, et al. 2014 AAHA weight management guidelines for dogs and cats. J Am Anim Hosp Assoc. 2014;50(1):1–11.
Gaylord L, Remillard R, Saker K. Risk of nutritional deficiencies for dogs on a weight loss plan. J Small Anim Pract. 2018;59(11):695–703.
Loftus JP, Wakshlag JJ. Canine and feline obesity: a review of pathophysiology, epidemiology, and clinical management. Vet Med. 2014;6:49–60. https://doi.org/10.2147/VMRR.S40868.
Wakshlag JJ, Struble AM, Warren BS, Maley M, Panasevich MR, Cummings KJ, et al. Evaluation of dietary energy intake and physical activity in dogs undergoing a controlled weight-loss program. J Am Vet Med A. 2012;240(4):413–9.
Hooda S, Vester Boler BM, Kerr KR, Dowd SE, Swanson KS. The gut microbiome of kittens is affected by dietary protein:carbohydrate ratio and associated with blood metabolite and hormone concentrations. The Brit J Nutr. 2013;109(9):1637–46.
Li Q, Lauber CL, Czarnecki-Maulden G, Pan Y, Hannah SS. Effects of the dietary protein and carbohydrate ratio on gut microbiomes in dogs of different body conditions. MBio. 2017;8(1):e01703–16.
Coelho LP, Kultima JR, Costea PI, Fournier C, Pan Y, Czarnecki-Maulden G, et al. Similarity of the dog and human gut microbiomes in gene content and response to diet. Microbiome. 2018;6:72.
Bierer TL, Bui LM. High-protein low-carbohydrate diets enhance weight loss in dogs. J Nutr. 2004;134(8 Suppl):2087s-s2089.
Laflamme DP, Hannah SS. Increased dietary protein promotes fat loss and reduces loss of lean body mass during weight loss in cats. Intern J Appl Res Vet Med. 2005;3(2):62–8.
Kushner RF, Doerfler B. Low-carbohydrate, high-protein diets revisited. Curr Opin Gastroen. 2008;24(2):198–203.
Wei A, Fascetti AJ, Liu KJ, Villaverde C, Green AS, Manzanilla EG, et al. Influence of a high-protein diet on energy balance in obese cats allowed ad libitum access to food. J Anim Physiol An N. 2011;95(3):359–67.
Vasconcellos RS, Borges NC, Goncalves KN, Canola JC, de Paula FJ, Malheiros EB, et al. Protein intake during weight loss influences the energy required for weight loss and maintenance in cats. J Nutr. 2009;139(5):855–60.
Blanchard G, Nguyen P, Gayet C, Leriche I, Siliart B, Paragon B-M. Rapid weight loss with a high-protein low-energy diet allows the recovery of ideal body composition and insulin sensitivity in obese dogs. J Nutr. 2004;134(8):2148S-S2150.
Blachier F, Beaumont M, Portune KJ, Steuer N, Lan A, Audebert M, et al. High-protein diets for weight management: Interactions with the intestinal microbiota and consequences for gut health. A position paper by the my new gut study group. Clin Nutr (Edinburgh, Scotland). 2019;38(3):1012–22.
Singh RK, Chang HW, Yan D, Lee KM, Ucmak D, Wong K, et al. Influence of diet on the gut microbiome and implications for human health. J Transl Med. 2017;15(1):73.
Lopez-Legarrea P, Fuller NR, Zulet MA, Martinez JA, Caterson ID. The influence of Mediterranean, carbohydrate and high protein diets on gut microbiota composition in the treatment of obesity and associated inflammatory state. Asia Pac J Clin Nutr. 2014;23(3):360–8.
Yao CK, Muir JG, Gibson PR. Review article: insights into colonic protein fermentation, its modulation and potential health implications. Aliment Pharm Ther. 2016;43(2):181–96.
Reaven GM. Insulin resistance and compensatory hyperinsulinemia: role in hypertension, dyslipidemia, and coronary heart disease. Am Heart J. 1991;121(4 Pt 2):1283–8.
El Hafidi M, Valdez R, Baños G. Possible relationship between altered fatty acid composition of serum, platelets, and aorta and hypertension induced by sugar feeding in rats. Clin Exp Hypertens. 2000;22(1):99–108.
Nguyen PG, Dumon HJ, Siliart BS, Martin LJ, Sergheraert R, Biourge VC. Effects of dietary fat and energy on body weight and composition after gonadectomy in cats. Am J Vet Res. 2004;65(12):1708–13.
Tomas J, Mulet C, Saffarian A, Cavin JB, Ducroc R, Regnault B, et al. High-fat diet modifies the PPAR-γ pathway leading to disruption of microbial and physiological ecosystem in murine small intestine. P Natl Acad Sci USA. 2016;113(40):E5934–43.
Kilburn LR, Koester LR, Schmitz-Esser S, Serão NVL, Rossoni Serão MC. High-fat diets led to OTU-level shifts in fecal samples of healthy adult dogs. Front Microbiol. 2020;11:564160.
Fu Y, Wang Y, Gao H, Li D, Jiang R, Ge L, et al. Associations among Dietary omega-3 polyunsaturated fatty acids, the gut microbiota, and intestinal immunity. Mediators Inflamm. 2021;2021:8879227.
Miyamoto J, Igarashi M, Watanabe K, Karaki SI, Mukouyama H, Kishino S, et al. Gut microbiota confers host resistance to obesity by metabolizing dietary polyunsaturated fatty acids. Nat Commun. 2019;10:4007.
Hill C, Guarner F, Reid G, Gibson GR, Merenstein DJ, Pot B, et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastro Hepat. 2014;11(8):506–14.
Joung H, Chu J, Kim BK, Choi IS, Kim W, Park TS. Probiotics ameliorate chronic low-grade inflammation and fat accumulation with gut microbiota composition change in diet-induced obese mice models. Appl Microbiol Biot. 2021;105(3):1203–13.
Giannoudaki E, Hernandez-Santana YE, Mulfaul K, Doyle SL, Hams E, Fallon PG, et al. Interleukin-36 cytokines alter the intestinal microbiome and can protect against obesity and metabolic dysfunction. Nat Commun. 2019;10:4003.
Molina-Tijeras JA, Diez-Echave P, Vezza T, Hidalgo-Garcia L, Ruiz-Malagon AJ, Rodriguez-Sojo MJ, et al. Lactobacillus fermentum CECT5716 ameliorates high fat diet-induced obesity in mice through modulation of gut microbiota dysbiosis. Pharmacol Res. 2021;167:105471.
Kathrani A, Larsen JA, Kass PH, Fascetti AJ. Effect of short-term probiotic Enterococcus faecium SF68 dietary supplementation in overweight and obese cats without comorbidities. Vet Rec Open. 2016;3(1):e000164.
Kang A, Kwak MJ, Lee DJ, Lee JJ, Kim MK, Song M, et al. Dietary supplementation with probiotics promotes weight loss by reshaping the gut microbiome and energy metabolism in obese dogs. Microbiol Spectr. 2024;12(3):e0255223.
Marcinakova M, Simonova M, Strompfova V, Laukova A. Oral application of Enterococcus faecium strain EE3 in healthy dogs. Folia Microbiol (Praha). 2006;51(3):239–42.
Alexander C, Cross TL, Devendran S, Neumer F, Theis S, Ridlon JM, et al. Effects of prebiotic inulin-type fructans on blood metabolite and hormone concentrations and faecal microbiota and metabolites in overweight dogs. Br J Nutr. 2018;120(6):711–20.
Lee HJ, Cho JH, Cho WJ, Gang SH, Park SH, Jung BJ, et al. Effects of synbiotic preparation containing Lactobacillus gasseri BNR17 on body fat in obese dogs: A pilot study. Animals. 2022;12(5):642.
Jewell DE, Jackson MI, Cochrane CY, Badri DV. Feeding fiber-bound polyphenol ingredients at different levels modulates colonic postbiotics to improve gut health in dogs. Animals. 2022;12(5):627.
Jewell DE, Jackson MI, Cochrane CY, Badri DV. Feeding fiber-bound polyphenol ingredients at different levels modulates colonic postbiotics to improve gut health in cats. Animals. 2022;12(13):1654.
Bybee SN, Scorza AV, Lappin MR. Effect of the probiotic Enterococcus faecium SF68 on presence of diarrhea in cats and dogs housed in an animal shelter. J Vet Intern Med. 2011;25(4):856–60.
Lappin MR, Veir JK, Satyaraj E, Czarnecki-Maulden G. Pilot study to evaluate the effect of oral supplementation of Enterococcus faecium SF68 on cats with latent feline herpesvirus 1. J Feline Med Surg. 2009;11(8):650–4.
Grzeskowiak L, Endo A, Beasley S, Salminen S. Microbiota and probiotics in canine and feline welfare. Anaerobe. 2015;34:14–23.
Kumar S, Pattanaik AK, Sharma S, Gupta R, Jadhav SE, Dutta N. Comparative assessment of canine-origin Lactobacillus johnsonii CPN23 and dairy-origin Lactobacillus acidophillus NCDC 15 for nutrient digestibility, faecal fermentative metabolites and selected gut health indices in dogs. J Nutr Sci. 2017;6:e38.
Gibson GR, Hutkins R, Sanders ME, Prescott SL, Reimer RA, Salminen SJ, et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastro Hepat. 2017;14(8):491–502.
Respondek F, Gerard P, Bossis M, Boschat L, Bruneau A, Rabot S, et al. Short-chain fructo-oligosaccharides modulate intestinal microbiota and metabolic parameters of humanized gnotobiotic diet induced obesity mice. PLoS ONE. 2013;8(8):e71026.
Wang H, Zhang X, Wang S, Li H, Lu Z, Shi J, et al. Mannan-oligosaccharide modulates the obesity and gut microbiota in high-fat diet-fed mice. Food Funct. 2018;9(7):3916–29.
Li X, Guo J, Ji K, Zhang P. Bamboo shoot fiber prevents obesity in mice by modulating the gut microbiota. Sci Rep-Uk. 2016;6:32953.
Lee AH, Jha AR, Do S, Scarsella E, Shmalberg J, Schauwecker A, et al. Dietary enrichment of resistant starches or fibers differentially alter the feline fecal microbiome and metabolite profile. Anim Microbiome. 2022;4:61.
Wernimont SM, Radosevich J, Jackson MI, Ephraim E, Badri DV, MacLeay JM, et al. The effects of nutrition on the gastrointestinal microbiome of cats and dogs: impact on health and disease. Front Microbiol. 2020;11:1266.
Swanson KS, Gibson GR, Hutkins R, Reimer RA, Reid G, Verbeke K, et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat Rev Gastro Hepat. 2020;17(11):687–701.
Kosuwon P, Lao-Araya M, Uthaisangsook S, Lay C, Bindels J, Knol J, et al. A synbiotic mixture of scGOS/lcFOS and Bifidobacterium breve M-16V increases faecal Bifidobacterium in healthy young children. Beneficial microbes. 2018;9(4):541–52.
Li HY, Zhou DD, Gan RY, Huang SY, Zhao CN, Shang A, et al. Effects and mechanisms of probiotics, prebiotics, synbiotics, and postbiotics on metabolic diseases targeting gut microbiota: A Narrative Review. Nutrients. 2021;13(9):3211.
Amabebe E, Robert FO, Agbalalah T, Orubu ESF. Microbial dysbiosis-induced obesity: role of gut microbiota in homoeostasis of energy metabolism. The Brit J Nutr. 2020;123(10):1127–37.
da Silva TF, Casarotti SN, de Oliveira GLV, Penna ALB. The impact of probiotics, prebiotics, and synbiotics on the biochemical, clinical, and immunological markers, as well as on the gut microbiota of obese hosts. Crit Rev Food Sci. 2021;61(2):337–55.
Salminen S, Collado MC, Endo A, Hill C, Lebeer S, Quigley EMM, et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat Rev Gastro Hepat. 2021;18(9):649–67.
Jewell DE, Jackson MI. Dietary fatty acids change circulating fatty acids, microbial putrefactive postbiotics and betaine status in the cat. Animals. 2020;10(12):2310.
Jackson MI, Jewell DE. Docosahexaenoate-enriched fish oil and medium chain triglycerides shape the feline plasma lipidome and synergistically decrease circulating gut microbiome-derived putrefactive postbiotics. PLoS ONE. 2020;15(3):e0229868.
Chaitman J, Ziese AL, Pilla R, Minamoto Y, Blake AB, Guard BC, et al. Fecal microbial and metabolic profiles in dogs with acute diarrhea receiving either fecal microbiota transplantation or Oral Metronidazole. Front Vet Sci. 2020;7:192.
Borody T. Chapter 18 - Fecal Microbiota Transplantation: Treatment of the Gut Microbiome. In: Eslick GD, editor. Gastrointestinal diseases and their associated infections. Philadelphia: Elsevier; 2019. p. 249–61.
Li KR, Zhou XX, Zhong ZJ, Liu HF, Lie M, Peng GN, et al. Indications for canine fecal microbiota transplantation. Thai J Vet Med. 2022;52(1):13–21.
Sugita K, Yanuma N, Ohno H, Takahashi K, Kawano K, Morita H, et al. Oral faecal microbiota transplantation for the treatment of Clostridium difficile-associated diarrhoea in a dog: a case report. BMC Vet Res. 2019;15:11.
Niina A, Kibe R, Suzuki R, Yuchi Y, Teshima T, Matsumoto H, et al. Improvement in clinical symptoms and fecal microbiome after fecal microbiota transplantation in a dog with inflammatory bowel disease. Vet Med. 2019;10:197–201.
Bryan CE, Cade JC, Mackin AJ, Sullivant AM. Evaluation of a structured individualised protocol as a potential cost-effective diagnostic and therapeutic approach to chronic diarrhoea in the dog. Vet Med Sci. 2019;5(2):210–21.
Bottero E, Benvenuti E, Ruggiero P. Fecal microbiota transplantation (FMT) in 16 dogs with idiopatic IBD. Veterinaria. 2017;31:31–45.
Pereira GQ, Gomes LA, Santos IS, Alfieri AF, Weese JS, Costa MC. Fecal microbiota transplantation in puppies with canine parvovirus infection. J Vet Intern Med. 2018;32(2):707–11.
Gal A, Barko PC, Biggs PJ, Gedye KR, Midwinter AC, Williams DA, et al. One dog’s waste is another dog’s wealth: A pilot study of fecal microbiota transplantation in dogs with acute hemorrhagic diarrhea syndrome. PLoS ONE. 2021;16(4):e0250344.
Li K, Yang J, Zhou X, Wang H, Ren Y, Huang Y, et al. The mechanism of important components in canine fecal microbiota transplantation. Vet Sci. 2022;9(12):695.
Furmanski S, Mor T. First case report of fecal microbiota transplantation in a cat in Israel. Isr J Vet Med. 2017;72(3):35–41.
Weese JS, Costa MC, Webb JA. Preliminary clinical and microbiome assessment of stool transplantation in the dog and cat. J Vet Intern Med. 2013;27(3):705.
Rojas C, Entrolezo Z, Jarett JK, Jospin G, Kingsbury D, Martin A, et al. Microbiome responses to fecal microbiota transplantation in cats with chronic digestive issues. Vet Sci. 2023;10(9):561.
Zhu L, Fu J, Xiao X, Wang F, Jin M, Fang W, et al. Faecal microbiota transplantation-mediated jejunal microbiota changes halt high-fat diet-induced obesity in mice via retarding intestinal fat absorption. Microb Biotechnol. 2022;15(1):337–52.
Jabs S, Biton A, Bécavin C, Nahori M-A, Ghozlane A, Pagliuso A, et al. Impact of the gut microbiota on the m6A epitranscriptome of mouse cecum and liver. Nat Commun. 2020;11:1344.
Samblas M, Milagro FI, Martínez A. DNA methylation markers in obesity, metabolic syndrome, and weight loss. Epigenetics. 2019;14(5):421–44.
Ridaura VK, Faith JJ, Rey FE, Cheng J, Duncan AE, Kau AL, et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science. 2013;341(6150):1241214.
Astrup A, Raben A, Geiker N. The role of higher protein diets in weight control and obesity-related comorbidities. Int J Obes. 2015;39:721–6.
Alexander JE, Colyer A, Morris PJ. The effect of reducing energy density, via the addition of water to dry diet, on body weight and activity in dogs. J Nutr Sci. 2017;6:e42.
Vadiveloo M, Parker H, Raynor H. Increasing low-energy-dense foods and decreasing high-energy-dense foods differently influence weight loss trial outcomes. Int J Obesity (2005). 2018;42(3):479–86.
Acknowledgements
This work was funded by research grants from the Jiangsu Provincial Double-Innovation Team Program (JSSCTD202147), Nutrition and Care of Maternal and Child Research Fund Project of the Biostime Institute of Nutrition and Care (Grant No.2022BINCMCF006), the Postgraduate Research & Practice Innovation Program of Jiangsu Province (KYCX22_3527), the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Author information
Authors and Affiliations
Contributions
DC, TS, and XL conceptualized and structured the manuscript. YL, JZ, KL, XS, JX, and XL prepared the original draft. DC, TS, XL, XX, SL and HL co-wrote the content. All authors contributed to the manuscript's editing, revision, and finalization. All authors have read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Li, K., Xiao, X., Li, Y. et al. Insights into the interplay between gut microbiota and lipid metabolism in the obesity management of canines and felines. J Animal Sci Biotechnol 15, 114 (2024). https://doi.org/10.1186/s40104-024-01073-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40104-024-01073-w