
Abstract
By hibernating, animals can reduce their energy requirements by at least ninety percent and survive for many months while slowly catabolizing body lipid reserves. Hibernation constitutes an effective strategy of animals in order to survive cold environments and limited availability of food. In this work, we investigate a periodic impulsive switched predator-prey system with hibernation and birth pulse. We firstly obtain the conditions of the globally asymptotically stable prey-extinction boundary periodic solution of the investigated system. Secondly, we obtain the permanent conditions of the investigated system. Finally, numerical analysis is presented to illustrate the results. Our results provide reliable tactic basis for the practical biological economics management.
Similar content being viewed by others

1 Introduction
Hibernation allows small mammals to minimize metabolic energy costs at a time when a scarcity of food and cold environmental temperatures endanger normal life. By hibernating, animals can reduce their energy requirements by at least ninety percent and survive for many months while slowly catabolizing body lipid reserves [1]. Hibernation constitutes an effective strategy of animals in order to survive cold environments and limited availability of food [2].
Biological resources are renewable resources. Clark [3] considered the economic and biological aspects of renewable resources management. In recent years, the optimal management of renewable resources, which has a direct relationship to sustainable development, has been studied extensively by many authors [4–8]. Especially, the predator-prey models with harvesting have been investigated by many papers [9–14]. Li and Wang [14] considered dynamics of an Ivlev-type predator-prey system with constant rate harvesting
where \(x(t)\) denotes the density of the prey population at time t. \(y(t)\) denotes the density of the predator population at time t. The biological meanings of the parameters can be found in [14]. There is a vast amount of literature on the applications of impulsive differential equations to investigate predator-prey systems and SI system [15–22]. Jiao et al. [20] also considered a three-dimensional predator-prey model with impulsive diffusion and harvesting on predator as follows:
where they suppose that system (1.2) is composed of two patches connected by diffusion and occupied by species \(x(t)\) and \(y_{i}(t)\) (\(i = 1, 2\)). \(x(t)\) and \(y_{i}(t)\) (\(i = 1, 2\)) represent the density of prey species in Patch 1 and the density of predator species in Patch i (\(i = 1, 2\)), respectively. The biological meanings of the parameters can be found in [20].
The hibernation constitutes an effective strategy of animals in order to survive cold environments and limited availability of food, it is a universal phenomenon in biological world. However, there are few papers considering and investigating mathematical models with winter hibernation. In this paper, we introduce the phenomenon of hibernation and focus on a periodic impulsive switched predator-prey system with hibernation and birth pulse.
The organization of this paper is as follows. In the next section, we introduce the model and background concepts. In Section 3, some important lemmas are presented. In Section 4, we give the globally asymptotically stable conditions of a prey-extinction periodic solution of system (2.1) and the permanent condition of system (2.1). In Section 5, a brief discussion and the simulations are given to conclude this work.
2 The model
In this section, a periodic impulsive switched predator-prey system with hibernation and birth pulse is modeled by the nonlinear impulsive differential equation
where the total population is divided into two subpopulations: prey population \(x(t)\) and predator population \(y(t)\). It is assumed that the prey population is hibernator, and the predators depend on prey as their source of food; if there is no prey, the predator population will disappear. The impulsive period is divided into hibernation and non-hibernation. The predator population is birth pulse in their non-hibernation of the prey population. Intrinsic rate of natural increase and density dependence rate of prey population are denoted by a and b, respectively. \(d_{1}\) is the natural death rate of the predator population. The predator population consumes the prey population with predation coefficients \(\beta_{1}\) in the non-hibernation period of prey population. \(k_{1}\) is the rate of conversion of nutrients into the predator population. \(d_{2}>0\) is the natural death rate of the prey population in the hibernation period of prey population. The predator population is birth pulse as intrinsic rate of natural increase and density dependence rate of prey population are denoted by \(a_{1}\) and \(b_{1}\) respectively at moments \(t=(n+l)\tau\), \(0< l<1\), \(n\in Z^{+}\), that is, the predator population is born in non-hibernation of prey population, and the predator population cannot have birth ability in hibernation of prey population. \(d_{3}>0\) is the natural death rate of the predator population in the hibernation period of prey population. The predator population consumes prey population with predation coefficients \(\beta_{2}\) in the hibernation period of prey population. \(k_{2}\) is the rate of conversion of nutrients into the predator population in the hibernation period of prey population. \(0<\mu<1\) is the harvesting coefficient of the prey population at moments \(t=(n+1)\tau\), \(n\in Z^{+}\). \(0<\mu_{1}<1\) is the harvesting coefficient of the predator population at moments \(t=(n+1)\tau\), \(n\in Z^{+}\). Time interval \((n\tau,(n+l)\tau]\) is the non-hibernation of prey population. Time interval \(((n+l)\tau,(n+1)\tau]\) is the hibernation of prey population.
3 Some lemmas
The solution of system (2.1), denoted by \(X(t)=(x(t),y(t))^{T}\), is a non-smooth function \(X: R_{+}\rightarrow R_{+}^{2}\), \(X(t)\) is continuous on \((n\tau,(n+l)\tau]\) and \(((n+l)\tau,(n+1)\tau ]\), \(n\in Z_{+}\). \(X(n\tau^{+})=\lim_{t\rightarrow n\tau^{+} }X(t)\) and \(X((n+l)\tau ^{+})=\lim_{t\rightarrow(n+l)\tau^{+} }X(t)\) exist. Obviously the global existence and uniqueness of solutions of system (2.1) are guaranteed by the smoothness properties of f, which denotes the mapping defined by the right-hand side of system (2.1) (see Lakshmikantham et al. [23]).
Lemma 3.1
For each solution \((x(t),y(t))\) of system (2.1), there exists a constant \(M>0\) such that \(x(t)\leq M\), \(y(t)\leq M \) with all t large enough.
Proof
Define \(V(t)=kx(t)+y(t)\), \(k=\max\{k_{1},k_{2}\}\), and \(d=\min\{d_{1},d_{2},d_{3}\}\). When \(t\in( n\tau,(n+l)\tau]\), we have
when \(t\in( (n+l)\tau,(n+1)\tau]\), we have
Then, taking \(\delta=\frac{k(a+d)^{2}}{4b}\), when \(t\neq n\tau\), \(t\neq (n+l)\tau\), we have
When \(t=(n+l)\tau\), \(V((n+l)\tau^{+})= V((n+l)\tau)-b[y(t)-\frac{a_{1}}{2b_{1}}]^{2}+\frac {a_{1}^{2}}{4b_{1}}\leq V((n+l)\tau)+\zeta\), where \(\zeta=\frac{a_{1}^{2}}{4b_{1}}\). When \(t=(n+1)\tau\), \(V((n+1)\tau^{+})= (1-\mu)x((n+1)\tau)+(1-\mu_{1})y((n+1)\tau) \leq V((n+1)\tau)\). By the lemma of [24], for \(t\in(n\tau,(n+l)\tau]\) and \(t\in( (n+l)\tau,(n+1)\tau]\), we have
So \(V(t)\) is uniformly ultimately bounded. Hence, by the definition of \(V(t)\), we have that there exists a constant \(M>0\) such that \(x(t)\leq M\), \(y(t)\leq M\) for t large enough. The proof is complete. □
If \(x(t)=0\), we can easily have the subsystem of system (2.1) as follows:
We can easily obtain the analytic solution of system (3.1) between pulses, i.e.,
Considering the second and fourth equations of system (3.1), we have the stroboscopic map of system (3.1)
Two fixed points of (3.3) are obtained as \(P_{1}(0)\) and \(P_{2}(y^{\ast})\), where
Lemma 3.2
[25]
Consider the following difference equation:
\(z^{\ast}\) satisfies
then \(z^{\ast}\) is called equilibrium of (3.5), and if
then the unique equilibrium \(z^{\ast}\) of (3.5) is globally asymptotically stable. Otherwise, it is not stable.
Lemma 3.3
-
(i)
If \((1-\mu_{1})(a_{1}+1)e^{-(d_{1}l\tau+d_{3}(1-l)\tau)}<1\), the fixed point \(P_{1}(0)\) of (3.3) is globally asymptotically stable.
-
(ii)
If \((1-\mu_{1})(a_{1}+1)e^{-(d_{1}l\tau+d_{3}(1-l)\tau)}>1\), the fixed point \(P_{2}(y^{\ast})\) of (3.3) is globally asymptotically stable.
Proof
Making notation as
then
and
From Lemma 3.2, we obtain that the fixed points \(P(0)\) and \(P(y^{\ast })\) of (3.3) are stable, and then they are globally asymptotically stable. □
It is well known that the following lemma can easily be proved.
Lemma 3.4
-
(i)
If \((1-\mu_{1})(a_{1}+1)e^{-[d_{1}l+d_{3}(1-l)]\tau}<1\), the triviality periodic solution of system (3.1) is globally asymptotically stable.
-
(ii)
If \((1-\mu_{1})(a_{1}+1)e^{-[d_{1}l+d_{3}(1-l)]\tau}>1\), the periodic solution \(\widetilde{y(t)}\) of system (3.1) is globally asymptotically stable, where \(\widetilde{y(t)}\) is defined as
$$ \widetilde{y(t)}= \left \{ \textstyle\begin{array}{l@{\quad}l} y^{\ast}e^{-d_{1}(t-n\tau)},& t\in(n\tau,(n+l)\tau], \\ (e^{-d_{1}l\tau}y^{\ast})e^{-d_{3}(t-(n+l)\tau)},&t\in((n+l)\tau ,(n+1)\tau], \end{array}\displaystyle \right . $$(3.11)and \(y^{\ast}\) is defined as (3.4).
4 Dynamics for system (2.1)
Theorem 4.1
Let \((x(t),y(t))\) be any solution of system (2.1). If
and
hold, then the prey-extinction boundary periodic solution \((0,\widetilde {y(t)})\) of (2.1) is globally asymptotically stable, where \(y^{\ast}\) is defined as (3.4).
Proof
First, we prove the local stability. Defining \(x_{1}(t)=x(t)\), \(y_{1}(t)=y(t)-\widetilde{y(t)}\), then we have the following linearly similar system for system (2.1) which concerns one periodic solution \((0, \widetilde{y(t)})\) to
and
It is easy to obtain the fundamental solution matrix
There is no need to calculate the exact form of ∗, as it is not required in the analysis that follows, and
There is no need to calculate the exact form of ⋆, as it is not required in the analysis that follows.
The linearization of the third and fourth equations of (2.1) is
and the linearization of the seventh and eighth equations of (2.1) is
The stability of the periodic solution \((0,\widetilde{y(t)})\) is determined by the eigenvalues of
where
and
According to the Floquet theory [24], if \(| \lambda_{1}|<1\) and \(| \lambda_{2}|<1\), i.e.,
and
hold, then \((0,\widetilde{y(t)})\) is locally stable, where \(y^{\ast}\) is defined as (3.4).
In the following, we will prove the global attraction. Choose \(\varepsilon>0\) such that
From the first and fifth equations of (2.1), we notice that
and
so we consider the following impulsive differential equation:
From Lemma 3.4 and the comparison theorem of impulsive equation [24], we have \(y(t)\leq y_{2}(t)\) and \(y_{2}(t)\rightarrow\widetilde{y(t)}\) as \(t\rightarrow\infty\). Then
for all t large enough, for convenience we may assume that (4.2) holds for all \(t\geq0\). From (2.1) and (4.2), we get
So
Hence \(x(n\tau)\leq x(0^{+})\rho^{n}\) and \(x(n\tau)\rightarrow0\) as \(n\rightarrow\infty\), therefore \(x(t)\rightarrow0\) as \(t\rightarrow\infty\).
Next we prove that \(y(t)\rightarrow\widetilde{y(t)}\) as \(t\rightarrow\infty\). For \(\varepsilon<\min\{\frac{d_{1}}{k_{1}\beta _{1}},\frac{d_{3}}{k_{2}\beta_{2}}\}\), there must exist \(t_{0}>0\) such that \(0< x(t)<\varepsilon\) for all \(t\geq t_{0}\). Without loss of generality, we may assume that \(0< x(t)<\varepsilon\) for all \(t\geq{0}\), then for system (2.1) we have
and
Then we have \(z_{2}(t)\leq y(t)\leq z_{1}(t)\) and \(z_{1}(t)\rightarrow\widetilde{y(t)}\), \(z_{2}(t)\rightarrow \widetilde{z_{2}(t)}\) as \(t\rightarrow\infty\). While \(z_{1}(t)\) and \(z_{2}(t)\) are the solutions of
and
respectively,
where \(z_{2}^{\ast}\) is defined as
Therefore, for any \(\varepsilon_{1}>0\), there exists \(t_{1}\), \(t>t_{1}\), such that
Let \(\varepsilon\rightarrow0\), so we have
for t large enough, which implies \(y(t)\rightarrow \widetilde{y(t)}\) as \(t\rightarrow\infty\). This completes the proof. □
The next work is the investigation of permanence of system (2.1). Before starting our theorem, we give the following definition.
Definition 4.2
System (2.1) is said to be permanent if there are constants \(m,M >0 \) (independent of initial value) and a finite time \(T_{0}\) such that for all solutions \((x(t), y(t))\) with all initial values \(x(0^{+})>0\), \(y(0^{+})>0\), \(m\leq x(t)\leq M\), \(m\leq y(t)\leq M\) hold for all \(t\geq T_{0}\). Here \(T_{0}\) may depend on the initial values \((x(0^{+}), y(0^{+}))\).
Theorem 4.3
Let \((x(t),y(t))\) be any solution of system (2.1). If
and
hold, then system (2.1) is permanent.
Proof
Suppose \((x(t), y(t))\) is a solution of (2.1) with \(x(0)>0\), \(y(0)>0\). By Lemma 3.1, we have proved that there exists a constant \(M >0\) such that \(x(t)\leq M\), \(y(t)\leq M\) for t large enough, we may assume \(x(t)\leq M\), \(y(t)\leq M\) for \(t\geq0\). From Theorem 4.1, we know \(y(t)>\widetilde{y(t)}-\varepsilon_{2}\) for all t large enough and \(\varepsilon_{2}> 0\), so \(y(t)\geq e^{-d_{1}l\tau}y^{\ast }(1+e^{-d_{3}(1-l)\tau})-\varepsilon_{2}=m_{2}\) for t large enough. Thus, we only need to find \(m_{1}>0\) such that \(x(t)\geq m_{1}\) for t large enough, we will do it in what follows.
By the conditions of this theorem, we can select \(m_{3}>0\), \(\varepsilon_{1}> 0\) small enough such that \(m_{3}<\min \{\frac{d_{1}}{k_{1}\beta_{1}},\frac{d_{3}}{k_{2}\beta_{2}}\}\), \(\sigma=al\tau-\beta_{1}\varepsilon-\beta_{2}\varepsilon -d_{2}(1-l)\tau-\frac{\beta_{1}z^{\ast}}{d_{1}-k_{1}\beta _{1}m_{3}}\times(1-e^{-(d_{1}-k_{1}\beta_{1}m_{3})l\tau}) -\frac{\beta_{2}e^{-(d_{1}-k_{1}\beta_{1}m_{3})l\tau}z^{\ast }}{d_{1}-k_{1}\beta_{1}m_{3}} (1-e^{-(d_{3}-k_{2}\beta _{2}m_{3})(1-l)\tau})> 0\) and
\((1-\mu_{1})(a_{1}+1)e^{-[(d_{1}-k_{1}\beta_{1}m_{3})l\tau +(d_{3}-k_{2}\beta_{2}m_{3})(1-l)\tau]}>1\). We will prove that \(x(t)< m_{3}\) cannot hold for \(t\geq 0\). Otherwise,
By Lemma 3.4, we have \(y(t)\geq z(t)\) and \(z(t)\rightarrow\overline{z(t)}\), \(t\rightarrow\infty\), where \(z(t)\) is the solution of
and
where \(z^{\ast}\) is defined as
Therefore, there exists \(T_{1}>0\) such that
and
For \(t\geq T_{1}\), let \(N_{1}\in N\) and \(N_{1}\tau> T_{1}\). Integrating (4.14) on \((n\tau,(n+1)\tau)\), \(n\geq N_{1}\), we have
then \(x((N_{1}+k)\tau)\geq (1-\mu_{1})^{k}x(N_{1}\tau^{+})e^{k\sigma}\rightarrow\infty\), as \(k\rightarrow\infty\), which is a contradiction to the boundedness of \(x(t)\). Hence there exists \(t_{1}>0\) such that \(x(t)\geq m_{1}\). The proof is complete. □
5 Discussion
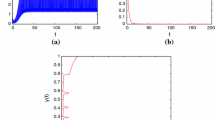
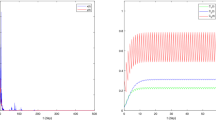
In this paper, according to the fact, a periodic impulsive switched predator-prey system with hibernation and birth pulse is proposed and investigated, we analyzed global asymptotic stability of the prey-extinction periodic solution of system (2.1) and obtained the conditions for the permanence of system (2.1). If it is assumed that \(x(0)=0.1\), \(y(0)=0.1\), \(a=2\), \(b=0.8\), \(d_{1}=0.5\), \(d_{2}=0.5\), \(d_{3}=0.4\), \(\beta_{1}=0.6\), \(k_{1} =0.3\), \(\beta_{2}=0.3\), \(k_{2}=0.4\), \(\mu =0.7\), \(\mu_{1}=0\), \(l=0.8\), \(\tau=1\), then the prey-extinction periodic solution \((0,\widetilde{y(t)})\) of system (2.1) is globally asymptotically stable (see Figure 1). If we assume that \(x(0)=0.1\), \(y(0)=0.1\), \(a=2\), \(b=0.8\), \(d_{1}=0.5\), \(d_{2}=0.5\), \(d_{3}=0.4\), \(\beta_{1}=0.6\), \(k_{1} =0.3\), \(\beta_{2}=0.3\), \(k_{2}=0.4\), \(\mu =0.5\), \(\mu_{1}=0\), \(l=0.8\), \(\tau=1\), then system (2.1) is permanent (see Figure 2).

Globally asymptotically stable prey-extinction periodic solution of system ( 2.1 ) with \(\pmb{x(0)=0.1}\) , \(\pmb{y(0)=0.1}\) , \(\pmb{a=2}\) , \(\pmb{b=0.8}\) , \(\pmb{d_{1}=0.5}\) , \(\pmb{d_{2}=0.5}\) , \(\pmb{d_{3}=0.4}\) , \(\pmb{\beta_{1}=0.6}\) , \(\pmb{k_{1} =0.3}\) , \(\pmb{\beta _{2}=0.3}\) , \(\pmb{k_{2}=0.4}\) , \(\pmb{\mu=0.7}\) , \(\pmb{l=0.8}\) , \(\pmb{\tau=1}\) . (a) Time-series of \(x(t)\); (b) time-series of \(y(t)\); (c) the phase portrait of the globally asymptotically stable prey-extinction periodic solution of system (2.1).

The permanence of system ( 2.1 ) with \(\pmb{x(0)=0.1}\) , \(\pmb{y(0)=0.1}\) , \(\pmb{a=2}\) , \(\pmb{b=0.8}\) , \(\pmb{d_{1}=0.5}\) , \(\pmb{d_{2}=0.5}\) , \(\pmb{d_{3}=0.4}\) , \(\pmb{\beta_{1}=0.6}\) , \(\pmb{k_{1} =0.3}\) , \(\pmb{\beta _{2}=0.3}\) , \(\pmb{k_{2}=0.4}\) , \(\pmb{\mu=0.5}\) , \(\pmb{l=0.8}\) , \(\pmb{\tau=1}\) . (a) Time-series of \(x(t)\); (b) time-series of \(y(t)\); (c) the phase portrait of the permanence of system (2.1).
From the simulation experiment of Figures 1 and 2, the parameter μ affects the dynamical behaviors of system (2.1). If all parameters of system (2.1) are fixed, when \(\mu=0.7\), the prey population of system (2.1) goes extinct; when \(\mu=0.5\), system (2.1) is permanent. From Theorem 4.1 and Theorem 4.3, we can easily deduce that there must exist a threshold \(\mu^{\ast}\). If \(\mu >\mu^{\ast}\), the prey-extinction periodic solution \((0,\widetilde {y(t)})\) of system (2.1) is globally asymptotically stable. If \(\mu<\mu^{\ast}\), system (2.1) is permanent. That is to say, impulsive harvesting rate of the prey population plays an important role in system (2.1). The impulsive harvesting rate of the prey population will also reduce the predator population. It tells us that destroying or excessive exploiting of the prey population will cause extinction of the predator population. Our results also provide reliable tactic basis for the practical biological economics management and the protection of biodiversity.
References
Wang, LCH, Lee, TF: Torpor and hibernation in mammals: metabolic, physiological, and biochemical adaptations. In: Fregley, MJ, Blatteis, CM (eds.) Handbook of Physiology: Environmental Physiology, pp. 507-532. Oxford University Press, New York (1996)
Staples, JF, Brown, JCL: Mitochondrial metabolism in hibernation and daily torpor: a review. J. Comp. Physiol., B 178, 811-827 (2008)
Clark, CW: Mathematical Bioeconomics. Wiley, New York (1990)
Liu, Z, Tan, R: Impulsive harvesting and stocking in a Monod-Haldane functional response predator-prey system. Chaos Solitons Fractals 34(2), 454-464 (2007)
Dong, L, Chen, L, Sun, L: Extinction and permanence of the predator-prey system with stocking of prey and harvesting of predator impulsively. Math. Methods Appl. Sci. 29, 415-425 (2006)
Liu, M, Bai, C: Optimal harvesting policy of a stochastic food chain population model. Appl. Math. Comput. 245, 265-270 (2014)
Leard, B, Rebaza, J: Analysis of predator-prey models with continuous threshold harvesting. Appl. Math. Comput. 217, 5265-5278 (2011)
Zhao, T, Tang, S: Impulsive harvesting and by-catch mortality for the theta logistic model. Appl. Math. Comput. 217, 9412-9423 (2011)
Gakkhar, S, Singh, B: The dynamics of a food web consisting of two preys and a harvesting predator. Chaos Solitons Fractals 34(4), 1346-1356 (2007)
Song, X, Chen, LS: Optimal harvesting and stability for a predator-prey system with stage structure. Acta Math. Appl. Sinica (Engl. Ser.) 18(3), 423-430 (2002)
Jiao, J, Meng, X, Chen, L: Harvesting policy for a delayed stage-structured Holling II predator-prey model with impulsive stocking prey. Chaos Solitons Fractals 41, 103-112 (2009)
Sangoh, B: Management and Analysis of Biological Populations. Elsevier, Amsterdam (1980)
Jiao, J, Chen, L, Cai, S: Dynamical analysis of a biological resource management model with impulsive releasing and harvesting. Adv. Differ. Equ. 2012, 9 (2012)
Li, L, Wang, W: Dynamics of a Ivlev-type predator-prey system with constant rate harvesting. Chaos Solitons Fractals 41(4), 2139-2153 (2009)
Jiao, J, Chen, L: An appropriate pest management SI model with biological and chemical control concern. Appl. Math. Comput. 196, 285-293 (2008)
Liu, XN, Chen, LS: Complex dynamics of Holling II Lotka-Volterra predator-prey system with impulsive perturbations on the predator. Chaos Solitons Fractals 16, 311-320 (2003)
Chen, LS, Chen, J: Nonlinear Biological Dynamic Systems. Science Press, Beijing (1993) (in Chinese)
Jiao, J, Chen, L: Nonlinear incidence rate of a pest management SI model with biological and chemical control concern. Appl. Math. Mech. 28(4), 541-551 (2007)
Song, X, Li, Y: Dynamic complexities of a Holling II two-prey one-predator system with impulsive effect. Chaos Solitons Fractals 33(2), 463-478 (2007)
Jiao, J, Cai, S, Chen, L: Dynamical analysis of a three-dimensional predator-prey model with impulsive harvesting and diffusion. Int. J. Bifurc. Chaos 21(2), 453-465 (2011)
Jiao, J, Chen, L: Dynamical analysis of a delayed predator-prey system with birth pulse and impulsive harvesting at different moments. Adv. Differ. Equ. 2010, Article ID 954684 (2010)
Jiao, J, Meng, X, Chen, L: Harvesting policy for a delayed stage-structured Holling II predator-prey model with impulsive stocking prey. Chaos Solitons Fractals 41(1), 103-112 (2009)
Lakshmikantham, V, Bainov, DD, Simeonov, P: Theory of Impulsive Differential Equations. World Scientific, Singapore (1989)
Bainov, D, Simeonov, P: Impulsive Differential Equations: Periodic Solutions and Applications. Pitman Monographs and Surveys in Pure and Applied Mathematics, vol. 66 (1993)
Chen, L, Chen, J: Nonlinear Biology Dynamical System. Scientific Press, Beijing (1993)
Acknowledgements
This work was supported by National Natural Science Foundation of China (11361014, 10961008).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
JJ carried out the main part of this article, LC corrected the manuscript, SC and LL brought forward some suggestions on this article. All authors have read and approved the final manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Jiao, J., Cai, S., Li, L. et al. Dynamics of a periodic impulsive switched predator-prey system with hibernation and birth pulse. Adv Differ Equ 2015, 174 (2015). https://doi.org/10.1186/s13662-015-0460-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-015-0460-4