
Abstract
A Lotka–Volterra commensal symbiosis model with density dependent birth rate that takes the form
where \(b_{ij}\), \(i=1, 2\), \(j=1, 2, 3, 4\), \(a_{11}\), \(a_{12} \), and \(a_{22}\) are all positive constants, is proposed and studied in this paper. The system may admit four nonnegative equilibria. By constructing some suitable Lyapunov functions, we show that under some suitable assumptions, all of the four equilibria may be globally asymptotically stable, such a property is quite different to the traditional Lotka–Volterra commensalism model. With introduction of the density dependent birth rate, the dynamic behaviors of the commensalism model become complicated.
Similar content being viewed by others

1 Introduction
The aim of this paper is to investigate the dynamic behaviors of the following commensalism model with density dependent birth rate:
where \(b_{ij}, i=1, 2, j=1, 2,3, 4\), \(a_{11}\), \(a_{12} \), and \(a_{22}\) are all positive constants. \(x(t)\), \(y(t)\) are the densities of the first and second species at time t, respectively. Here we make the following assumptions:
-
(a)
\(\frac{b_{11}}{b_{12}+b_{13}x}\) is the birth rate of the first species which is density dependent, the birth rate of the species is declining as the density of the species is increasing;
-
(b)
\(b_{14}\) is the death rate of the first species, \(a_{11}\) is the density dependent coefficient of the first species;
-
(c)
\(\frac{b_{21}}{b_{22}+b_{23}y}\) is the birth rate of the second species, it is declining as the density of the species is increasing;
-
(d)
\(b_{24}\) is the death rate of the second species, \(a_{22}\) is the density dependent coefficient of the second species;
-
(e)
The relationship between the two species is commensalism, i.e., the second species has positive effect on the first species, while the first species has no influence on the second species, we describe such of relationship by using the bilinear function \(a_{12}xy\).
During the last decades, many scholars investigated the dynamic behaviors of the mutualism model or commensalism model [1, 2]. Some essential progress has been made in this direction. Such topics as the stability of the positive equilibrium [1, 3–20], the persistence of the system [21–27], the existence of the positive periodic solution [17, 28–30], the extinction of the species [2, 21, 31], the influence of harvesting [3, 9, 11, 12, 19], the influence of feedback control variables [1, 8, 18, 21, 22, 25, 26], the influence of stage structure [5, 7], etc. have been extensively investigated.
Sun and Wei [4] for the first time proposed and studied the following two species commensalism symbiosis model:
where x and y are the densities of the first and second species at time t, respectively. System (1.2) admits four equilibria \(E_{1}(0, 0), E_{2}(k_{1},0), E_{3}(0, k_{2})\), and \(E_{4}(k_{1}+\alpha k_{2} k_{2})\).
Concerned with the stability property of the above equilibria, Sun and Wei [4] obtained the following result:
Theorem A
\(E_{1}(0, 0), E_{2}(k_{1},0), E_{3}(0, k_{2})\) are all unstable, \(E_{4}(k_{1}+\alpha k_{2}, k_{2})\) is locally stable.
Noting that the authors of [4] did not give any global stability property of the equilibrium, with the aim of putting forward the study in this direction, Han and Chen [18] proposed the following commensalism model:
System (1.3) admits a positive equilibrium \(P_{0}(x_{0},y_{0})\), where
Concerned with the stability property of this equilibrium, by constructing some suitable Lyapunov function, the authors obtained the following result:
Theorem B
The positive equilibrium \(P_{0}(x_{0},y_{0})\) of system (1.3) is globally stable.
It came to our attention that in system (1.3), if we did not consider the relationship of the two species, then the equations for both species reduce to the traditional logistic equation. For example, the first species takes the form
where \(b_{1}\) is the intrinsic growth rate and \(a_{11}\) is the density dependent coefficient. System (1.4) could be revised as
where \(b_{11}\) is the birth rate of the species and \(b_{14}\) is the death rate of the species. Already, Brauer and Castillo-Chavez [32], Tang and Chen [33], Berezansky et al. [34] have showed that in some cases the density dependent birth rate of the species is more suitable. If we take the famous Beverton–Holt function [34] as the birth rate, then system (1.5) should be revised to
Similarly, the second species could be expressed as follows:
(1.6), (1.7) together with the cooperation relationship between the species will lead to system (1.1).
As far as system (1.1) is concerned, one interesting issue is to find out the influence of the nonlinear density birth rate. Is it possible for system (1.1) to admit some similar dynamic behaviors as those of systems (1.2) and (1.3), or does system (1.1) admit some new characteristic property?
The aim of this paper is to find out the answers to the issues above. The rest of the paper is arranged as follows. In Sect. 2, we investigate the stability property of the equilibria of system (1.1); Sect. 3 presents some numerical simulations to show the feasibility of the main results. We end this paper with a brief discussion.
2 Global asymptotic stability
We first establish a lemma, which is useful for proving the main result.
Lemma 2.1
Consider the following equation:
Assume that \(a>bd\), then the unique positive equilibrium \(y^{*} \) of system (2.1) is globally asymptotically stable.
Proof
Set
Since \(a>bd\), it follows that \(F(0)= \frac{a}{b}-d>0\). Also,
hence \(F(y)\) is a strictly decreasing function. One could easily see that \(F(+\infty )=-\infty \), thus there exists a unique positive solution \(y^{*}\) such that \(F(y^{*})=0\). Indeed,
The above analysis shows that
-
(1)
There is \(y^{*}\), as expressed by (2.2), such that \(F(y^{*}) = 0\);
-
(2)
For all \(y^{*} > y > 0, F(y)= \frac{a}{b+cy}-d-ey > 0\);
-
(3)
For all \(y > y^{*} > 0, F(y)= \frac{a}{b+cy}-d-ey < 0\).
Now let us consider the Lyapunov function
Direct calculation shows that
Thus, \(y^{*}\) is globally asymptotically stable. This ends the proof of Lemma 2.1. □
Now we are in a position to consider the existence and stability property of the equilibria of system (1.1). The equilibrium of system (1.1) is determined by the equation
System (1.1) always admits a boundary equilibrium \(A_{1}(0,0)\). Assume that
holds, then
admits a unique positive solution \(x^{*}\), where
Assume that (2.4) and
hold, then system (1.1) admits the nonnegative boundary equilibrium \(A_{2}(x^{*},0)\). Assume that
holds, then
admits a unique positive solution \(y^{*}\), where
Assume that (2.7) and
hold, then system (1.1) admits the nonnegative boundary equilibrium \(A_{3}(0,y^{*})\). Assume that (2.7) and
hold, then system (1.1) admits the unique positive equilibrium \(A_{4}(x_{1},y_{1})\), where
and \(x_{1}\) is the unique positive solution of the equation
Remark 2.1
From the above discussion, one could easily see that the inequalities
and
is enough to ensure the existence of the unique positive equilibrium of system (1.1).
Concerned with the stability property of the above four nonnegative equilibria, we have the following result.
Theorem 2.1
-
(1)
Assume that (2.7) and (2.10) hold, then system (1.1) admits a unique positive equilibrium \(A_{4}(x_{1},y_{1})\), which is globally asymptotically stable;
-
(2)
Assume that (2.7) and (2.9) hold, then the boundary equilibrium \(A_{3}(0,y^{*})\) is globally asymptotically stable;
-
(3)
Assume that (2.4) and (2.6) hold, then the boundary equilibrium \(A_{2}(x^{*},0)\) is globally asymptotically stable;
-
(4)
Assume that
$$ b_{11}< b_{12}b_{14} $$(2.15)and (2.6) hold, then the boundary equilibrium \(A_{1}(0,0)\) is globally asymptotically stable.
Remark 2.2
Conditions (2.7) and (2.10) are necessary to ensure that system (1.1) admits a positive equilibrium. Hence, it follows from Theorem 2.1(1) that if system (1.1) admits a positive equilibrium, it is globally asymptotically stable.
Remark 2.3
From Remark 2.2 it immediately follows that under the assumption that (2.13) and (2.14) hold, system (1.1) admits a unique globally asymptotically stable positive equilibrium.
Proof of Theorem 2.1
(1) Obviously, \(x_{1}\), \(y_{1}\) satisfy the equations
Now let us consider the Lyapunov function
One could easily see that the function \(V_{1}\) is zero at the positive equilibrium \(A_{4}(x_{1}, y_{1})\) and is positive for all other positive values of x, y. By applying (2.16), the time derivative of \(V_{1}\) along the trajectories of (1.1) is
It then follows from (2.18) that \(D^{+}V_{1}(t)<0\) strictly for all \(x, y>0\) except the positive equilibrium \(A_{4}(x_{1}, y_{1})\), where \(D^{+}V_{1}(t)=0\). Thus, \(V_{1}(x,y)\) satisfies Lyapunov’s asymptotic stability theorem [35], and the positive equilibrium \(A_{4}(x_{1}, y_{1})\) of system (1.1) is globally asymptotically stable.
(2) Inequality (2.9) implies that for enough small positive constant ε, one has
holds. Obviously, \(y^{*}\) satisfies the equation
Also, it follows from Lemma 2.1 that the unique positive equilibrium \(y^{*}\) of system
is globally asymptotically stable. That is,
Hence, for ε satisfies (2.19), there exists enough large \(T_{1}\) such that
Now let us consider the Lyapunov function
One could easily see that the function \(V_{2}\) is zero at the boundary equilibrium \(A_{3}(0, y^{*})\) and is positive for all other positive values of x, y. By applying (2.20) and (2.23), for \(t>T_{1}\), the time derivative of \(V_{2}\) along the trajectories of (1.1) is
It then follows from (2.19) that \(D^{+}V_{2}(t)<0\) strictly for all \(x, y>0\) except the boundary equilibrium \(A_{3}(0, y^{*})\), where \(D^{+}V_{2}(t)=0\). Thus, \(V_{2}(x,y)\) satisfies Lyapunov’s asymptotic stability theorem [35], and the boundary equilibrium \(A_{3}(0, y^{*})\) of system (1.1) is globally asymptotically stable.
(3) Obviously, \(x^{*}\) satisfies the equation
Now let us consider the Lyapunov function
One could easily see that the function \(V_{3}\) is zero at the boundary equilibrium \(A_{2}(x^{*}, 0)\) and is positive for all other positive values of x, y. By applying (2.25), the time derivative of \(V_{3}\) along the trajectories of (1.1) is
It then follows from (2.6) that \(D^{+}V_{3}(t)<0\) strictly for all \(x, y>0\) except the boundary equilibrium \(A_{2}(x^{*}, 0)\), where \(D^{+}V_{3}(t)=0\). Thus, \(V_{3}(x,y)\) satisfies Lyapunov’s asymptotic stability theorem [35], and the boundary equilibrium \(A_{2}(x^{*}, 0)\) of system (1.1) is globally asymptotically stable.
(4) Now let us consider the Lyapunov function
One could easily see that the function \(V_{4}\) is zero at the boundary equilibrium \(A(0, 0)\) and is positive for all other positive values of x, y. The time derivative of \(V_{4}(x,y)\) along the trajectories of (1.1) is
It then follows from (2.6) and (2.15) that \(D^{+}V_{4}(t)<0\) strictly for all \(x, y>0\) except the boundary equilibrium \(A_{1}(0, 0)\), where \(D^{+}V_{4}(t)=0\). Thus, \(V_{4}(x,y)\) satisfies Lyapunov’s asymptotic stability theorem [35], and the boundary equilibrium \(A_{1}(0, 0)\) of system (1.1) is globally asymptotically stable.
This ends the proof of Theorem 2.1. □
3 Numeric simulations
Now let us consider the following four examples.
Example 3.1
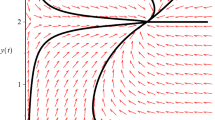
In this system, corresponding to system (1.1), we take \(b_{11}=b_{13}=b_{14}=a_{11}=a_{12}=b_{21}=b_{23}=b_{24}=a_{22}=1, b_{12}=b_{22}=2\). Since \(b_{11}< b_{12}b_{14}, b_{21}< b_{22}b_{24}\), it follows from Theorem 2.1(4) that the boundary equilibrium \(A_{1}(0,0)\) is globally asymptotically stable. Figure 1 supports this assertion.

Dynamic behaviors of system (3.1) with the initial condition \((x(0),y(0)) =(1,0.3),(0.4,2),(0.02,2),(1, 2)\), and \((0.1,2)\), respectively
Example 3.2
In this system, corresponding to system (1.1), we take \(b_{12}=b_{13}=b_{14}=a_{11}=a_{12}=b_{22}=b_{23}=b_{24}=a_{22}=1, b_{11}=b_{21}=2\). Since \(b_{11}>b_{12}b_{14}, b_{21}>b_{22}b_{24}\), it follows from Remark 2.3 that the unique positive equilibrium \(A_{4}(0.4142,0.6364)\) is globally asymptotically stable. Figure 2 supports this assertion.

Dynamic behaviors of system (3.2) with the initial condition \((x(0),y(0)) =(1,0.3),(0.4,2),(0.02,2),(1, 2)\), and \((0.1,2)\), respectively
Example 3.3
In this system, corresponding to system (1.1), we take \(b_{12}=b_{13}=b_{14}=a_{11}=a_{12}=b_{21}=b_{23}=b_{24}=a_{22}=1, b_{11}=b_{22}=2\). Since \(b_{11}>b_{12}b_{14}, b_{21}< b_{22}b_{24}\), that is, inequalities (2.4) and (2.6) hold, it follows from Theorem 2.1(3) that the boundary equilibrium \(A_{2}(0.4142,0)\) is globally asymptotically stable. Figure 3 supports this assertion.

Dynamic behaviors of system (3.3) with the initial condition \((x(0),y(0)) =(1,0.3),(0.4,2),(0.02, 2),(1, 2)\), and \((0.1,2)\), respectively
Example 3.4
In this system, corresponding to system (1.1), we take \(b_{11}=b_{13}=b_{14}=a_{11}=a_{12}=b_{22}=b_{23}=b_{24}=a_{22}=1, b_{12}=b_{21}=2\). Since \(b_{11}< b_{12}b_{14}, b_{21}>b_{22}b_{24}\), that is, inequalities (2.7) and (2.9) hold, it follows from Theorem 2.1(2) that the boundary equilibrium \(A_{2}(0, 0.4142)\) is globally asymptotically stable. Figure 4 supports this assertion.

Dynamic behaviors of system (3.4) with the initial condition \((x(0),y(0)) =(0.4,1),(1,0.3),(0.02,1),(1, 1)\), and \((1,0.1)\), respectively
4 Discussion
Recently, many scholars have studied the dynamic behaviors of the commensal symbiosis model [13–31]. All of the works of [13–31] are based on the traditional logistic model, as was showed in the introduction section. Especially, Han and Chen [18] showed that the unique positive equilibrium \(P_{0}(x_{0},y_{0})\) of system (1.3) is globally asymptotically stable (see Theorems A and B in the Introduction section for more details), this means that all other equilibria of system (1.3) are unstable.
In this paper, we argued that the birth rate of the species may be density dependent; indeed, this is one of the phenomena that could be observed in the nature and society. We propose system (1.1). Theorem 2.1 shows that under some suitable assumptions, all of the four equilibria may be globally asymptotically stable. That is, with introduction of the density dependent birth rate, the dynamic behaviors of the system become complicated. Such kind of phenomenon is not observed in [13–31].
Our study shows that the birth rate is one of the essential factors in determining the dynamic behaviors of the species. To control the number of the species, maybe one of the useful methods is to control the birth rate of the species.
References
Yang, K., Miao, Z.S., Chen, F.D., Xie, X.D.: Influence of single feedback control variable on an autonomous Holling-II type cooperative system. J. Math. Anal. Appl. 435(1), 874–888 (2016)
Chen, F.D., Xie, X.D., Miao, Z.S.: Extinction in two species nonautonomous nonlinear competitive system. Appl. Math. Comput. 274, 119–124 (2016)
Deng, H., Huang, X.Y.: The influence of partial closure for the populations to a harvesting Lotka–Volterra commensalism model. Commun. Math. Biol. Neurosci. 2018, Article ID 10 (2018)
Sun, G.C., Wei, W.L.: Qualitative analysis of commensal symbiosis model of two population. Math. Theory Appl. 23(3), 65–68 (2003)
Chen, F.D., Xie, X.D., Chen, X.F.: Dynamic behaviors of a stage-structured cooperation model. Commun. Math. Biol. Neurosci. 2015, Article ID 4 (2015)
Yang, K., Xie, X.D., Chen, F.D.: Global stability of a discrete mutualism model. Abstr. Appl. Anal. 2014, 709124 (2014)
Li, T.T., Chen, F.D., Chen, J.H., Lin, Q.X.: Stability of a stage-structured plant-pollinator mutualism model with the Beddington–DeAngelis functional response. J. Nonlinear Funct. Anal. 2017, Article ID 50 (2017)
Han, R.Y., Chen, F.D., Xie, X.D., Miao, Z.S.: Global stability of May cooperative system with feedback controls. Adv. Differ. Equ. 2015, Article ID 360 (2015)
Xie, X.D., Chen, F.D., Xue, Y.L.: Note on the stability property of a cooperative system incorporating harvesting. Discrete Dyn. Nat. Soc. 2014, 327823 (2014)
Xie, X.D., Yang, K., Chen, F.D., Xue, Y.L.: Global attractivity of an integrodifferential model of mutualism. Abstr. Appl. Anal. 2014, 928726 (2014)
Lei, C.Q.: Dynamic behaviors of a non-selective harvesting may cooperative system incorporating partial closure for the populations. Commun. Math. Biol. Neurosci. 2018, Article ID 12 (2018)
Chen, F.D., Wu, H.L., Xie, X.D.: Global attractivity of a discrete cooperative system incorporating harvesting. Adv. Differ. Equ. 2016, Article ID 268 (2016)
Wu, R.X., Li, L., Zhou, X.Y.: A commensal symbiosis model with Holling type functional response. J. Math. Comput. Sci. 2016(2), 364–371 (2016)
Chen, J.H., Wu, R.X.: A commensal symbiosis model with non-monotonic functional response. Commun. Math. Biol. Neurosci. 2017, Article ID 5 (2017)
Wu, R.X., Li, L.: Dynamic behaviors of a commensal symbiosis model with ratio-dependent functional response and one party can not survive independently. J. Math. Comput. Sci. 16(3), 495–506 (2016)
Wu, R.X., Li, L., Lin, Q.F.: A Holling type commensal symbiosis model involving Allee effect. Commun. Math. Biol. Neurosci. 2018, Article ID 5 (2018)
Li, T.T., Lin, Q.X., Chen, J.H.: Stability analysis of a Lotka–Volterra type predator-prey system with Allee effect on the predator species. Commun. Math. Biol. Neurosci. 2016, Article ID 22 (2016)
Han, R.Y., Chen, F.D.: Global stability of a commensal symbiosis model with feedback controls. Commun. Math. Biol. Neurosci. 2015, Article ID 15 (2015)
Lin, Q.F.: Dynamic behaviors of a commensal symbiosis model with non-monotonic functional response and non-selective harvesting in a partial closure. Commun. Math. Biol. Neurosci. 2018, Article ID 4 (2018)
Lin, Q.F.: Allee effect increasing the final density of the species subject to the Allee effect in a Lotka–Volterra commensal symbiosis model. Adv. Differ. Equ. 2018, Article ID 196 (2018)
Han, R.Y., Xie, X.D., Chen, F.D.: Permanence and global attractivity of a discrete pollination mutualism in plant-pollinator system with feedback controls. Adv. Differ. Equ. 2016, Article ID 199 (2016)
Chen, J.H., Xie, X.D.: Feedback control variables have no influence on the permanence of a discrete N-species cooperation system. Discrete Dyn. Nat. Soc. 2009, Article ID 306425 (2009)
Zhao, L., Qin, B., Chen, F.: Permanence and global stability of a May cooperative system with strong and weak cooperative partners. Adv. Differ. Equ. 2018, Article ID 172 (2018)
Yang, L.Y., Xie, X.D., Chen, F.D.: Dynamic behaviors of a discrete periodic predator-prey-mutualist system. Discrete Dyn. Nat. Soc. 2015, 247269 (2015)
Chen, F.D., Yang, J.H., Chen, L.J., Xie, X.D.: On a mutualism model with feedback controls. Appl. Math. Comput. 214(2), 581–587 (2009). https://doi.org/10.1016/j.amc.2009.04.019
Chen, L.J., Chen, L.J., Li, Z.: Permanence of a delayed discrete mutualism model with feedback controls. Math. Comput. Model. 50(1), 1083–1089 (2009)
Yang, W.S., Li, X.P.: Permanence of a discrete nonlinear N-species cooperation system with time delays and feedback controls. Appl. Math. Comput. 218(7), 3581–3586 (2011)
Wang, D.H.: Dynamic behaviors of an obligate Gilpin–Ayala system. Adv. Differ. Equ. 2016, 270 (2016)
Xie, X.D., Miao, Z.S., Xue, Y.L.: Positive periodic solution of a discrete Lotka–Volterra commensal symbiosis model. Commun. Math. Biol. Neurosci. 2015, Article ID 2 (2015)
Xue, Y.L., Xie, X.D., Chen, F.D., Han, R.Y.: Almost periodic solution of a discrete commensalism system. Discrete Dyn. Nat. Soc. 2015, Article ID 295483 (2015). https://doi.org/10.1155/2015/295483
Miao, Z.S., Xie, X.D., Pu, L.Q.: Dynamic behaviors of a periodic Lotka–Volterra commensal symbiosis model with impulsive. Commun. Math. Biol. Neurosci. 2015, Article ID 3 (2015)
Brauer, F., Castillo-Chavez, C.: Mathematical Models in Population Biology and Epidemiology. Texts in Applied Mathematics, vol. 40. Springer, New York (2001). https://doi.org/10.1007/978-1-4757-3516-1
Tang, S.Y., Chen, L.S.: Density-dependent birth rate, birth pulses and their population dynamic consequences. J. Math. Biol. 44(2), 185–199 (2002)
Berezansky, L., Braverman, E., Idels, L.: Nicholsons blowflies differential equations revisited: main results and open problems. Appl. Math. Model. 34(6), 1405–1417 (2010). https://doi.org/10.1016/j.apm.2009.08.027
Chen, L.S.: Mathematical Models and Methods in Ecology. Science Press, Beijing (1988) (in Chinese)
Acknowledgements
The authors would like to thank Dr. Xinyu Guan for useful discussion about the mathematical modeling.
Funding
The research was supported by the National Natural Science Foundation of China under Grant (11601085) and the Natural Science Foundation of Fujian Province (2017J01400).
Author information
Authors and Affiliations
Contributions
All authors contributed equally to the writing of this paper. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that there is no conflict of interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Chen, F., Xue, Y., Lin, Q. et al. Dynamic behaviors of a Lotka–Volterra commensal symbiosis model with density dependent birth rate. Adv Differ Equ 2018, 296 (2018). https://doi.org/10.1186/s13662-018-1758-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-018-1758-9